日本落叶松×长白落叶松杂种幼林生长节律1)
2014-08-02孙晓梅张守攻
赖 猛 孙晓梅 张守攻
(林木遗传育种国家重点实验室(中国林业科学研究院林业研究所),北京,100091)
日本落叶松×长白落叶松杂种幼林生长节律1)
赖 猛 孙晓梅 张守攻
(林木遗传育种国家重点实验室(中国林业科学研究院林业研究所),北京,100091)
对造林后2、3年生57个日本落叶松×长白落叶松杂种无性系的物候性状和生长节律进行定株观察。物候期变异分析结果表明,各物候因子在无性系水平上的分化较明显。利用方差分析方法研究不同时期无性系间树高的变异,利用秩次相关方法确定不同时期树高生长量之间的关系。结果表明:不同时期测定的树高在无性系间的差异均达极显著水平;2年生杂种无性系的遗传变异系数、遗传方差占总方差百分率和无性系重复力等遗传参数,随时间的变化趋势大致可分为3个阶段,3年生杂种无性系各遗传参数随时间的变化可分为2个阶段;不同时期树高间的相关性均达极显著水平,相关系数表现出随时间的增长而持续上升的趋势;在一个年生长周期中,日本落叶松×长白落叶松杂种有2个较明显的速生期,6月初至7月中旬为第1个速生期,第2个速生期为整个8月份。
日本落叶松;长白落叶松;杂种;遗传变异;生长节律
Larixkaempferi;Larixolgensis; Hybrid; Genetic variation; Growth rhythm
落叶松(Larixspp.)具有适应性强,早期速生、成林快、病虫害少的特点,是我国主要针叶纸浆用材和工业用材树种,我国已成为世界上落叶松人工林经营的第一大国[1]。日本落叶松(Larixkaempferi)和长白落叶松(L.olgensis)在辽宁东部地区被广泛种植。研究表明日本落叶松×长白落叶松(L.kaempferi×L.olgensis)集合了双亲的优良性状,在生长干形、材性和抗逆等方面表现出明显的杂种优势[2-4]。
研究树种的生长节律和物候性状变异,不仅能为林木引种提供适应性、抗逆性的依据,同时也可为树木遗传改良提供亲本和早期选择的参考[3]。本文以辽东地区2、3年生日本落叶松×长白落叶松杂种无性系人工幼林为研究对象,对其物候期和生长节律进行了系统观察,研究1 a内不同时期树高生长量的遗传变异模式及彼此之间的相关关系,旨在揭示其早期的生长特性,以期为日本落叶松×长白落叶松杂种人工幼林的抚育管理提供可靠的科学依据。
1 材料与方法
1.1 杂种无性系来源
2003年在辽宁省清源县大孤家林场对日本落叶松和长白落叶松进行杂交制种,具体的组合见表1。2003年杂交→2004年播种→2005年移植→2006年营造试验林。

表1 杂交组合设计
试验林位于辽宁省清源县大孤家林场西南沟二板石沟,长白山系千山山脉龙岗支脉北坡,属中温带季风气候区,年均气温6 ℃,最低温度-30 ℃,最高温度34 ℃,年均降水量650 mm。林地土壤为棕色森林土,pH值为6.2~6.8。试验林共包括日本落叶松×长白落叶松杂种共57个无性系,来源于辽宁大孤家地区。2006年春用2年生扦插苗营建无性系评比林,采用随机完全区组设计,5次重复,6株小区,株行距2 m×2 m。
1.2 调查方法
