整合生物量及碳汇的帽儿山地区兴安落叶松31年生种源选择1)
2014-07-27朱航勇张含国贾庆斌
朱航勇 张含国 张 振 姚 宇 贾庆斌
(林木遗传育种国家重点实验室(东北林业大学),哈尔滨,150040)
整合生物量及碳汇的帽儿山地区兴安落叶松31年生种源选择1)
朱航勇 张含国 张 振 姚 宇 贾庆斌
(林木遗传育种国家重点实验室(东北林业大学),哈尔滨,150040)
林木遗传改良的主要目的是选育生产力大的良种,但是随着木材市场和全球气候变化,林业生产更加关注生物量和碳汇功能。对帽儿山31年生兴安落叶松生物量和碳储量的分配模式,以及同生长性状相关性、异俗生长模型和遗传力进行分析,并初步选择了生物量和碳储量大的优良种源。兴安落叶松树干(含树皮)生物量、碳储量分别占地上部分总体的74.17%、62.50%,伐倒木、立木材积同生物量呈极显著相关,碳质量分数从大到小依次为:树叶、树皮、树枝、树干,胸径处碳质量分数与总体平均碳质量分数相关系数为0.615,显著水平为0.058。对于生物量估算各类异速生长模型中,Y=aDbHc和Y=aDb两种模型较适宜兴安落叶松的生物量估算。两个模型对树干生物量和地上部分生物量预测误差小于5%,预测准确性较好。将实验林数据带入Y=aDb模型中,生物量与碳储量种源遗传力约为0.36,最优种源为友好林业局,生物量和碳储量分别比整体平均值高40.38%、43.92%,比最差中央站种源高130.66%和130.44%。
兴安落叶松;种源选择;生物量;碳汇
Journal of Northeast Forestry University.-2014,42(7).-5~10,59
We analyzed the partitioning model of the biomass and carbon stock of 31 years old Dahurian Larch (Larixgmelinii(Rupr.) Rupr.), their relationship with the growth trait, selection and primary selected provenance for biomass and carbon sequestration. The stem (bole wood and bark) ofL.gmeliniiaccounted for 74.17% of biomass and 62.50% of carbon aboveground. The correlation between felled and standing timber volume with biomass was highly significant. The sequence of carbon concentration for different organ was foliage>bark>branch>bole wood. The correlation coefficient between carbon concentration at diameter at breast height (DBH) level and average of total aboveground was 0.615, with the significant level of 0.058. The models ofY=aDbHcandY=aDbare most suitable for estimating the biomass ofL.gmeliniiamong all the allometry model with 5% of standard error. When the inventory data of provenances test is taken in theY=aDbmodel, the provenance heritability of biomass and carbon stock is 0.36. The best provenance is Youhao, biomass and carbon stock exceed the test average by 40.38% and 43.92%, and exceed the worst provenance (Zhongyangzhan) by 130.66% and 130.44%, respectively.
