不同土壤水分对向日葵光合光响应的影响
2014-07-07云文丽侯王海梅李建军张
云文丽侯 琼*王海梅李建军张 超
1)(内蒙古气象科学研究所,呼和浩特010051)
2)(内蒙古巴彦淖尔市临河观象台,临河015000)
3)(内蒙古生态与农业气象中心,呼和浩特010051)
不同土壤水分对向日葵光合光响应的影响
云文丽1)侯 琼1)*王海梅1)李建军2)张 超3)
1)(内蒙古气象科学研究所,呼和浩特010051)
2)(内蒙古巴彦淖尔市临河观象台,临河015000)
3)(内蒙古生态与农业气象中心,呼和浩特010051)
以食用向日葵为试验材料,大田试验采取人为控制土壤水分在胁迫、适宜和过湿(土壤田间持水量的40%~54.9%,55%~69.9%和70%~90%)条件下,研究了向日葵3个生育期(二对真叶—花序形成期、花序形成—开花期、开花—成熟期)的光合光响应特性。结果表明:在试验设置光强条件下,各生育期净光合速率随着光合有效辐射的增加而增加,同等的光合有效辐射下净光合速率也随着土壤水分的减少依次降低,尤其是随着光合有效辐射的增大愈加明显。土壤湿度对最大净光合速率和表观量子效率的影响并不是同步的,最大净光合速率随着土壤湿度的增加而增大,而表观量子效率在一定程度的水分胁迫情况下出现最大值。不同的土壤水分含量对光补偿点和光饱和点影响不同,光饱和点随着土壤水分的增加而增加,光补偿点却相反,表明水分胁迫使向日葵可利用光的范围缩小,而适宜水分则扩大了光的利用范围,更有利于干物质积累。暗呼吸速率随着植物的生长进程逐渐降低,不同生育期的水分胁迫均导致暗呼吸速率降低。
向日葵;生育期;光合参数;土壤水分
引 言
光合作用是植物最基本的生命活动,水分胁迫是光合作用过程中最主要的限制因子之一,植物在水分亏缺条件下光合作用会显著降低[1-2]。研究作物对水分胁迫的响应及适应机制,是对作物进行合理调控、实现农业节水的前提,对揭示水分胁迫影响植物生长发育过程具有重要作用[3-4]。因此,近年来,土壤水分与光合作用之间关系的研究在国内日益受到重视[5-9]。食用向日葵是河套灌区经济效益十分显著的经济作物,目前向日葵种植面积占总播种面积的四分之一以上,已经成为当地农业的支柱产业[10]。深入了解其光合作用对水分和光照环境的响应,是向日葵生理生态研究的重要内容之一。近几年一些学者已经就向日葵的光合作用开展了研究:徐惠风等[11]通过FQ型CO2分析仪离体测定向日葵净光合速率;崔良基等[12-13]通过测定向日葵叶面积、光合势、干物重等群体生理指标,计算向日葵净光合速率。但针对不同生育期、不同土壤水分梯度下向日葵光合生理特性的研究鲜见报道。本试验采取人为控制水分的方法,利用LI-6400便携式光合测定系统,测定向日葵不同生育期在不同土壤水分条件下光合作用对光强变化的响应,并采用直角双曲线模型拟合光响应曲线,探讨光响应特征参数的变化规律,阐明向日葵不同生育期光合作用对土壤水分的响应规律,为揭示向日葵抗旱生理机制提供参考,也为作物模拟模型的应用提供数据基础[14-15]。
