孵化温度影响中华鳖孵出幼体表型特征和幼体的早期生长
2014-07-05吴义莲许雪峰
吴义莲,许雪峰
中华鳖(Pelodiscus sinensis)是一种经济鳖类,中华鳖在我国广泛分布,除新疆、西藏和青海外,其它各省均有分布。中华鳖在我国用于食物、宠物和保健品的开发具有较为悠久的历史[1],每年的消耗量以万吨计。野外中华鳖在毫无节制的开采下已不多见,这就促成了过去几十年中华鳖人工养殖业的蓬勃发展[2]。
对于卵生爬行动物而言,卵孵化成功率的高低是中华鳖养殖企业能否快速扩繁、提高效益的关键因素。因此,怎样提高孵化成功率是中华鳖养殖业重点关注的问题之一[3-5]。爬行动物已有的研究表明,在极端的温度下,爬行动物的胚胎不能发育完全[6]。低温延缓或滞留胚胎发育,但通常对胚胎发育产生较小或没有致死效应,然而极端高温可能导致胚胎畸形率和死亡率大幅度增长[7-8]。在后代可存活的温度范围内,较高温度可导致胚胎发育较快,因此,缩短了胚胎发育周期。中华鳖的早期研究表明孵化温度影响孵化成功率和孵出幼体表型[9-13]。在24 - 34℃范围内,孵化期随着温度的升高而减少,但是其孵化成功率却降低,甚至导致孵出畸形幼体[9-13]。但是短期将卵至于极端温度却不影响孵化成功率和幼体的表型。
中华鳖养殖业重点关注的另一个问题是孵出幼体质量如何影响其后续的生长。尽管孵化温度可使得胚胎发育的速度达到最大,但通常不会使后代表型达到最佳。很多的研究表明,爬行动物胚胎所经历的热环境对幼体表型特征有精细地影响,包括个体大小、形态、能量储存、行为和胚后生长[12]。
在本研究中,我们设置了5个温度孵化中华鳖卵,检测温度对孵出幼体表型特征和后续生长的影响,研究结果对中华鳖养殖企业优化孵化程式以获得优质种苗和扩繁有着重要意义。
1 材料与方法
1.1 雌体繁殖和卵孵化
实验用中华鳖卵于2012年5月中旬至6月初取自全椒养殖场。本研究共收集了41窝501枚中华鳖卵,平均窝卵重和平均窝卵数分别为36.3±1.4g和 12.2±0.5枚。