Keywords Dahurian Larch (Larixgmelinii(Rupr.)Rupr.); Provenance; Biomass; Carbon sequestration
兴安落叶松(Larixgmelinii(Rupr.) Rupr.)是我国寒温带针叶林的主要建群种,林分面积大,蓄积量多,分别占我国寒温带有林地面积和蓄积量的55%和75%[1-2]。兴安落叶松具有木材用途广、适应性强、速生等优点,现已成为东北地区森林经营和人工造林的主要树种之一[3]。国内对于兴安落叶松育种研究较早,在种源区划、地理变异及优良种源选择取得很多成就。通过选育优良种源造林,显著提高了林分的生产力[4-7]。但是,这些研究往往只关注材积的生长,而忽视木材干质量(生物量)等性状。近年来随着对树木的综合利用程度的提高,影响着木材市场的走向。对于造纸业,生物量与纸浆产量的相关性较材积更好,因此,相对于材积而言,市场越来越关注生物量[8]。随着全球气候变化和京都议定书的签订,使森林碳汇与各国的政治、经济利益密切相关。在清洁发展机制下,造林和再造林可以在森林碳汇交易平台销售碳排放指标[9]。因此,从生物量和碳汇角度,选育林木良种进行造林有着重要的生态和经济意义。
从目前国外的研究来看,桉树的育种项目引入生物量和碳汇这两个性状后,对原有的经济性状的相对权重无显著的影响[10];辐射松的碳汇研究结果表明,通过选育林木良种造林可以显著的提高碳汇价值和经济效益[11]。
生物量研究需要进行破坏性取样。样本的取得、处理,不仅需要投入大量的资金和劳动力,而且对科研试验林也会造成极大的破坏。因此,通常采用异俗生长方程对树木生物量进行估算[12-14]。从育种研究的角度,对不同种源和家系单独建立异俗生长方程,需要在苗期和树木年龄较小时进行,这种方法周期长、投入大。目前,异俗生长方程主要应用于生态领域,利用树种通用异俗生长方程对森林调查数据进行大范围的估算。虽然通用异俗生长方程,对不同地区和立地条件会产生估测误差,但是这种估测误差在大范围的地区和立地条件下被中和[15]。而对同一育种试验林建立通用方程估算生物量的误差是否影响选育结果,目前还有没有研究报道。
本文通过对31年生兴安落叶松种源实验林进行研究,分析31年生兴安落叶松的生物量、碳储量及其分配情况,验证通用异俗生长方程准确性。并对碳汇和生物量两个性状整合到兴安落叶松育种项目,尝试分析生物量和碳储量的遗传力,以便进行优良种源的选择。
1 研究地自然概况
研究地点选择在东北林业大学帽儿山实验林场,该林场具备典型的中国东北地区的森林类型和景观形态。平均海拔300 m,坡度为10°~15°,土壤类型为暗棕壤。典型的大陆温带季风型气候,夏季温暖湿润,冬季寒冷干燥。年降水量为600~800 mm,主要集中在7—8月份,这两个月占全年降水量的80%左右,年蒸发量为884 mm。年平均温度为2.8 ℃,1月份平均气温为-31 ℃,7月份平均气温为32 ℃。>5 ℃的积温为2 897 ℃,无霜期为120~140 d,光照时间1 850 h,平均风速1.5 m/s[16]。
兴安落叶松种源试验林,采用完全随机区组设计,5次重复,17个种源,小区株数70株,定植时间:1983年,株行距为1.5×2.0 m,1995年进行隔行去行间伐,2001年进行隔1株去2株间伐[4,17]。
2 研究方法
2.1 取样和测量方法
2010年7月末对31年生的兴安落叶松种源试验林进行生长调查。在最优、中等、较差三个种源区内,按照径级总共选择10株树木,三个种源都含有优势木、中等木、被压木。解析木自地表伐倒后,测量树高。树高≥15 m,则2 m截段,树高<15 m,则1 m截段,分别测量每段的鲜质量。两端和中央位置的带皮直径测量后,在木段中央处截取一个5 cm厚的圆盘,同时在伐倒木基部、胸径处截取5 cm厚圆盘带回实验室;树冠按照长度平均分为三层,将每层树枝分别称质量,选取标准枝称质量后,将其枝叶分离,分别测定鲜质量,取样带回实验室。