1 试验与方法
1.1 试验材料与设计
试验于2012年在内蒙古巴彦淖尔市农业气象试验站进行,供试品种为向日葵LD5009,属中熟种,平均生育期为110 d。6月2日播种,大小行种植,种植密度约为47250株/hm2,浇水、中耕锄草、施用化肥、病虫害防治等均以当地大田管理为准。9月18日成熟。土壤为壤土,0~1 m土壤容重平均值为1.36 g/cm3,田间持水量平均为26.8%。试验为小区设计,每小区面积为4 m×6 m,小区之间设置隔离层,隔离层下面为聚乙烯塑料,上面为水泥,其深度为2 m。试验场设有大型活动式防雨棚,有降水时,将棚推放到控制小区之上。灌水为井灌,灌水量通过水表记录。
试验是在人为控制水分的条件下进行的。水分设3个梯度,即土壤水分占田间持水量的40%~54.9%,55%~69.9%,70%~90%,视为缺水(T1)、基本适宜(T2)和偏湿(T3)情况。水分控制时段分3类:二对真叶—花序形成期、花序形成—开花期、开花—成熟期,其他时段水分按55%~69.9%供给,向日葵整个生育期各处理间土壤水分的控制情况见图1。由图1可以看出,各处理间3个生育时期的土壤含水量存在明显差异,达到了预期控制目的。
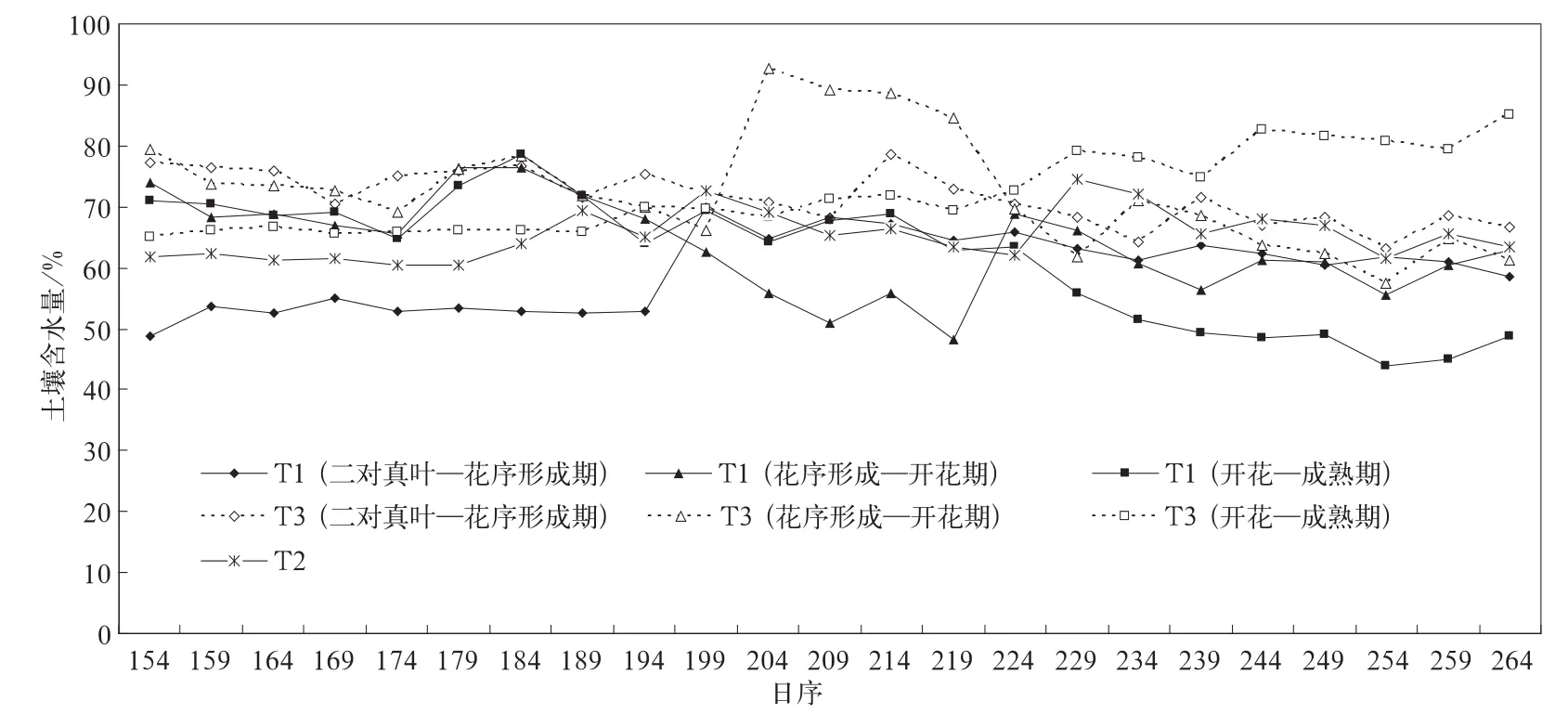
图1 向日葵全生育期土壤水分变化Fig.1 Change of the soil water content of sunflower for whole bearing period
1.2 测定项目
1.2.1 土壤水分的测定
土壤水分采用CPN-503中子土壤水分仪(美国)结合烘干法定点观测,测定深度0~200 cm,每隔10 cm为1层,每隔5 d测定1次,表层(0~20 cm)土壤水分采用土钻法。利用0~50 cm的土壤含水量确定小区是否处于控水范围。
1.2.2 光强响应测定
采用美国LI-COR公司生产的LI-6400便携式光合测定仪,对不同生长发育阶段不同水分处理的向日葵进行光合光响应测定。选择晴空09:00—11:00(北京时,下同)进行观测,光源为 LI-6400人工光源(LI-6400-02B红蓝光源),将红蓝光源LED设定12个光合有效辐射梯度:2000,1800,1500,1200,1000,700,500,200,100,50,20,0μmol·m-2·s-1,测定叶片净光合速率(Pn),每个梯度控制测定时间为120 s。测定时叶片温度为(29.3±5)℃,空气相对湿度为(23.8±4.5)%,样本室CO2浓度为(370±10)μmol·μmol-1,测定时各处理对应土壤水分存在明显差异。不同生育期不同水分梯度定株测定3株,测定部位为从上向下完全展开的第3位功能叶。