实验设置了21、24、28和32℃四个恒定温度和一个自然波动温度孵化中华鳖卵。所有卵在入孵前根据卵动植物极的形态初步检测其是否受精,未受精卵不用于卵孵化,在总共501枚卵中共有140枚为未受精卵。孵化期以产卵结束开始至幼体出壳为止。根据研究的经验,32℃部分幼体在孵化期末时采用人为辅助方式破壳。孵化成功率以孵出的幼体数除以总卵数来计算。
1.2 幼体饲养和幼体形态特征测量
幼体孵出后,在2小时内用Mettler天平测量幼体的湿重(精确到g),孵出幼体的背甲长、背甲宽和腹甲宽用Mitutoyo数显游标卡尺测量,剪趾标记,以备后续识别,然后释放到全椒养殖场饲养幼体。在第一个越冬季前(11月中旬)测量饲养幼体的体重、背甲长、背甲宽和腹甲宽,以检测孵化温度对生活史特征的影响。
1.3 数据分析
所有数据在作参数统计分析之前,分别检验其正态性(Kolmogorov-Smirnov test)和方差同质性(F-max test)。经检验,部分的原始数据须要经过ln转化以后才能用于参数统计分析。用卡方检验、单因子方差分析(one-way ANVOA)、单因子多变量方差分析(one-way MANVOA)和Tukey’s 检验比较等统计方法来处理和比较相应的数据。描述性统计值用平均值±标准误表示,显著性水平设置为 α= 0.05。
2 结果
2.1 孵化温度对孵出幼体形态特征的影响
孵化温度影响中华鳖的孵化期(Kruskal-Wallis ANOVA, H(4,N=277)= 245.90,P< 0.001),孵化期随着温度的升高呈非线性变短(图1)。21、24、28、32(℃)和波动温度的孵化成功率分别为 63.2% (60/95)、58.8% (40/68)、67.6% (50/74)、62.9% (61/97)和 83.5% (66/79),五个温度的孵化成功率差异不显著(卡方检验, χ2= 5.52,df= 4,P> 0.23)。