样品在105 ℃杀青后,在70 ℃下烘干至衡质量,称质量并记录;带皮圆盘样品烘干后,将树皮剥离,分别称质量。通过烘干前后的质量,计算样品含水率。由样品的含水率和各部分鲜质量计算出各部分以及整株树木的生物量。
全部样品称质量后,通过带锯床和粉碎机逐级对取样进行粉碎,取2~3 g样品经Retsch MM400型球磨机研磨,粉末装入纸袋以备碳质量分数分析。
将烘干至衡质量的样品,在Jena HT1300型固体模块内,1 200 ℃高纯氧下充分燃烧,燃烧后的气体经过Jena Multi C/N 2100S型TOC分析仪检测,得到各部分的碳质量分数,经过计算得到各部分碳储量。
伐倒木材积用各木段带皮直径,按分区求积法计算;立木材积运用树高和胸径,按形数法计算。
2.2 数据分析
异俗生长模型采用:Y=aXb;Y=aDbHc。其中:Y为生物量或者碳储量;公式Y=aXb中,X为材积(V)、立木材积(Sv)、胸径(D)、胸径与树高(H)的乘积(D×H)、胸径的平方和树高(D2×H)的乘积;应用PROC NLIN(SAS Institute 1992)进行分析。直接采用非线性回归而不进行数据对数转换,可以省略一系列校正参数[18]。
最优模型的选择:
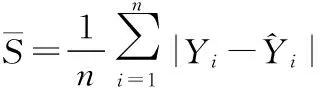


遗传力估算:
hi=VA/VP;

线性模型:
Yijt=μ+Pi+Bj+PBij+eijt。
式中:Yijt为第i个区组第j个种源第t的观测值,μ为总体平平均值,Bj为区组,Pi为家系,PBij为种源和区组的交互作用,eijt为误差。
3 结果与分析
3.1 生物量与生长量
兴安落叶松生长量和生物量(见表1)。通过对表中数据进行比较和相关性分析,结果表明:兴安落叶松树干生物量(不含树皮)占地上部分生物量的比率变异范围为57.04%~72.24%,平均值为63.93%、标准差为5.70,变异系数为8.91%;树皮生物量占地上部分生物量的比率变异范围为5.65%~16.60%,平均值为10.23%、标准差为3.51,变异系数为34.28;树枝生物量占地上部分生物量的比率变异范围为13.36%~28.72%,平均值为20.23%、标准差为5.20,变异系数为25.70;树叶生物量占地上部分生物量的比率变异范围为4.61%~6.79%,平均值为5.60%、标准差为0.66,变异系数为11.72。树皮和树干之和所占的生物量占地上部分生物变异范围为65.95%~81.74%,平均值为74.17%,标准差为5.49,变异系数为7.40%;含树皮的树干部分是地上部分生物量的主体,平均占地上部分生物量74.17%。胸径、树高同各部分生物量均呈显著正相关。树高和胸径同树枝占地上部分生物量比率呈显著正相关相关系数分别为0.635和0.692;树高和树冠生物量占总生物量的比重显著相关,相关系数为0.650。伐倒木材积同各部分生物量均极显著相关;立木材积与各部分生物量同样呈现极显著相关。因此,用非破坏性取样比较单株生物量大小的时候,可以用立木材积来代替生物量进行简单的定性比较。

表1 兴安落叶松生长量和生物量
3.2 兴安落叶松生物量异俗生长方程

表2 各部分生物量回归模型统计表
注:纸浆材生物量为树干生物量(不含皮)与树枝生物量之和。
Y=aDb和Y=aDbHc回归模型的参数(见表3)。两个模型预测值的相对误差小于5%的有:地上部分生物量、树干(带皮)、树干(不带皮)、纸浆材的生物量。两种模型对这些部分的生物量预测准确度较高,把预测这部分生物量的异俗生长方程用于育种研究,遗传差异不易被预测误差所掩盖,其选择结果是可靠的。而对于其它部分的方程而言,预测相对误差≥5%,遗传差异会被模型的预测误差所掩盖。树枝、树叶和树皮生物量模型的预测相对误差均大于9%,说明兴安落叶松树枝、树叶和树皮生物量很难准确估算。地上部分生物量、树干生物量、纸浆材的生物量为育种主要改良经济性状,由于对这些性状的预测准确性较好。因此,在育种研究中,可以选择这两种回归模型进行估算。

表3 各部分生物量最优模型参数统计表
3.3 兴安落叶松碳储量和碳质量分数