1.3 数据处理
将测定的向日葵叶片净光合速率与光合有效辐射之间的关系 (净光合速率的光响应)分别用直角双曲线与非直角双曲线方法进行拟合[16]。
利用SPSS16.0统计软件,通过迭代法分别对每一组光响应曲线进行拟合,得出相应的表观量子效率、光饱和时的表观最大净光合速率(Pmax,单位:μmol·m-2·s-1)、暗呼吸速率(Rd,单位:μmol·m-2·s-1),并且计算光补偿点和光饱和点[17]。
2 结果分析
2.1 不同生育期不同水分梯度的光响应曲线
模拟光响应曲线的数学模型较多,目前应用广泛的有直角双曲线和非直角双曲线模型。由于两种模型的表达方程不同,因此对向日葵的光响应曲线进行拟合得到的参数有所不同。用直角双曲线拟合的表观量子效率为0.0583~0.0767μmol·μmol-1,而由非直角双曲线拟合得到多为0.1244μmol·μmol-1,基本高于直角双曲线的拟合结果。理论分析表明,在实验室适宜的条件下,最大量子效率为0.08~0.125。由此可以看出,利用非直角双曲线拟合的结果接近理论上限,不符合生理意义。但由直角双曲线得出的值与在一般大田条件下的0.05~0.07范围相符。最后选择利用 Michaelis-Menten模型拟合得到的叶片光响应曲线(图2),各个生育时期不同水分处理的向日葵光合光响应曲线拟合效果较好,拟合度R2值较高(表1)。

图2 向日葵不同发育期在不同土壤水分下的光合光响应曲线(a)二对真叶—花序形成期,(b)花序形成—开花期,(c)开花—成熟期Fig.2 Theresponsecurvesofnetphotosynthetic ratetolightintensityofsunflowerunderdifferent soilmoisturesindifferentdevelopmentstages(a)twopairsofleaves-budstage,(b)budstage-floweringstage,(c)floweringstage-maturitystage
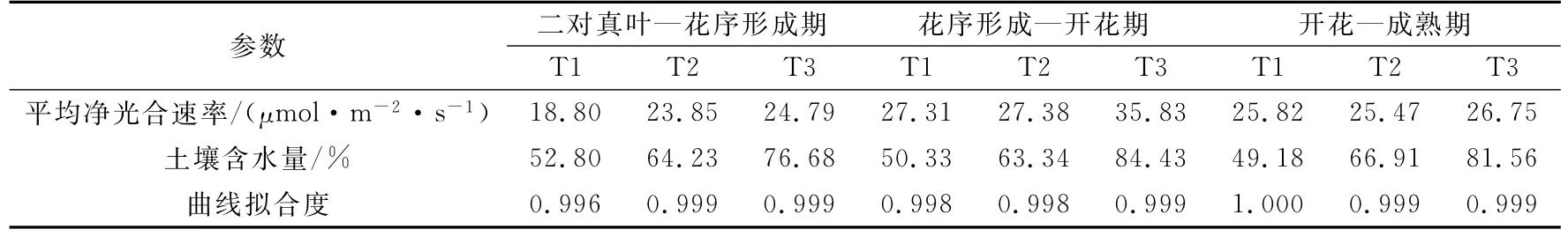
表1 向日葵不同生育期光合观测时土壤含水量、曲线拟合度和平均净光合速率的比较Table1 Thesoilwatercontent,curvefittingandnetphotosyntheticrateof sunflowerindifferentdevelopmentstages