五个温度下入孵卵,卵重没有差异(one-way ANOVA,F4,408= 0.93,P= 0.45),然而,孵化温度影响孵出幼体的湿重、幼体的背甲宽和腹甲宽(one-way ANOVA, allP< 0.001),但不影响孵出幼体的背甲长(F4,271= 1.32,P=0.26)。波动温度孵化的幼体较其它温度在湿重、背甲宽和腹甲宽上要小(表1)。
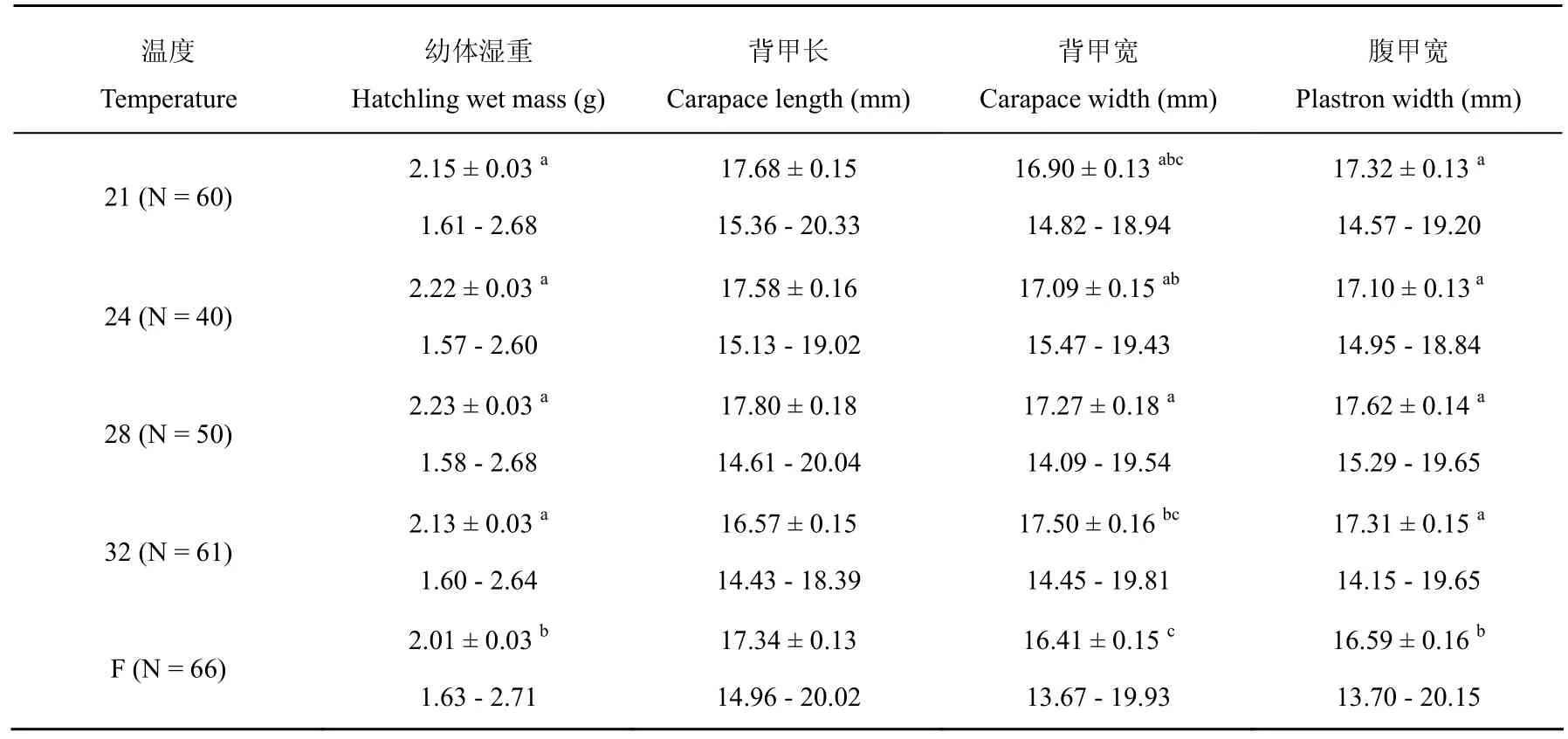
表1 中华鳖孵出幼体形态特征描述性统计值
2.2 孵化温度对孵出幼体早期生长的影响
来源于21、24、28、32(℃)和波动温度孵化的幼体在第一个越冬季前的存活率分别为76.7%、75.0%、78.0%、73.8%和62.1%,存活率差异不显著(卡方检验, χ2= 2.22,df= 4,P>0.69)。然而,体重、背甲长、背甲宽和腹甲宽在第一个越冬季前差异显著(one-way ANOVA, allP< 0.01),较低温度孵出(21和24℃)的幼体比较高温度(28、32℃和波动温度)的幼体体重轻,背甲长、背甲宽和腹甲宽小(Tukey’s test, allP<0.0001)。对体重、背甲长、背甲宽和腹甲宽进行多变量方差分析表明,不同温度来源的幼体形态特征差异显著(one-way MANOVA, Wilks λ = 0.34,F4,16= 15.78,P< 0.01)(表 2)。
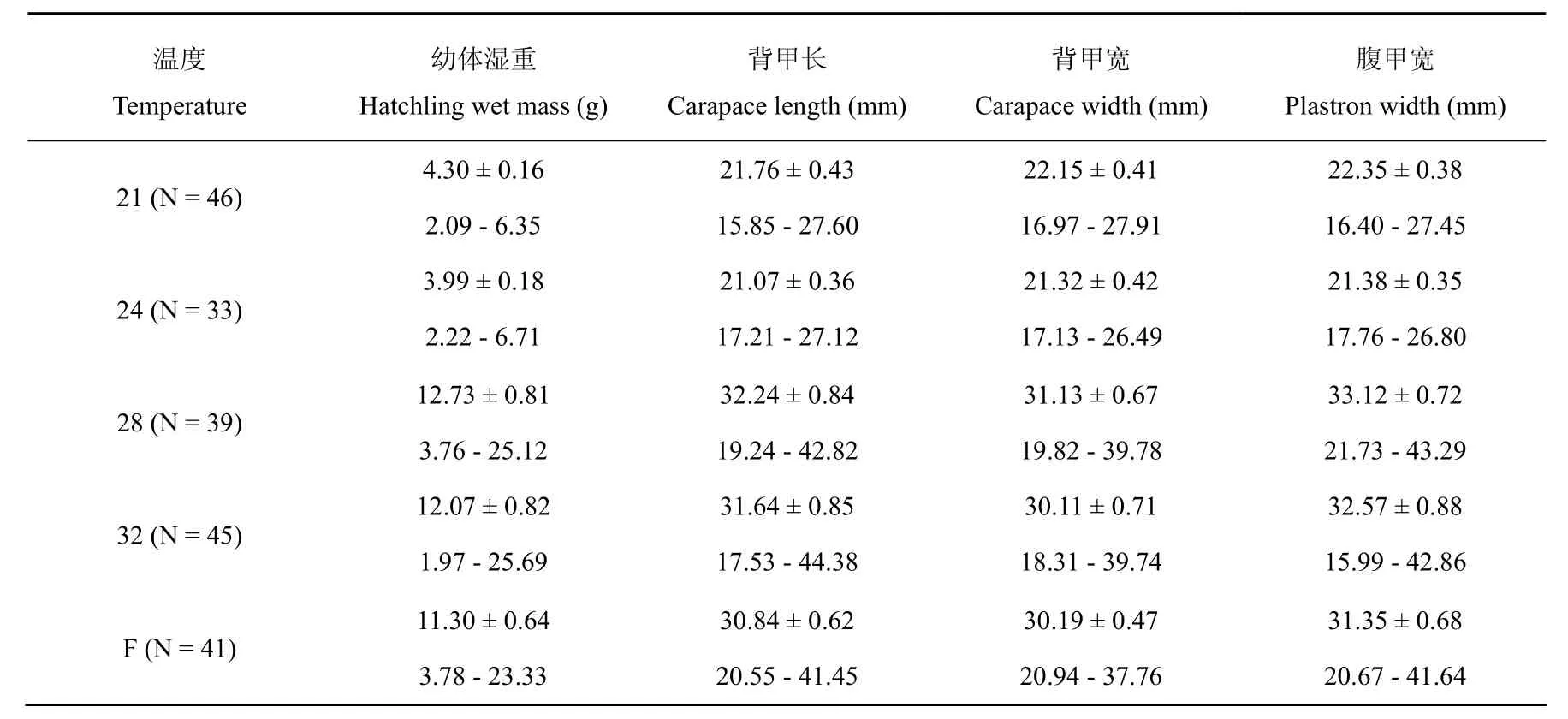
表2 中华鳖幼体第一个越冬季前形态特征描述性统计值
3 讨论
已有的研究表明孵化温度对胚胎发育具有不同的作用效力,高温能加快爬行动物胚胎发育的速率,但却会导致胚胎致畸或死亡[13-15]。与其他很多对爬行动物研究结果的报道一样,在卵孵化过程中,中华鳖的胚胎所经历的孵化热环境影响孵化成功率、孵化期和幼体的大小。孵化期随着孵化温度的升高而变短,32℃胚胎发育最快,而21℃最慢(图1)。与其他研究不同的是,温度不影响孵化成功率。