园林树木树冠不同部位的蒸腾速率1)
2014-07-27代新竹郭太君
代新竹 林 萌 郭太君 黄 勇
(长春市动植物公园,长春,130022) (吉林农业大学)
园林树木树冠不同部位的蒸腾速率1)
代新竹 林 萌 郭太君 黄 勇
(长春市动植物公园,长春,130022) (吉林农业大学)
以吉林农业大学校园内的6种园林树木为研究对象,将树冠由外向内分为表层(距树冠表面10 cm),外层、中层和内层(各占剩余部分的1/3)。分别取上述各层的中间部位叶片(记为表层A、外层B、中层C、内层D),测定单位叶面积蒸腾速率的变化规律,探讨园林树木蒸腾速率测定的合理取样部位。结果表明:9种园林树木单位叶面积蒸腾速率的变化,均为由树冠表层向树冠内层顺次降低。表层A单位叶面蒸腾速率(Tr)是全株平均值的1.3倍,Tr(外层B)∶Tr(全株平均值)≈1∶1。因此,在测定园林树木单位叶面积蒸腾速率时,以外层B或与中层C偏向外层B处的叶片作为取样部位更为合理。
园林树木;蒸腾速率;取样部位
Journal of Northeast Forestry University.-2014,42(7).-24~26,55
We studied the variation of transpiration rate and light intensity of unit leaf area of six kinds of landscape trees cultivated in Jilin Agricultural University. We divided the crown of each tree outside-in and named layer into four parts of 0-10 cm to the crown surface (Part A), outer part (Part B), middle part (Part C) and inner part (Part D), and selected the leaves in the middle position of four parts. The variation of transpiration rate and light intensity of unit leaf area of nine kinds of landscape trees decreased from Part A to Part D. Transpiration rate in Part A of unit leaf area is 1.3 times as large as that of the average of the entire plant’s. The ratio of the average of the entire plant and the mean value of Part B is close to 1∶1. Therefore, it is reasonable to select the leaf in Part B or Part C but close to Part B as experimental materials for evaluating the transpiration rate of unit leaf area of the landscape trees.
Keywords Landscape trees; Transpiration rate; Sampling location