由图2可以看到,向日葵不同生育期在不同水分处理下Pn均表现出随着光合有效辐射增加而增加,但最大光强设置为2000μmol·m-2·s-1时,光合曲线仍有上升趋势,尚未达到饱和,说明向日葵属强喜光作物,光饱和点较高。不同生育时段在不同水分梯度下光合光响应曲线均呈现相似变化规律,即在同等的光合有效辐射下Pn值表现为随水分减少而减少,尤其是随着光合有效辐射的增大愈加明显。光合光响应曲线的变幅表现出在某一生育期内水分不同对它们光能利用率的影响不同。在光合有效辐射超过500μmol·m-2·s-1后,Pn差异开始增大,因此通过500~2000μmol·m-2·s-1测定的Pn进行平均(表1)。由表1可知,不同生育期对光能利用率不同,表现出花序形成—开花期最高,开花—成熟期次之,二对真叶—花序形成期最低。不同生育期不同的水分梯度对光能利用率的影响明显不同,二对真叶—花序形成期(营养生长期)水分胁迫对该时期的光能利用率影响较大,而处理T2和处理T3差异不明显。花序形成—开花期(生殖生长期)处理T3的光能利用率远高于其他两种水分处理,说明生殖生长期对水分的需求和敏感程度高于营养生长期。开花—成熟期作物处于灌浆期,水分对作物生长的影响变得不明显,净光合速率差异不显著。
2.2 不同生育期不同水分梯度的光合参数
表观量子效率、光饱和时的最大光合作用速率、暗呼吸速率和光饱和点以及光补偿点是描述植物光合作用特征的重要生理生态特征参数[1]。
2.2.1 最大净光合速率
光饱和时的最大光合作用速率(Pmax)是叶片光合能力的一个度量。由表2可以看出,不同的生育期不同水分梯度下Pmax均表现为随着土壤水分减少而降低,但差异不同。二对真叶—花序形成期水分适宜处理T2较水分胁迫处理T1增加了55.7%,过湿处理T3较胁迫处理T1增加了68.6%。花序形成—开花期适宜处理较胁迫处理仅增加了0.12%,二者相差不大,过湿处理T3较胁迫处理T1增加了58.1%。开花—成熟期水分对各处理的影响差异不显著,Pmax在44.08~53.26μmol·m-2·s-1之间变化。水分对作物不同生育期的影响程度不同,在营养生长期试验设计的适宜处理对光合潜力的发挥影响不大,而水分胁迫严重限制了光合潜力的发挥,导致作物苗期生长缓慢;生殖生长期土壤偏湿使光合潜力极大地发挥出来,最大净光合达到最大;灌浆期植株生长缓慢水分限制作用减小,所以作物对水分差异的反应不明显。
2.2.2 表观量子效率
表观量子效率是反映植物在弱光下吸收、转换和利用光能能力的指标[18],是光合作用中光能转化效率的指标之一,该值高说明叶片光能转化效率高[19]。由表2可见,土壤湿度对最大净光合速率和表观量子效率的影响并不同步。最大净光合速率随土壤湿度的增加而增大,而表观量子效率在生长发育早期(二对真叶—花序形成期)在水分胁迫情况下出现最大值,即胁迫处理最大,适宜处理次之,过湿处理最低,且在开花期水分胁迫下表观量子效率也较高,但在开花—成熟期则相反,说明作物生长前期对水分胁迫的自身调节能力要强于后期。