兴安落叶松各解析木各部分碳储量、碳质量分数均值(见表4)。通过对表中数据进行比较和相关性分析,结果表明:兴安落叶松各部分碳储量分配存在极显著差异(F值12.08,P值小于0.01),兴安落叶松树干(不含树皮)碳储量占地上部分比率变异范围为54.93%~71.60%,平均值为62.50%,标准差为6.09,变异系数为9.74;树皮碳储量占地上部分的比率变异范围为6.21%~16.85%,平均值为10.86%,标准差为3.54;变异系数32.62;树枝碳储量占地上部分比率变异范围为13.24%~29.18%,平均值为20.67%,标准差5.41,变异系数为26.18;树叶碳储量占总体比率变异范围为4.82%至7.31%,平均值为5.97%,标准差为0.79,变异系数为13.23。树高和胸径与碳储量均呈显著正相关,树高、胸径同树枝碳储量占地上部分的比率呈显著正相关,其相关系数分别为0.702和0.652。

表4 兴安落叶松各解析木各部分碳储量、碳质量分数均值
兴安落叶松各部分碳质量分数差异显著(F值10.30,P值小于0.01)。其中:树干(不含树皮)碳质量分数平均值为44.496%,树皮碳质量分数平均值为48.507%,树枝碳质量分数平均值为46.512%,树叶碳质量分数平均值为48.511%。兴安落叶松各部分碳质量分数由大到小的排序为:树叶、树皮、树枝、树干。其中树高、胸径与树皮、树叶碳质量分数成显著的正相关。对解析木木段(不含树皮)不同位置(树高)与其碳质量分数进行相关分析,相关系数为0.216,P值为0.018。随着树高的增加,碳质量分数随之增大。
由表5可知。兴安落叶松胸径处的碳质量分数和树干(不含树皮)平均碳质量分数相关显著,其相关系数为0.785,P值为0.007,可以用非破坏性取样代替平均碳质量分数进行比较。但是兴安落叶松胸径处碳质量分数与地上部分的平均碳质量分数相关系数为0.615(P值为0.058),虽然在5%显著水平下相关不显著,这可能是由于样本容量小引起的。

表5 兴安落叶松各部分碳质量分数相关分析
注:表中数值为相关系数,括号内数值为P值。
3.4 碳储量非线性回归
Tolunay.D[20]按照生物量模型建立了碳储量的回归模型。按照生物量模型相同的分析方法,选择最优的碳储量模型(见表6)。对树木地上部分、树干(不带皮)、树枝、树叶、树冠碳储量预测误差最小的模型为Y=aVb;对树干(带皮)、树皮碳储量预测误差最小的模型为Y=aDbHc。因此,同兴安落叶松碳储量高度相关的为材积性状,同生物量模型相同。材积是通过破坏性取样得到,而立木材积预测误差除对树叶预测误差最小以外,其余的预测误差的都比模型Y=aDbHc、Y=aDb大。因此,在不考虑伐倒木材积的情况下,最优碳储量模型除树叶碳储量之外,其余皆为Y=aDbHc模型。与生物量预测相同,树高不易测量,且测量误差较大,而Y=aDb与Y=aDbHc模型预测误差相差较小。因此,在研究中可根据具体情况,选择适宜模型。
兴安落叶松非线性回归预测碳储量对于不同部分预测结果相差较大(见表7),其中Y=aDb和Y=aDbHc两种模型对于地上部分和树干(带皮)碳储量预测相对误差均小于5%,预测比较准确,其它部分预测相对误差均大于9%。对比生物量模型的预测准确度,碳储量模型除树冠外,其余模型预测相对误差均增加,碳储量较生物量难以准确预测。对于地上部分碳储量,两种模型预测的相对误差分别为4.26%和3.99%,小于5%,可以用于估算地上部分碳储量。
表6 碳储量回归模型及模型统计表

表7 各部分碳储量最优模型参数统计表

3.5 遗传力估算和优良种源选择