2007年(2年生)对试验林进行物候性状的定株观测,2007—2008年(2、3年生)对试验林进行生长节律的定株观测。在Ⅰ~Ⅲ区组,从每个无性系的6个分生株中选取2棵生长发育正常的树为标准株,观测的物候相及方法同文献[3]。生长节律的调查为5月15日起到9月30日止,每5 d调查1次,记录树高生长量。
1.3 统计分析方法
对调查数据进行方差分析及有关遗传参数的估算。树高生长量无性系间方差分析采用线性固定模型[5]:
yij=μ+αi+βj+αβi(j)+εij。
式中:yij代表jth区组内ith无性系的观测值;μ代表平均值;αi代表ith无性系的固定效应;βi代表jth区组的固定效应;αβi(j)代表jth区组ith无性系的固定效应;εij代表随机误差。
无性系重复力计算公式如下[6-7]:
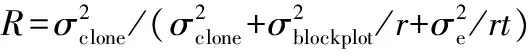
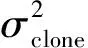
遗传变异系数公式如下[6-7]:

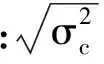
方差分析采用SAS/STAT9.0软件中PROCANOVA过程计算;方差分量用PROCVARCOMP过程中带限制的最大似然法计算[8]。
2 结果与分析
2.1 物候期变异
2007年(2年生)杂种无性系物候观测结果见表2。主、侧枝芽的物候期基本一致。4月上旬芽开始萌动,下旬时芽开始展开,5月上旬时芽已经完全展开。杂种无性系新梢开始生长发生在5月中旬;开始封顶的时间发生在8月下旬,完全封顶的时间大大致在9月中旬。
不同无性系间物候期差异较明显。春季各物候因子的早晚变异幅度较小,为6~10 d;封顶期和生长期物候因子的变异幅度较大,为31~37 d。从变异系数来看,侧枝芽膨胀的变异系数最大达18.28%,顶芽完全展叶的变异系数最小为4.11%。
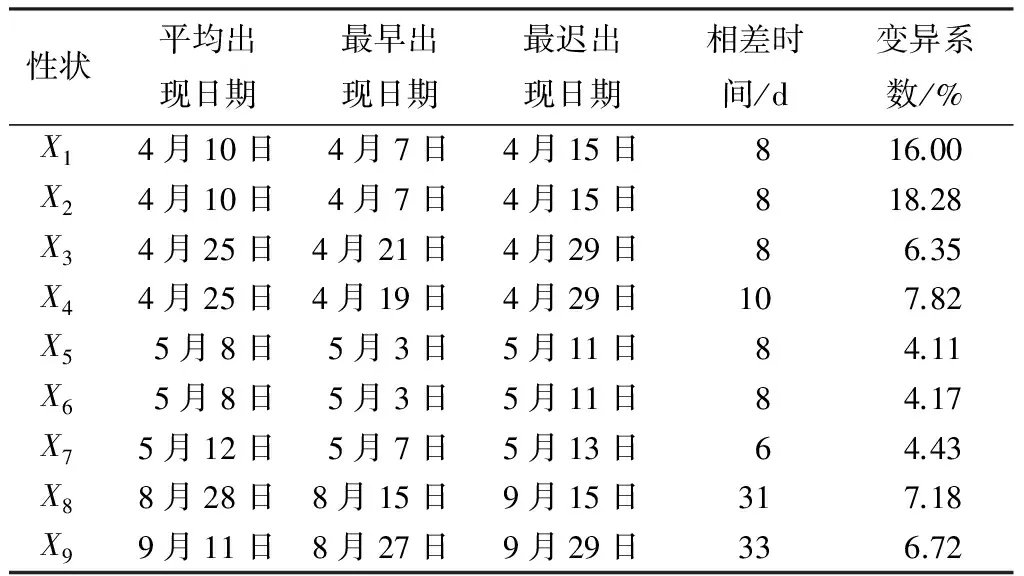
表2 杂种无性系各物候期的差异
注:x1为顶芽膨大;x2为侧枝芽膨大;x3为顶芽开始展叶;x4为侧枝芽开始展叶;x5为顶芽完全展叶;x6为侧枝芽完全展叶;x7为开始抽梢;x8为开始封顶;x9为完全封顶。
2.2 不同时期树高的遗传变异
对2、3年生日本落叶松×长白落叶松57个杂种无性系定株调查观测的28次树高进行遗传变异分析,结果见表3。在不同时期测定的树高在无性系间的差异均达极显著水平,说明树高性状在无性系水平上具有很大的遗传改良潜力;2年生无性系重复力的变化范围为0.55~0.82,3年生无性系为0.40~0.79,此结果说明,各无性系树高性状受中度或中度以上的遗传控制,具有较强的无性系选择潜力。
从表3中还可得知,2年生时,树高的遗传变异系数、遗传方差占总方差百分率和无性系重复力等遗传参数,随时间的变化趋势大致可分为3个阶段:5月15日至6月15日,树高的遗传变异系数、遗传方差占总方差百分率和无性系重复力均处于较高水平;6月15日后,其数值有一定程度的下降,并维持在较低水平直至7月底;进入8月份后,各遗传参数的数值相对第2阶段有一定程度的提高,并维持在中等偏上水平直至9月底生长结束。3年生时,各遗传参数随时间的变化大致可分为两个阶段:8月中旬以前,各遗传参数均处于较高水平;进入8月中旬以后,各遗传参数维持到中等水平直至生长结束。