然而,在实验过程中,高温(32℃)和低温(21℃)孵化的中华鳖幼体通常不能自行破壳。本研究中,由于我们在接近特定温度孵化期限内采用了人为辅助的方式帮助中华鳖幼体破壳,造成了不同温度孵化成功率差异不显著。因此,研究结果并不与其他已有的研究相悖[9-13]。温度影响孵出幼体的表型特征,本研究中,波动温度孵出的幼体大小较其它恒定温度小,表明在中华鳖卵孵化中,较大温度范围的波动对幼体大小没有有益的影响(表1),我们的结果与其他已有的研究一致[13]。
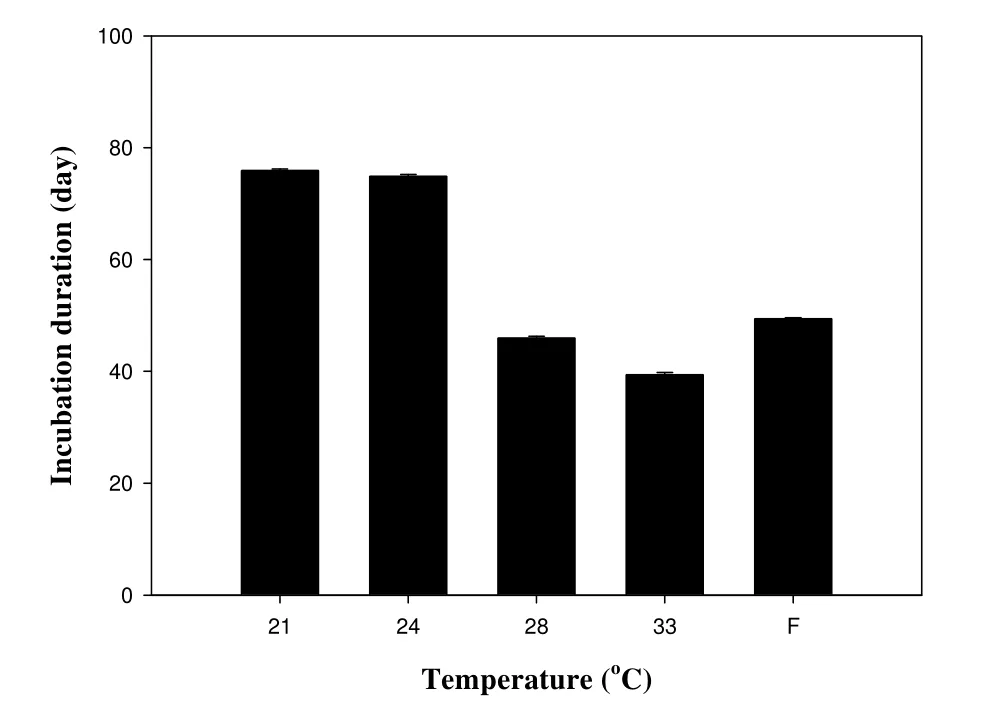
图1 孵化温度对孵化期的影响数据以平均值±标准误表示
胚胎较早孵出意味着距越冬期更长,适宜于幼体早期生长的时间也越长,因此,孵化期可能是一个与个体生长适合度有关的特征。本研究的结果发现,在第一个越冬季前,来源于较高温度的幼体比较低温度的幼体要大(表2)。究其原因,较高温度早孵出的幼体至第一个越冬季前有更长的时间(约一个月)摄入更多的能量用于生长,所以个体相对较大。大多数动物的个体大小与适合度相关[16-18],然而,实验室和养殖业在食物资源丰富、热环境及其它生物和非生物适合的情况下,孵化环境对孵出幼体大小的效应在长时间幼体后期的生长上常不容易检测得到,中华鳖的早期研究已表明孵化温度诱导的幼体大小的变异对幼体的后期生长没有显著的影响[12]。本研究发现波动温度孵化出的后期幼体的存活率在数值上较其它温度低,这一结果也佐证了波动温度孵化中华鳖卵并不比恒定温度有优势[15]。
[1]Chen TH, Chang HC, Lue KY. Unregulated trade in turtle shells for Chinese traditional medicine in East and Southeast Asia: the case of Taiwan [J]. Chelonian Conservation and Biology, 2009, 8 (1): 11–18.