将胸径数据带入Y=aDb模型中,得到生物量和碳储量数据。应用PROC GLM和PROC Mixed分析胸径、生物量和碳储量数据[21]。分析结果表明:种源之间胸径、生物量及碳储量差异极显著。按Dickerson法计算单株遗传力和种源遗传力[18]。胸径的单株遗传力为0.112,种源遗传力为0.356;地上部分生物量单株遗传力和种源遗传力分别为0.099、0.356;树干(不含皮)生物量单株遗传力和种源遗传力分别为0.101、0.356;制浆用材生物量单株遗传力和种源遗传力分别为0.098、0.356;地上部分碳储量的单株遗传力和种源遗传力分别为0.098、0.356。虽然其它数值是胸径的函数,但是单株遗传力较胸径略有下降,单株遗传力基本维持在0.10上下,种源遗传力往往受实验设计影响比较大[22],种源遗传力为0.356。生长最好的友好种源,地上部分生物量平均单株生物量为259.5 kg,超出最差种源中央站种源(116.3 kg)123.12%,超出总体平均值42.64%;树干(不含皮)生物量友好种源(160.0 kg)超出最差中央站种源(75.4 kg)112.20%,超出总体平平均值(114.0 kg)40.38%;纸浆用材生物量友好种源(217.8 kg)超过最差中央站种源(94.3 kg)130.66%,超出总体平均值(151.1 kg)44.14%;碳储量友好种源(119.6 kg)超出最差中央站种源(51.9 kg)130.44%,超过总体平均值43.92%。因此,选择优良兴安落叶松种源造林可以极大提高生产力和固碳量。
4 结论与讨论
兴安落叶松树干(含树皮)生物量、碳储量分别占地上部分总体的74.17%、62.50%,伐倒木、立木材积同生物量呈极显著相关,碳质量分数从大到小依次为:树叶、树皮、树枝、树干,胸径处碳质量分数与总体平均碳质量分数相关系数为0.615,显著水平为0.058。对于生物量估算各类异速生长模型中,Y=aDbHc和Y=aDb两种模型较适宜兴安落叶松的生物量估算。两个模型对树干生物量和地上部分生物量预测误差小于5%,预测准确性较好。将实验林数据带入Y=aDb模型中,生物量与碳储量种源遗传力约为0.36,最优种源友好,生物量和碳储量分别比整体平均值高40.38%、43.92%,比最差中央站种源高130.66%和130.44%。
从生物量和碳储量回归模型的拟合效果,最优的模型是材积,即生物量和碳储量是材积的函数,因此,整合碳汇育种的兴安落叶松项目可行。这与Whittock S.P.等[10]对桉树的研究结果是一致的,不会影响的关键经济性状的权重。由于本研究所用的材积是伐倒木材积,虽然立木材积与伐倒木材积显著相关,且相关系数为0.997。但是通过立木材积拟合的模型预测误差,大于通过树高和胸径模型或单一胸径模型的预测误差。对于大兴安岭兴安落叶松人工林研究,15~26年生的兴安落叶松人工林碳汇能力最强,树干碳库占乔木碳库总储量的54.3%~73.9%[1]。本研究取材于生长在帽儿山地区的兴安落叶松,林龄为31 a,处于树干碳汇能力由高峰开始下降的阶段,树干碳储量占地上部分碳储量的比率变异范围为54.93%~71.60%。在这一时期,对种源进行选择的效果要比早期好。从白云杉的生物量遗传力研究结果来看,单株和家系遗传力变异范围分别为0.186~0.359、0.352~0.536,遗传力处于中等水平。因此,树木生物量分配模式与树木的生长效率有关[23],选择与育种可以改变生物量的分配[12]。通过选育不但可以提高树木的生物量,而且可以提高树木的生长效率。
对于生物量和生物量分配,胸径和树高的作用是不同的,从生物量最优方程以及方程参数中可以发现,胸径相对于树高与生物量呈现更大的正相关。因此,以生物量改良为目的的育种研究,应侧重于胸径的改良。现在的生物量改良往往侧重于树高,树高主要影响生长效率(即树干部分占总体生物量的比率)。对于给定胸径,树高同生长效率成正比[24];相反对于给定树高,胸径同生长效率呈反比[25]。
对于欧洲赤松的种源研究结果来看,不同纬度群体的树高、胸径、生物量差异显著,但是生物量模型的参数无显着差异,可以应用通用方程进行后续研究[26]。
本研究的碳质量分数相对较低。主要有两个因素造成的,其一,由于烘干造成挥发性碳物质的损失,其二,烘干温度低,水分相对含量较高引起的。对于长白落叶松,挥发性碳的比率为3.47%[27],海岸松65 ℃烘干和103 ℃烘干,不同组分的含碳率低0.6%~1.9%[28]。本研究采用70 ℃烘干,造成的系统误差大约在4%~5%。由于实验条件对所有样品误差一样,育种研究是对数据进行比较,由系统造成的误差对选育结果没有影响。