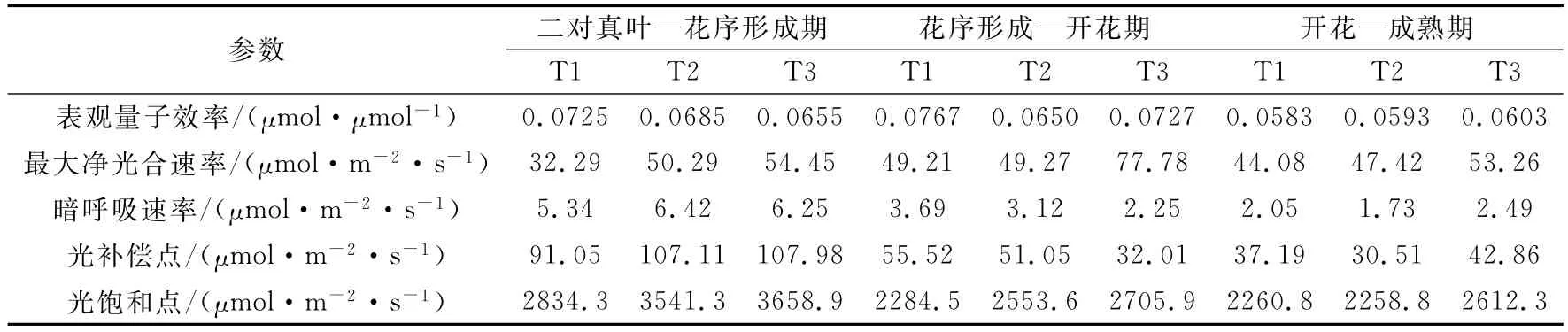
表2 向日葵不同发育期不同土壤水分条件下的光响应曲线特征参数Table 2 Characteristic parameters of sunflower light response curves under different soil moistures in different development stages
2.2.3 光饱和点和光补偿点
光补偿点是植物利用弱光能力大小的重要指标,该值越小表明利用弱光的能力越强。光饱和点是植物利用强光能力大小的指标[8,20]。由表2可以看出,营养生长期向日葵的光饱和点和光补偿点值最高,表明该时期是向日葵生长最旺盛的阶段,对光的需求量最大,耐阴性最差。进入生殖生长期光饱和点和光补偿点值逐渐趋于稳定,成熟期差异更小。二对真叶—花序形成期胁迫处理T1利用弱光的能力最强,其他两种处理对弱光的利用能力相当。该时期对强光的利用随着土壤水分的增加而增加,但当土壤水分增加到偏湿时光饱和点增加并不显著,可见过多的水分在该时期并不会引起干物质的显著增加。花序形成—开花期不同土壤水分含量下光补偿点差异显著,表现出随着土壤水分的增加逐渐减少,而光饱和点却相反,表明水分胁迫在该时期使向日葵对光能的利用范围发生了变化,越干旱越向弱光区偏移,光能利用范围越狭窄,而偏湿的土壤水分使向日葵利用光能的范围变宽,更有利于干物重的积累,这与韩刚等[7]对4种沙生灌木的光合特性研究结果一致。同时水分胁迫使补偿点增高,光饱和点下降,李林芝等[6]对羊草的研究也得到了类似结论,表明水分胁迫下使植物叶片气孔关闭,叶肉细胞的光合活性下降,光能利用率降低。开花—成熟期随着土壤水分的增加,延缓了叶片的衰老,过湿处理较其他水分处理的光饱和点和光补偿点均较高,但水分胁迫对光合的影响越来越小。
2.2.4 暗呼吸速率
暗呼吸速率(Rd)反映的是植物在没有光照条件下的呼吸速率[21],该特征值主要和叶片的生理活性有关[7]。向日葵表现出在营养生长阶段叶片的Rd达到最大,不同处理的Rd平均值为6,继而随着植物的生长Rd逐渐降低,在生殖生长阶段平均值为3.019μmol·m-2·s-1,成熟期降为最低,平均值为2.09μmol·m-2·s-1,表明叶片的生理活性在营养阶段最为旺盛。二对真叶—花序形成期Rd在水分适宜条件下最高,胁迫处理较适宜处理降低了16.8%,说明在营养生长期水分胁迫使向日葵叶片生理活性降低,在一定程度上减少呼吸对光合产物的消耗,增加干物质积累。花序形成—开花期Rd表现出随着土壤水分增加而减少,可能是由于该时期是作物的需水关键期,而水分胁迫激发了叶片的活性,相反,过湿处理最大净光合速率最高而Rd最低,更有利于干物质的积累。开花—成熟期叶片逐渐衰老,活性降低,暗呼吸消耗也呈现出降低态势。