[2]Shi HT, Parham JF, Fan ZY, Hong ML, Yin F. Evidence for the massive scale of turtle farming in China [J].Oryx, 2008, 42 (1): 147–150.
[3]Du WG, Hu LJ, Lu JL, Zhu LJ. Effects of incubation temperature on embryonic development rate, sex ratio and post-hatching growth in the Chinese three-keeled pond turtle, Chinemys reevesii [J]. Aquaculture, 2007,272 (1-4): 747–753
[4]Du WG, Wang L, Shen JW. Optimal temperatures for egg incubation in two Geoemydid turtles: Ocadia sinensis and Mauremys mutica [J]. Aquaculture, 2010,305 (1-4): 138–142.
[5]Zhu XP, Wei CQ, Zhao WH, Du HJ, Chen YL, Gui JF.Effects of incubation temperatures on embryonic development in the Asian yellow pond turtle [J].Aquaculture, 2006, 259 (1-4): 243–248.
[6]Vinegar A. Evolutionary implications of temperature induced anomalies of development in snake embryos [J].Herpetologica, 1974, 30 (1): 72–74.
[7]Andrews RM, Qualls CP, Rose BR. Effects of low temperature on embryonic development of Sceloporus lizards [J]. Copeia, 1997, 1997 (4): 827–833.
[8]杜卫国,计翔. 孵化温度对灰鼠蛇卵孵化期、孵化成功率和孵出幼体特征的影响. [J]. 生态学报, 2002, 22(4): 548–553.
[9]Choo BL, Chou LM. Effect of temperature on the incubation period and hatchability of Trionyx sinensis Wiegmann eggs [J]. Journal of Herpetology, 1987, 21(3): 230–232.
[10]杜卫国,计翔. 孵化温度对中华鳖胚胎物质和能量利用的影响. [J]. 生态学报, 2001, 47 (5): 512–517.
[11]Du WG, Ji X. The effects of incubation thermal environments on size, locomotor performance and early growth of hatchling soft-shelled turtles, Pelodiscus sinensis [J]. Journal of Thermal Biology, 2003, 28 (4):279–286.
[12]Ji X, Chen F, Du WG, Chen HL. Incubation temperature affects hatchling growth but not sexual phenotype in the Chinese soft-shelled turtle Pelodiscus sinensis[J]. Journal of Zoology, 2003, 261 (4): 409–416.
[13]Li H, Zhou ZS, Wu T, et al. Do fluctuations in incubation temperature affect hatchling quality in the Chinese soft-shelled turtle Pelodiscus sinensis [J]?Aquaculture, 2013, 406-407: 91-96
[14]Bell K, Blomberg S, Schwarzkopf L. Detrimental influence on performance of high temperature incubation in a tropical reptile: is cooler better in the tropics [J]. Oecologia, 2013, 171 (1): 83–91.
[15]Booth DT. Influence of incubation temperature on hatchling phenotype in reptiles [J]. Physiological and Biochemical Zoology, 2006, 79(2): 274–281.
[16]Janzen FJ, Tucker JK, Paukstis GL. Experimental analysis of an early life-history stage: avian predation selects for larger body size of hatchling turtles [J].Journal of Evolutionary Biology, 2000, 13 (6): 947–954.
[17]Janzen FJ, Tucker JK, Paukstis GL. Experimental analysis of an early life-history stage: selection on size of hatchling turtles [J]. Ecology, 2000, 81 (8):2275–2280.
[18]Ji X, Du WG, Qu YF, Lin LH. Nonlinear continuum of egg size-number tradeoffs in a snake: is egg-size variation fitness related [J]? Oecologia, 2009, 159 (4):689–696.