对于生物量和碳储量的研究。选择直接法还是间接法,以及在选择何种模型进行生物量和碳储量估算,要综合考虑研究的具体情况,选择最适宜的方法途径。本研究分析了兴安落叶松生物量异俗生长方程的估算误差,同时尝试建立了碳储量的异俗生长方程,这两类方程对于地上部分和树干(含树皮)的估算误差都小于5%,相对较小,在10%遗传增益的选择标准下,对选择结果没有影响。如果提高样品容量,降低方程的估算误差,还可以进一步提高育种选择的准确性。
按照本研究的结果,在帽儿山地区31年生兴安落叶松最优种源生物量和碳储量超过最差种源100%以上,超过总体平均值40%以上。因此,选择优良种源造林,可以大幅度提高兴安落叶松人工林的经济效益和生态效益。
[1] 齐光,王庆礼,王新闯,等.大兴安岭林区兴安落叶松人工林植被碳贮量[J].应用生态学报,2011,22(2):273-279.
[2] 孙玉军,张俊,韩爱惠,等.兴安落叶松幼中龄林的生物量与碳汇功能[J].生态学报,2007,27(5):1756-1762.
[3] 刘世荣,柴一新,蔡体久,等.兴安落叶松人工群落生量物与净初级生产力的研究[J].东北林业大学学报,1990,18(2):40-46.
[4] 杨传平,姜静,唐盛松,等.帽儿山地区21年生兴安落叶松种源试验[J].东北林业大学学报,2002,30(6):1-5.
[5] 杨传平,黄秦军,肖国志,等.兴安落叶松生长变异及早期选择[J].东北林业大学学报,1996,24(5):122-127.
[6] 杨传平,杨书文,刘桂丰,等.兴安落叶松种源试验研究(Ⅰ):地理变异的规律与模式[J].东北林业大学学报,1990,18(4):10-21.
[7] 杨传平,杨书文,夏德安,等.兴安落叶松种源试验研究(Ⅱ):最佳种源的选择[J].东北林业大学学报,1991,19(S2):68-76.
[8] Avery T E, Burkhart H E. Forest measurements[M]. New York: McGraw-Hill Book Company,1994.
[9] IPCC. The scientific basis. Contribution of working group I To the third assessment report of the Intergovern-mental Panel on Climate Change[M]. Cambridge: Cambridge University Press,2001.
[10] Whittock S P, Dutkowski G W, Greaves B L, et al. Integrating revenues from Carbon sequestration into eco-nomic breeding objectives for Eucalyptus globulus pulpwood production[J]. Annals of Forest Sci-ence,2007,64(3):239-246.
[11] Jayawickrama K J. Potential genetic gains for Carbon sequestration: a preliminary study on radiata pine planta-tions in New Zealand[J]. Forest Ecology and Management,2001,152(1/3):313-322.
[12] Rweyongeza D M, Yeh F C, Dhir N K. Heritability and correlations for Biomass production and allocation in white spruce seedlings[J]. Silvae Genetica,2005,54(4/5):228-235.