随着城市化的急剧发展,城市用地日益紧张,如何在城市有限的土地资源上通过优化城市绿地的空间布局和结构,充分发挥绿地的综合功能,协调城市发展与环境建设的关系,实现城市可持续发展,成为亟待解决的问题[1]。城市园林绿地不仅是城市功能的重要组成部分,同时,城市绿地的植物还具有美化、改善和保护城市环境的功能,在城市生态系统中具有不可替代性,已经成为景观生态学、城市园林生态学以及环境科学的热点[2]。
影响城市绿地生态效应的特征因素,主要包括绿地面积、绿地形状(长宽比、高度、边界曲度、周长、面积比等)、绿地景观结构、绿地内部组成、植被指数、生物量等景观生态因子[2]。由于园林植物种类自身属性不同,构成了城市绿地景观和生态的多样性。为了更好地发挥园林植物的生态功能,人们从不同的角度评价和筛选园林绿化树种。比如对园林树木增湿降温生态功能的评价和优良资源的筛选,取样测定部位基本是利用树冠边缘叶片的蒸腾速率,来估算增湿降温的生态效应[3-6]。由于树冠质地(如郁闭度)的不同,树冠表面和树冠内部的光照条件等环境因子会发生变化,利用树冠边缘叶片的蒸腾速率进行估算全株的增湿降温生态效应的方法,仍有待商榷。因此,文中以9种园林树木为研究对象,试图通过对树冠不同部位叶片蒸腾速率变化规律的研究,并结合试材的生活型和树冠投影系数,力求探讨园林树木蒸腾速率测定的合理取样部位,为筛选生态效益优良的园林树木提供依据。
1 试验地概况
试验在吉林农业大学校园内进行。吉林农业大学位于吉林省长春市东南部,北纬43°80′~43°82′,东经125°40′~125°42′,属温带大陆性季风气候,四季分明。年平均降水量568.5 mm,年平均气温4.8 ℃,最高温度39.5 ℃,最低温度-39.8 ℃,日照时间2 688 h。
2 材料与方法
试验材料及其基本特征见表1。
将树冠由外向内分为表层、外层、中层和内层(记为表层A、外层B、中层C、内层D)(图1)。表层为距树冠表面10 cm处,外层、中层和内层各占剩余部分的1/3,分别取上述各层的中间部位叶片测定蒸腾速率(Tr)。
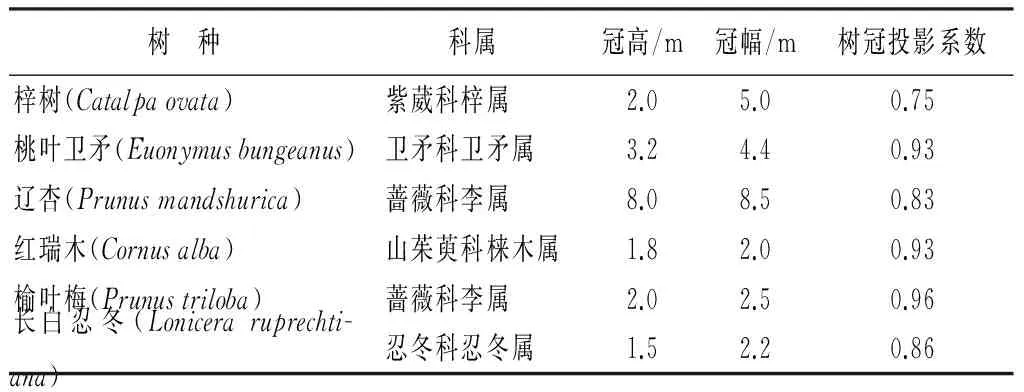
表1 试验材料基本特征

图1 树冠分层及测定点分布模式图
测定时间为7月下旬,为排除气象因素干扰,测定时间均安排在天气晴朗的条件下进行。测定仪器采用英国PP Systems国际有限公司生产的TPS-1便携式光合作用测定系统。在自然光照条件下,为保证每个时刻和测定数据的准确性,测定时分两个阶段进行。第一阶段与第二阶段测定的树种顺序相反。每天测定3个树种。每日测定时间从08:00—18:00,每隔2 h测定1次,每种树木选择长势相似的3株,每个测定点选取3枚叶片。光照强度的测定与蒸腾速率测定同时进行。结果取2个阶段的平均值。
投影系数的测定采用数码相机拍照结合Photoshop图像处理软件计算的方法得出,具体方法与柏军华等[7]报道相同。
3 结果与分析
3.1 树冠不同部位单位叶面积蒸腾速率的变化
6个树种单位叶面积蒸腾速率的变化规律表现为(表2、表3),同一树种树冠不同部位单位叶面积蒸腾速率的变化,均呈现由表层向内层逐渐下降的趋势,且树种间相差较大。如长白忍冬表层A蒸腾速率是内层D的1.3倍,桃叶卫矛则高达3.1倍;不同树种单位叶面积蒸腾速率的变化较大,如梓树最高为6.47 mmol·m-2·s-1,辽杏最低,为2.39 mmol·m-2·s-1,二者相差4.08 mmol·m-2·s-1;无论是同一树种各测定部位与其平均值比较,还是6个树种蒸腾速率平均值与各测定部位平均值比较,均表现出与外层B比较接近,而表层A为平均值的1.3倍。