植物在水分胁迫下能够维持其Pmax和表观量子效率在较高水平,表明其能有效利用弱光及强光进行光合作用,对光的生态适应性强[20],并在一定程度上反映了植物光合作用对干旱的适应性较强[22]。本文向日葵表观量子效率为0.0583~0.0767μmol·μmol-1,Pmax为33.29~77.78μmol·m-2·s-1,说明即使受到水分胁迫向日葵仍具有高光能利用率。不同生育期土壤湿度不但影响Pmax,而且影响表观量子效率,并且对二者的影响并不同步。每个发育期Pmax均随着土壤湿度增加而增大,而表观量子效率(生长前期)却在一定程度的水分胁迫下出现最大值,表明干旱情况下,作物有一定的自身调节机制,提高初始光合效率,尽可能使光合作用达到较高,部分弥补水分胁迫带来的副作用,且这种调节能力随着植物的生长逐渐降低,这与植物生理学研究结果[23]基本一致。
在不考虑水分影响情况下,向日葵整个生育期光饱和点和光补偿点分别为30.51~107.98μmol·m-2·s-1和2260.8~3658.9μmol·m-2·s-1,与文献[24]中阳性植物的光补偿点(50~100μmol·m-2·s-1)相一致,较光饱和点(1500~2000μmol·m-2·s-1)更高,说明向日葵是耐阴性较差特别喜光的典型阳生植物。崔良基等[12-13]从群体生理指标的角度进行了研究,也得出向日葵叶片光合能力在各种作物中十分突出。徐惠风等[11]通过FQ型CO2分析仪离体测定向日葵净光合速率,得出向日葵光饱和点为1700μmol·m-2·s-1,光补偿点为95~120μmol·m-2·s-1。其中,光饱和点远低于本试验结果,光补偿点略高。与本试验研究结果不同,主要是试验方法不同所致,同时也受到测定发育期和周围环境因子等因素的影响。
3 结 论
本文利用向日葵观测数据及其拟合光响应曲线模型提取的光合特征参数,分析了不同水分胁迫下向日葵各生育时期的光响应特征,主要结论如下:
1)在试验设置光强下,各生育期对光强均表现出净光合速率随着光合有效辐射增加而增加;同等光合有效辐射下,净光合速率也随着土壤水分增加而增加,尤其是随着光合有效辐射的增大愈加明显。
2)土壤湿度对最大净光合速率和表观量子效率的影响并不同步,最大净光合速率随着土壤湿度的增加而增大,而表观量子效率在一定程度的水分胁迫情况下出现最大值。
3)不同的土壤水分含量对光补偿点和光饱和点影响不同,光饱和点随着土壤水分的增加而增加,光补偿点却相反,表明水分胁迫使向日葵可利用光的范围缩小,而适宜水分则扩大了光的利用范围,更有利于干物质积累。