[13] Zianis D, Mencuccini M. Aboveground biomass relationships for beech (FagusmoesiacaCz.) trees in Vermio Mountain, Northern Greece, and generalised equations for Fagus sp[J]. Annals of Forest Sci-ence,2003,60:439-448.
[14] Parresol B R. Assessing tree and stand biomass: A review with examples and critical comparisons[J]. Forest Science,1999,45(4):573-593.
[15] Xiang W H, Liu S H, Deng X W, et al. General allometric equations and biomass allocation of Pinus masson-iana trees on a regional scale in southern China[J]. Ecological Research,2011,26(4):697-711.
[16] Wang C K. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J]. Forest Ecology and Management,2006,222(1/3):9-16.
[17] 鄂文峰,王传宽,杨传平,等.兴安落叶松边材心材生长特征的种源效应[J].林业科学,2009,45(6):109-115.
[18] Dickerson G E. Techniques for research in quantitative animal genetics[C]//Techniques and procedures in animal science research, New York: Am Soc Anim Sci,1969.
[19] Schmidt A, Poulain M, Klein D, et al. Allometric above-belowground biomass equations for Nothofagus pumi-lio (Poepp. & Endl.) natural regeneration in the Chilean Patagonia[J]. Annals of Forest Sci-ence,2009,66(5):513-513.
[20] Tolunay D. Carbon concentrations of tree components, forest floor and understorey in young Pinus sylvestris stands in north-western Turkey[J]. Scandinavian Journal of Forest Research,2009,24(5):394-402.
[21] SAS Institute Inc.. Statistical Analysis System (SAS): SAS procedure guide, verison 6[CP]. Third edn. Cary: SAS Institute Inc.,1990.
[22] White T L, Hodge G R. Predicting breeding values with applications in forest tree improvement[M]. Dor-drecht: Kluwer Academic Publishers,1989.
[24] Skovsgaard J P, Bald C, Nord-Larsen T. Functions for biomass and basic density of stem, crown and root sys-tem of Norway spruce (Piceaabies(L.) Karst.) in Denmark[J]. Scandinavian Journal of Forest Re-search,2011,26(11):3-20.
[25] Goodman A M, Ennos A R. A comparative study of the response of the Roots and shoots of sunflower and ma-ize to mechanical stimulation[J]. Journal of Experimental Botany,1996,47(33):1499-1507.
[26] Oleksyn J, Reich P B, Chalupka W, et al. Differential above-and below-ground biomass accumulation of Euro-pean Pinus sylvestris populations in a 12-year-old provenance experiment[J]. Scandinavian Journal of Forest Research,1999,14(1):7-17.
[27] Thomas S C, Malczewski G. Wood Carbon content of tree species in Eastern China: interspecific variability and the importance of the volatile fraction[J]. Journal of Environmental Management,2007,85(3):659-662.
[28] Bert D, Danjon F. Carbon concentration variations in the Roots, stem and crown of maturePinuspinaster(Ait.)[J]. Forest Ecology and Management,2006,222(1/3):279-295.
Integrating Biomass and Carbon Sequestration of 31-Year Dahurian Larch (Larixgmelinii(Rupr.) Rupr.) Provenance Selection in Maoershan/
Zhu Hangyong, Zhang Hanguo, Zhang Zhen, Yao Yu, Jia Qingbin(State Key Laboratory of Tree Genetics and Breeding, Northeast Forestry University, Harbin 150040, P. R. China)//
1) 国家“十二五”科技支撑项目(2011BAD37B02-1)。
朱航勇,男,1981年2月生,林木遗传育种国家重点实验室(东北林业大学),博士研究生。E-mail:zhhy504@hotmail.com。
张含国,林木遗传育种国家重点实验室(东北林业大学),教授。E-mail:hanguozhang1@sina.com。
2013年11月8日。
S722.3+3; S718.46
责任编辑:王广建。