表2 不同树种树冠不同部位单位叶面积蒸腾速率的变化

为了对比分析同一树种不同部位,以及不同树种相同部位的蒸腾速率相对于平均值的偏离程度和离散度,以及差异显著性。对6种园林树木种内和种间不同部位的蒸腾速率进行了标准差(SD)以及变异系数(CV)的统计分析表明(表2):树冠不同部位蒸腾速率的变化种内大于种间,如SD在种间变化在0.27(长白忍冬)~1.35(桃叶卫矛),平均0.87;种内在1.24(内层D)~1.79(表层A),平均为1.47。变异系数种间为0.10(长白忍冬)~0.47(桃叶卫矛),平均为0.25;种内变异系数为0.32~0.47,平均为0.40。乔木生活型树冠不同部位蒸腾速率标准差(SD)以及变异系数(CV)大于灌木生活型的。如试材中梓树、桃叶卫矛和辽杏为乔木,SD和CV平均值分别为1.18和0.35,变化幅度分别在0.92~1.35和0.20~0.47;而红瑞木、榆叶梅和长白忍冬这3种灌木SD和CV平均值分别为0.57和0.16,变化幅度分别为0.27~0.77和0.10~0.22。
各树种不同部位之间蒸腾速率的多重比较显示(表2),表层A与其他部位比较,除桃叶卫矛外层B和红瑞木中层C外,其他各部位均显著或极显著低于表层A;外层B与中层C和内层D之间,差异明显和不明显基本占一半。说明表层A与树冠其他部位的蒸腾速率差异已经达到极显著水平。方差分析结果(表3)表明,6个树种在不同的树冠部分对单位叶面积蒸腾速率的影响达到极显著水平(P<0.01)。
3.2 树冠不同部位相对光照强度的变化
树冠不同部位光照强度的变化,各树种均呈由外至内依次明显降低的趋势,如外层B、中层C、内层D的光照强度仅为表层A的21.87%~39.56%。树冠冠投影系数与树冠内部的光照强度相对较高,树冠冠投影系数则相对较低,如梓树、辽杏和长白忍冬树冠投影系数较小,如辽杏外层B、中层C、内层D的光照强度占表层A的68.82%、56.99%和48.39%;而桃叶卫矛分别占20.19%、12.26%和10.51%。不同树种树冠不同测定部位之间,除桃叶卫矛和长白忍冬的中层C和内层D之间差异不明显外,其他均达到极显著水平(表4)。方差分析结果(表5)表明,6个树种在不同的树冠部分的光照强度之间达到极显著差异(P<0.01)。
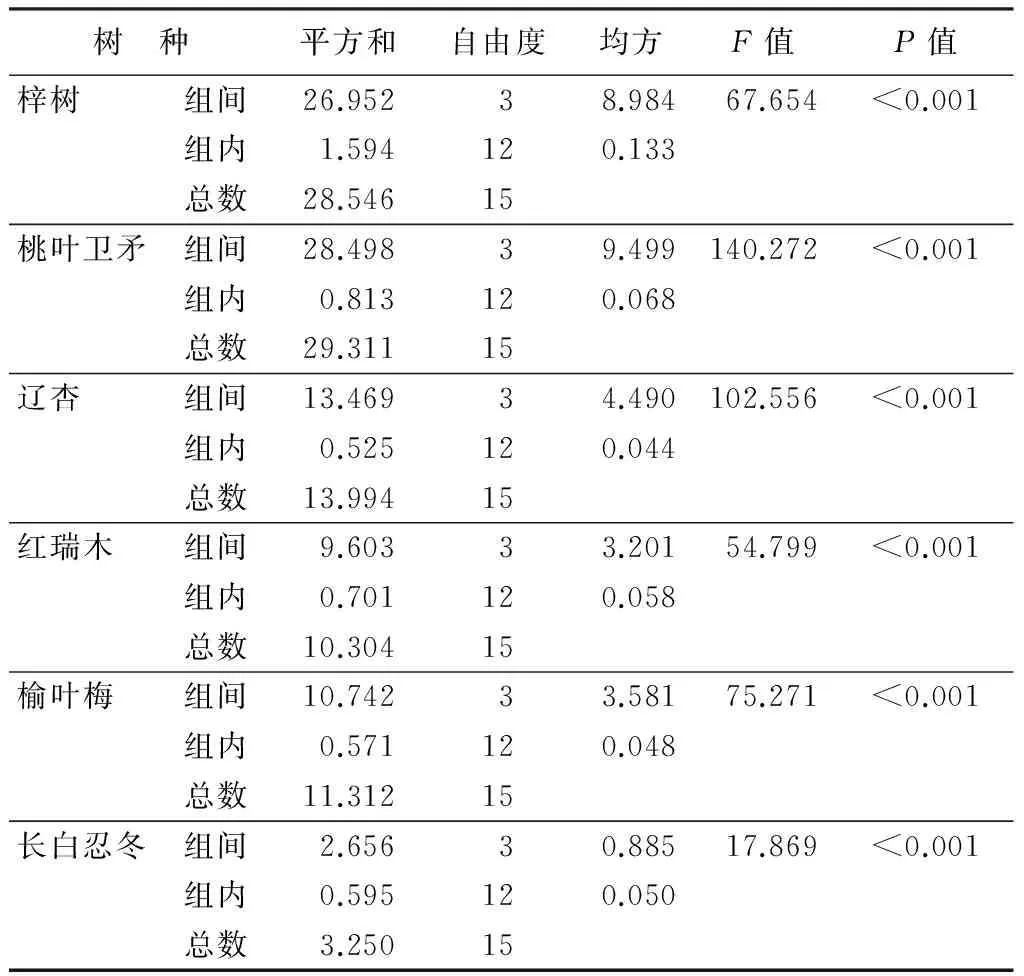
表3 各树种蒸腾速率方差分析结果
注:P<0.01为极显著差异。

表4 各树种各层光照强度的变化
注:同列不同小写字母表示差异显著(P<0.05);同列不同大写字母表示差异极显著(P<0.01)。

表5 各树种光照强度方差分析结果
注:P<0.01为极显著差异。
4 结束语
由于树冠质地(如枝条长短、数量、叶片数量和大小等)直接影响树冠内部的微生境,如光照、温度和风速等,致使树冠不同部位叶片蒸腾速率存在较大的差异。树冠投影系数是树冠质地的综合反映,投影系数大则树冠内部的光照强度及风速等相对减弱,叶片蒸腾速率也会随之发生变化。因此,合理的取样测定部位直接影响蒸腾速率测定结果的代表性和准确性。在进行园林树木增湿降温生态功能评价时,有关以基本测定树冠外缘南侧叶片的蒸腾速率来计算增湿降温环境生态效应[3-6,8-11]已有报道。由于不同树种树冠投影系数的差异,树冠表面和内部的微生境会发生变化,导致树冠不同部位叶片蒸腾速率的差异明显。如树冠枝叶稀疏的辽杏树冠冠投影系数较小,树冠内部的光照强度相对较高,表层A的蒸腾速率为全株平均值的1.3倍;树冠投影系数较高的桃叶卫矛树冠内部的光照强度相对较低,表层A蒸腾速率较全株平均值高达1.5倍。本研究认为,在测定园林树木的蒸腾速率时,取树冠外层B或与中层C偏向外层B处的叶片作为取样部位,基本能够代表树冠的整体蒸腾水平。这与前文报道的园林树木固碳释氧生态功能评价的取样部位结果相同[12]。