4)暗呼吸速率随着植物的生长进程逐渐降低,不同生育期的水分胁迫均导致暗呼吸速率降低。
[1] 于贵瑞,王秋凤.植物光合、蒸腾与水分利用的生理生态学.北京:科学出版社,2010.
[2] 欧立军,陈波,邹学校.干旱对辣椒光合作用及相关生理特征的影响.生态学报,2012,32(8):2612-2619.
[3] 田育红,刘鸿雁,陆佩玲.山东禹城地区夏玉米生长与环境因子关系的初步分析.应用气象学报,2003,14(4):509-512.
[4] 刘建栋,王馥棠,于强,等.华北地区冬小麦叶片光合作用模型在农业干旱预测中的应用研究.应用气象学报,2003,14(4):469-478.
[5] 马全林,王继和,纪永福,等.固沙树种梭梭在不同水分梯度下的光合生理特征.西北植物学报,2003,23(12):2120-2126.
[6] 李林芝,张德罡,辛晓平,等.呼伦贝尔草甸草原不同土壤水分梯度下羊草的光合特性.生态学报,2009,29(10):5271-5279.
[7] 韩刚,赵忠.不同土壤水分下4种沙生灌木的光合光响应特性.生态学报,2010,30(15):4019-4026.
[8] 刘建栋,周秀骥,于强.中国黄淮海地区冬小麦光合作用特征参数.应用气象学报,2003,14(3):257-265.
[9] 吴玮,景元书,马玉平,等.干旱环境下夏玉米各生育时期光响应特征.应用气象学报,2013,24(6):723-730.
[10] 杨松,刘俊林,卢淑贤,等.河套灌区向日葵适宜播种农业气象指标研究.中国农学通报,2009,25(11):176-179.
[11] 徐惠风,徐克章,刘兴土.向日葵光合特性及其对不同生态条件的响应.农村生态环境,2004,20(1):20-23.
[12] 崔良基,梁国战,王德兴.夏播条件下向日葵光合性能与杂交种产量的关系.中国油料作物学报,2002,24(1):48-50.
[13] 崔良基,王德兴,宋殿秀,等.不同向日葵品种群体光合生理参数及产量比较.中国油料作物学报,2011,33(2):147-151.
[14] 马玉平,王石立,王馥棠.作物模拟模型在农业气象业务应用中的研究初探.应用气象学报,2005,16(3):293-303.
[15] 孙琳丽,马玉平,景元书,等.基于约束性分析的数据与作物模型同化方法.应用气象学报,2013,24(3):287-296.
[16] 陆佩玲,罗毅,刘建栋,等.华北地区冬小麦光合作用的光响应曲线的特征参数.应用气象学报,2000,11(2):236-241.
[17] 王建林,于贵瑞,房全孝,等.不同植物叶片水分利用效率对光和CO2的响应与模拟.生态学报,2008,28(2):525-533.