2.3 不同时期树高间的秩次相关
大量研究表明,日本落叶松、长白落叶松生长性状的早晚相关紧密[9-11],那么一个生长周期内,不同时期的树高是否存在相关性呢?利用2、3年生57个杂种无性系不同时期28次测量的数据,计算年中各时期树高生长秩次相关系数(表4、表5)。不同时期的树高相关均达极显著水平,相关系数表现出随时间的增长而持续上升的趋势。自8月15日起,秩相关系数高且趋于稳定,2年生时为0.90~0.99,3年生时达0.96~0.99,此时起便可对当年生树高进行测量与研究。

表3 杂种无性系不同观测时期树高遗传变异
注:** 表示差异达极显著水平。

表4 2年生杂种无性系不同观测时期树高增长秩次相关
2.4 不同时期树高的生长节律
2.4.1 树高平均连续生长量
2、3年生日本落叶松×长白落叶松杂种无性系树高每5 d内平均连续生长量见图1。2、3年生的林木年生长进程均可划分为3个时期:生长前期、生长中期和生长后期,主要的生长趋势均为前期慢,中期快,后期慢,但生长量差别大。整体看,2、3年生无性系的生长前期为5月中旬至5月底,6—8月为生长中期,9月份开始进入生长后期。3年生无性系树高平均生长量为107.39 cm,是2年生无性系树高平均生长量56.60 cm的1.9倍。在6月中旬前,3年生无性系树高平均生长量的生长优势不明显,从6月下旬开始,两者的差距明显加大。
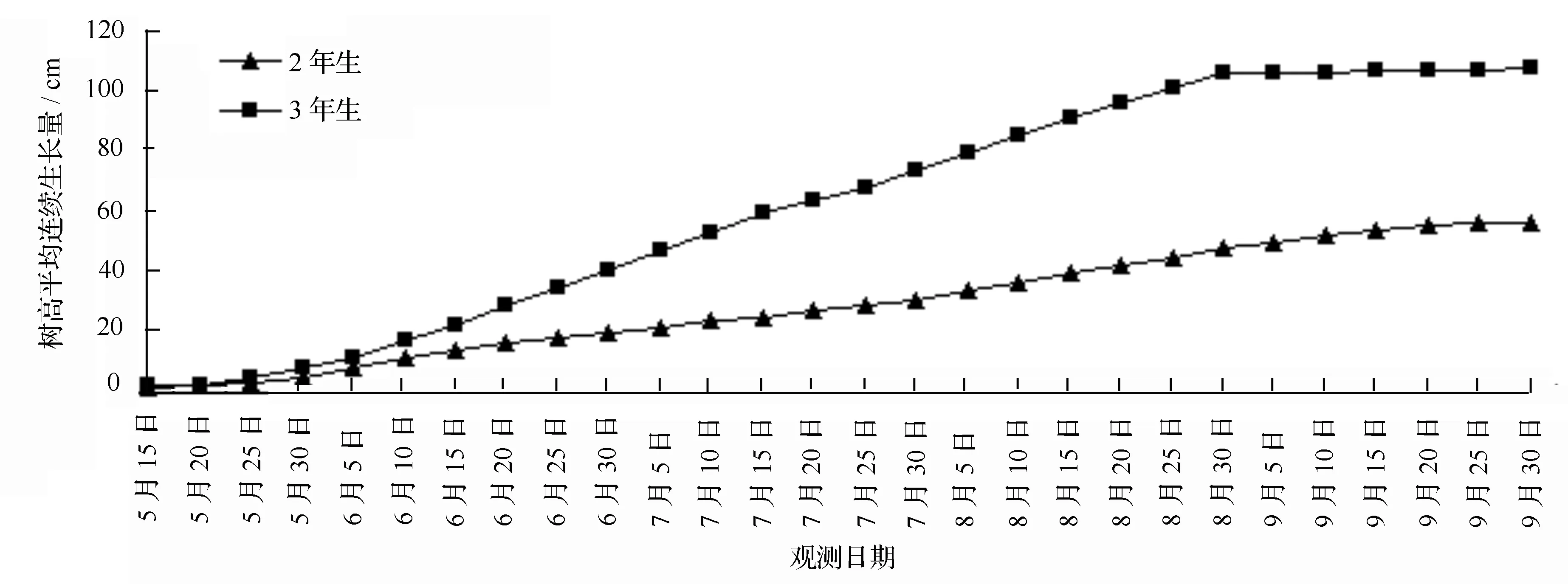
图1 日本落叶松×长白落叶松杂种无性系每5 d树高平均连续生长量
2.4.2 树高平均净生长量
为掌握林木在1年中具体的生长动态,图2给出了2、3年生57个日本落叶松×长白落叶松杂种无性系树高每5 d内的平均净生长量。可以看出,每5 d内树高的净生长量差异较大。

图2 日本落叶松×长白落叶松杂种无性系每5 d树高平均净生长量
在1个年生长周期中,2年生日本落叶松×长白落叶松杂种无性系有两个较明显的速生期。从5月15日—5月30日,林木生长缓慢,每5 d的平均净生长量仅为1.083 cm,生长期为15 d,占全年高生长量的5.95%。从5月30日—6月15日,为持续快速生长的第1个速生期,每5 d的平均净生长量为3.01 cm,生长期为15 d,占全年高生长量的16.53%。6月15日—7月30日为生长平稳期,每5 d平均净生长量为1.836 cm。随后,林木生长进入第2个速生期,持续时间为整个8月,每5 d平均净生长量为2.808 cm,生长期为30 d,占全年树高生长量的30.84%。进入9月后,生长逐渐减缓,直至封顶停止生长。
图2表明,3年生日本落叶松×长白落叶松杂种无性系的生长节律与2年生无性系的大体相同,亦有两个较明显的速生期,但3年生无性系第1个速生期的持续时间要长于2年生无性系,其具体的生长节律如下:5月15日—6月5日,林木生长较慢,每5 d的平均净生长量仅为2.408 cm,生长期为20 d,占全年高生长量的8.97%。从6月5日—7月15日,为第1个速生期,也是全年生长最快的时期,每5 d的平均净生长量为5.954 cm,生长期为40 d,占全年高生长量的44.35%。7月下旬相对而言为短暂的生长缓慢期。接着林木生长进入第2个速生期,从7月底至8月底,每5 d平均净生长量为5.343 cm,生长期为35 d,占全年树高生长量的33.83%。进入9月份后,林木生长速度锐减,直至封顶停止生长。