不同生活型的树种,树冠不同部位的蒸腾速率及其与平均值偏离程度和离散度具有差异,乔木生活型大于灌木生活型,其原因可能是植物本身长期适应所处环境而形成的一种特性。这种对环境条件的适应性,可能是乔木生活型树冠不同部位的蒸腾速率,受光照强度、风速、温度和空气湿度等的影响大于灌木生活型所致。如在自然群落中,乔木多处于群落垂直结构的上层,灌木多处于群落垂直结构的下层,其生存的环境条件如光照强度、风速、温度和空气湿度等明显不同,这些环境条件是影响增湿降温量变化的重要生态因子。
[1] 郑西平.北京城市绿地系统功能评价与发展策略[D].北京:北京林业大学,2011.
[2] 苏泳娴,黄光庆,陈修治,等.城市绿地的生态环境效应研究进展[J].生态学报,2011,31(23):7287-7300.
[3] 陈娟,史红文,廖建雄,等.武汉市春季49种园林植物的光合和蒸腾特性研究[J].河南大学学报:自然科学版,2013,43(1):62-68.
[4] 陈少鹏,庄倩倩,郭太君,等.长春市园林树木固碳释氧与增湿降温效应研究[J].湖北农业科学,2012,51(4):750-756.
[5] 郑鹏,史红文,邓红兵,等.武汉市65个园林树种的生态功能研究[J].植物科学学报,2012,30(5):468-475.
[6] 陆贵巧,谢宝元,谷建才,等.大连市常见绿化树种蒸腾降温的效应分析[J].河北农业大学学报,2006,29(2):65-67.
[7] 柏军华,王克如,初振东,等.叶面积测定方法的比较研究[J].石河子大学学报:自然科学版,2005,23(2):216-218.
[8] 王忠君.福州国家森林公园生态效益与自然环境旅游适宜性评价研究[D].北京:北京林业大学,2004.
[9] 廖容,崔洁,卓春丽,等.成都市32种立体绿化植物降温增湿效应比较研究[J].江苏农业科学,2012,40(6):178-181.
[10] 焦绪娟,赵文飞,张衡亮,等.几种绿化树种降低城市热岛效应的研究[J].江西农业大学学报,2007,29(1):89-93.
[11] 莫健彬,王丽勉,秦俊,等.上海地区常见园林植物蒸腾降温增湿能力的研究[J].安徽农业科学,2007,35(30):9506-9507,9510.
[12] 林萌,郭太君,代新竹.9种园林树木固碳释氧生态功能评价[J].东北林业大学学报,2013,41(6):29-32.
Transpiration Rate of Landscape Trees in Different Positions/
Dai Xinzhu(Changchun Zoological and Botanical Park, Changchun 130022, P. R. China); Lin Meng, Guo Taijun, Huang Yong(Jilin Agricultural University)//
代新竹,女,1976年6月生,长春市动植物公园,高级工程师。
2013年6月24日。
Q945.17+2
1) 吉林省科技发展计划项目(20095038)。
责任编辑:任 俐。