[18] 李合生.现代植物生理学.北京:高等教育出版社,2002.
[19] 许大全.光合作用效率.上海:上海科学技术出版社,2002.
[20] 伍维模,李志军,罗青红,等.土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特征的影响.林业科学,2007,43(5):30-35.
[21] Coley P D.Herbivory and defensive characteristics of tree species in a low land tropical forest.Ecological Monographs,1983,53(2):209-233.
[22] 刘世荣,赵广东,马全林.沙木廖和沙枣对地下水位变化的生理生态响应 Ⅱ.叶片光合作用及其对温度和光的反应.植物生态学报,2003,27(2):223-227.
[23] 邹琦.作物抗旱生理生态研究.济南:山东科学技术出版社,1994.
[24] 蒋高明.植物生理生态学.北京:高等教育出版社,2004.
Effects of Different Soil Moistures on Photosynthetic Characteristics of Sunflower
Yun Wenli1)Hou Qiong1)Wang Haimei1)Li Jianjun2)Zhang Chao3)
1)(Meteorogical Research Institute of Inner Mongolia,Inner Mongolia Meteorological Bureau,Hohhot010051)
2)(Linhe Astronomical Observatory,Bayannaoer Meteorological Bureau of Inner Mongolia,Linhe015000)
3)(Inner Mongolia Ecological and Agricultural Meteorological Center,Hohhot010051)
The soil drought is one of key factors limiting photosynthesis in northwest areas of China.In order to understand the influence of drought stress on crop,the light response curves and several parameters of photosynthesis of sunflower are measured with Li-6400 Portable Photosynthesis System under three soil moisture grades:Arid soil,suitable soil moisture and wetter soil moisture(corresponding soil water contents are 40%-54.9%,55%-69.9%and 70%-90%,respectively)in different development stages(two pairs of leaves-bud stage,bud stage-flowering stage,flowering stage-maturity stage).Results show that all coefficients of light response curve equations fitted using rectangular hyperbola model are above 0.99,meaning good compatibility between the model and the response process of photosynthesis of sunflower to light.The light response curves of sunflower growing under different soil moisture in different development stages show similar trends,the net photosynthetic rate(Pn)increases with photosynthetic active radiation rapidly at first and then slowly.For different development stages,changes of light response curves show a similar regulation with reduced soil moisture,butPnof sunflower under wetter soil moisture is greater than that under suitable soil moisture and drought stress at the same photosynthetic ac-tive radiation,and differences are statistically significant with the photosynthetic active radiation increasing.Influences of soil moisture on the maximum net photosynthetic rate(Pmax)and apparent quantum efficiency are not synchronous.Pmaxincreases with soil moisture and apparent quantum efficiency under the condition of water stress in maximum.In the entire growth period of sunflower in the Hetao Irrigation District,light compensation point and light saturation point are 30.51-107.98μmol·m-2·s-1and 2260.8-3658.9μmol·m-2·s-1,respectively.It shows that sunflower with high solar energy utilization efficiency is the typical sun plants,and is particularly fond of light.The effect of soil moisture content in light compensation point and light saturation point is different.The light saturation point increases with soil moisture,while light compensation point is the opposite.According to the variation of light compensation point and light saturation point,sunflower under suitable soil moisture not only expands the scope of the use of light but also is conducive to the accumulation of dry matter,sunflower under drought stress narrows the range of available light.The dark respiration rate(Rd)decreases gradually with plant growth,and decreases under drought stress in different development stages,which is conducive to reduce the influence of drought stress on dry matter accumulation of crops.
sunflower;development stages;photosynthetic parameters;soil moisture
云文丽,侯琼,王海梅,等.不同土壤水分对向日葵光合光响应的影响.应用气象学报,2014,25(4):476-482.
2013-10-16收到,2014-05-08收到再改稿。
公益性行业(气象)科研专项(GYHY201206021)
*通信作者,email:1983572958@qq.com