表5 3年生杂种无性系不同观测时期树高增长秩次相关
3 结论与讨论
辽东地区的日本落叶松×长白落叶松杂种无性系各物候因子在无性系水平上的分化较明显,从变异系数来看,芽完全展叶(主、侧枝)和抽新稍的变异系数要小于芽膨大(主、侧枝)、芽开始展叶(主、侧枝)、封顶和生长期的变异系数,造成此现象的原因可能是芽膨大、芽开始展叶和封顶等物候因子对气候因子的反应较为的敏感,致使无性系间出现较大的变异。
对2、3年生日本落叶松×长白落叶松57个杂种无性系每5 d定株调查观测树高,对树高的方差分析表明,自5月15日以来,各无性系在树高上的差异均达极显著水平,直至生长停止。表明无性系间的生长差异是存在的,这里有遗传上的差异,亦有对环境适应性的差异。2年生无性系的遗传变异系数、遗传方差占总方差百分率和无性系重复力等遗传参数随时间的变化大致可分为3个阶段,呈高—低—高的变化趋势;3年生时,各遗传参数随时间的变化大致可分为两个阶段,呈高—低的变化趋势。
相关分析表明,不同时期的树高相关性均达极显著水平,相关系数表现出随时间的增长而持续上升的趋势,8月中旬起,秩次相关系数高且趋于稳定,2年生时为0.90~0.99,3年生时达0.96~0.99,这与许少娥等[12]对杉木的研究结果相类似。
在1个年生长周期中,2、3年生的日本落叶松×长白落叶松杂种均有2个较明显的速生期。2年生无性系的第1个速生期从为5月30日—6月15日,第2个速生期为整个8月份;2个速生期的生长时间计约45 d,占全年生长时间的33.33%,但期间树高的生长量占全年高生长量的47.37%。3年生无性系的第1个速生期为6月5日—7月15日,第2个速生期为7月底至8月底;两个速生期的生长时间约75 d,占全年生长时间的55.56%,但树高生长量占全年高生长量的78.18%。因此,综合2、3年生杂种无性系的生长节律来看,在1个年生长周期中,日本落叶松×长白落叶松杂种的速生期有两个,分别为6月初至7月中旬和整个8月份。
根据2、3年生林木每5 d平均连续生长量趋势图可知,林木年生长进程可划分为生长前期、生长中期和生长后期3个时期,生长趋势为前期慢,中期快,后期慢,此结论与曹健康[13]对光皮桦幼林生长节律的研究结果一致。生长前期受自身条件(叶面积较小)和外界环境(气温低)的影响,林木生长的十分缓慢。进入生长中期后,林木本身的生物量不断累积,叶面积增大同时外界气温开始增高,林木表现出高增长状态。进入生长后期后,林木受自身遗传特性的影响及外界条件的限制,净生长量逐渐减少直至生长停止。因此,在日本落叶松×长白落叶松杂种无性系的年生长过程中,应做好生长中期特别是6月初至7月中旬和8月份两个速生期的抚育管理及施肥工作,以促进林木的生长。在生长后期,应停止施肥,以促进林木木质化,提高林木质量,使林木安全越冬。
[1] 马常耕,孙晓梅.我国落叶松遗传改良现状及发展方向[J].世界林业研究,2008,21(3):58-63.
[2] 孙晓梅,张守攻,王笑山,等.生长调节剂对落叶松杂种生根和幼苗生长的影响[J].北京林业大学学报,2006,28(2):68-72.
[3] 孙晓梅,张守攻,周德义,等.落叶松种间及种内和种间杂种家系间物候变异与早期选择[J].林业科学,2008,44(1):77-84.
[4] 张含国,张成林,兰士波,等.落叶松杂种优势分析及家系选择[J].南京林业大学学报:自然科学版,2005,29(3):69-72.
[5] Hansen J K, Roulund H. Genetic parameters for spiral grain, stem form, pilodyn and growth in 13 year old clones of sitka spruce (Piceasitchensis)(Bong.) Carr[J]. Silvae Genetica,1996,46(2/3):107-113.
[6] Hai P H, Jansson G, Harwood C, et al. Genetic variation in growth, stem straightness and branch thickness in clonal trials of Acacia auriculiformis at three contrasting sites in Vietnam[J]. Forest Ecology and Management,2008,255:156-167.
[7] Hai P H, Harwood C, Kha L D, et al. Genetic gain from breeding Acacia auriculiformis in Vietnam[J]. Journal of Tropical Forest Science,2008,20(4):313-327.
[8] 黄少伟,谢维辉.实用SAS编程与林业试验数据分析[M].广州:华南农业大学出版社,2001.
[9] 孙晓梅,张守攻,候义梅,等.短轮伐期日本落叶松家系生长性状遗传参数的变化[J].林业科学,2004,40(6):68-74.
[10] 丁振芳,王景章,方海峰,等.日本落叶松家系早期选择技术[J].东北林业大学学报,1997,25(3):65-67.
[11] 张含国,李希才,袁桂华,等.长白落叶松生长和材质性状早晚相关及早期选择[J].东北林业大学学报,1996,24(5):12-18.
[12] 许少娥,林绪平,胡德活,等.杉木优良家系苗高生长节律分析[J].中南林学院学报,2001,21(2):29-33.
[13] 曹健康,方乐金.光皮桦幼林生长节律的观察研究[J].林业科学研究,2006,19(3):404-407.
1) “十二五”国家科技支撑项目(2012BAD01B01)资助。
赖猛,男,1986年8月生,林木遗传育种国家重点实验室(中国林业科学研究院林业研究所),博士研究生。
孙晓梅,林木遗传育种国家重点实验室(中国林业科学研究院林业研究所),研究员。E-mail:xmsun@forestry.ac.cn。
2013年5月29日。
S722.3+3
Growth Rhythm of YoungLarixkaempferi×LarixolgensisHybrid/Lai Meng, Sun Xiaomei, Zhang Shougong(State Key Laboratory of Tree Genetics and Breeding, Research Institute of Forestry, Chinese Academy of Forestry, Beijing 100091, P. R. China)//Journal of Northeast Forestry University.-2014,42(4).-11~15,31
责任编辑:潘 华。
We observed and measured two and three years old phenological traits and growth rhythm after afforestation of 57Larixkaempferi×Larixolgensishybrid clones in Dagujia Forestry Station in Qingyuan County, Liaoning Province. The results of analysis phenological variation show that the phonological traits are distinctively differentiated among clones. We used variance analysis and rank correlation analysis, respectively, to analyze variation of height among clones at different times and relationship of height between different times. The height is significantly different among clones at different times. The age trends of genetic variation coefficients, genetic variance accounting for the percent of total variance and repeatability of two years old hybrid clones for height can be divided into three stages. The age trends of genetic parameters of three years old hybrid clones for height can be divided into two stages. The height has strongly positive genetic correlations with different times, and the correlation coefficients are increased with aging. During one year growing period, the height has two obviously living period, one is from early June to mid-July and the other from July 25 to August 30.
