稻瘟病菌中侵染初期特异表达的转录因子筛选
2014-06-27孟秀利鲁国东福建农林大学植物保护学院福建福州350002浙江大学生物与技术研究所浙江杭州30029
孟秀利, 李 亚, 周 波, 鲁国东*(.福建农林大学植物保护学院, 福建 福州 350002; 2.浙江大学生物与技术研究所, 浙江 杭州 30029)
稻瘟病是危害水稻的重要病害,此病在我国南北稻区每年都不同程度的发生和流行,产量损失达数亿千克,给我国的粮食安全带来隐患(邹声浩,2009)。鉴于此,弄清楚稻瘟病菌的侵染模式以便更好的控制稻瘟病的流行就变得尤为重要。转录因子是反式作用因子中最大的家族,对真菌生长发育和致病性有重要影响。稻瘟病菌侵染水稻时相关基因的表达离不开转录因子的参与。关于稻瘟病菌转录因子在侵染时期的作用很多人都有研究。稻瘟病菌转录因子MoMSN2敲除后,突变体生长速率明显下降,菌丝生长致密,并且突变体几乎不产生分生孢子,不能形成附着胞,且丧失了对水稻的致病性。MoMYB1敲除后也影响了稻瘟病菌的生长速率,分生孢子梗的发育等(赵倩,2010)。张莉林对稻瘟病菌294个转录因子进行敲除,发现49个转录因子影响了稻瘟菌的生长、产孢和致病性(张莉林,2013)。Sook-Young Park等人研究了206个稻瘟病菌的转录因子,发现这206个转录因子50%的与产孢相关,敲除10个产孢时期特异表达的转录因子,分生孢子不能形成(Sook-Young Parketal.,2012)。
本实验在稻瘟病菌数据库中找到稻瘟病菌假定的的转录因子有500个。通过对500个假定的转录因子的序列进行分析,发现有些序列在稻瘟病菌基因组序列中高度重复,去除这些重复序列后对剩余的486个转录因子进行RT-PCR筛选, 目的是找到在侵染初期特异表达的转录因子。这些数据的获得不仅可以让我们更清楚稻瘟病菌转录因子的作用,更重要的是对于研究无毒基因的调控模式也大有意义。因为有文献指出,稻瘟病菌的42个预测分泌蛋白中有12个基因在侵染水稻早期显著表达(李湘龙,2012),同时Isabelle Fudal等证明无毒基因ACE1在侵染钉的侵入时期起重要作用(Isabelle Fudaletal., 2007),所以找到在侵染时期特异表达的转录因子对于研究稻瘟病菌无毒基因的表达以及克隆也有着重要的意义。
1 材料与方法
1.1 试验材料
菌株R88-002由韩国 Kyung Hee 大学的 Jong-Seong Jeon 教授提供, RNA提取试剂盒购于上海普洛生物产品有限公司, 反转录试剂盒购于宝生物工程有限公司, rTaq酶购于泰京生物技术公司。
1.2 试验方法
1.2.1 RNA提取 R88-002菌株活化后并转接在CM培养基上(20×Nitrate salts 50 ml,1000×Trace element 1 ml,D-glucose 10 g, peptone 2 g, Yeast Extract 1 g, casamino acid 1 g,1000×Vitamin solution 1 ml, Agar 15 g 定容到1000 ml))待菌丝长满全皿后,用RNA提取试剂盒按说明中方法,提取菌丝RNA。侵染初期材料是采取洋葱表皮侵染的方法获得。方法是将新买的洋葱侵泡一夜后撕取洋葱表皮,将稻瘟病菌孢子滴到洋葱疏水表面,放在放有吸水纸的盘中用保鲜膜密封保湿,24 h后用试剂盒提取RNA。
1.2.2 反转录获得cDNA 首先用gDNA Eeaser 去除RNA中的DNA,然后利用反转录酶将RNA 反转录成cDNA。按说明书方法进行。
1.2.3 486个转录因子引物的设计 首先在数据库中将这些基因的基因序列找出来,在这些基因序列的CDS区内部设计正向引物和反向引物,两个引物之间跨越一个内含子,扩增片段大小控制在500 bp以内,两个引物的3,末端以C或G结束,引物的长度在18-20 bp之间,正向和反向引物的GC含量尽量接近,且GC含量在引物序列中的含量在45%-65%之间。用这些原则进行引物设计,然后用DNAMAN软件进行引物的检验,对于有严重的发卡结构的引物进行重新设计。引物设计后,由生工生物工程股份有限公司合成。
2 结果
2.1 菌丝阶段RT-PCR扩增结果
486个转录因子经过RT-PCR扩增,共有160个转录因子在菌丝时期有表达,其余326个转录因子未检测到表达。有表达的转录因子根据其表达量相对于Tublin基因表达量的高低,分为低、中、高三类,其中比Tublin基因表达量低的有120个,与Tublin表达量相同的有 32个,比Tublin基因表达量高的有8个。这些在菌丝阶段有表达的转录因子对应的基因号见表1。

表1 用菌丝的RNA进行RT-PCR筛选结果Table 1 The screening results using hypha RNA by RT-PCR
2.2 侵染初期特异表达的转录因子的PCR验证
为了加快侵染初期特异表达的转录因子的筛选,我们选择在菌丝阶段不表达的326个转录因子用稻瘟病菌侵染初期的材料进行再次筛选。经过RT-PCR,发现326个转录因子中有30个转录因子在侵染初期特异表达,这些转录因子对应的基因编号是MGG_00080 、MGG_10276 、MGG_10280、MGG_15787、MGG_01887、MGG_01779、MGG_12776、MGG_04213、MGG_10694、MGG_13927 、MGG_10197、MGG_09276、MGG_09312、MGG_03758、MGG_01325、MGG_11457、MGG_17702 、MGG_07674 、MGG_05049、MGG_06453 、MGG_06566、MGG_06269、MGG_06258、MGG_09841、MGG_04401 、MGG_02045、MGG_00780 、MGG_01042 、MGG_09628、MGG_00810。这30个转录因子按照结构域的不同可以分为10种类型,分别是C2H2 Zinc finger类转录因子、Zn2Cys6类转录因子、HMG类转录因子、Winged helix repressor DNA-binding 类转录因子、Transcription factor jumonji 类转录因子、ssDNA-binding transcriptional regulator 转录因子、Zinc finger MIZ-type 类转录因子、Homeodomain-like 类转录因子、Zinc finger DHHC-type和Nucleic acid-binding, OB-fold 类转录因子。具体分类见表2。
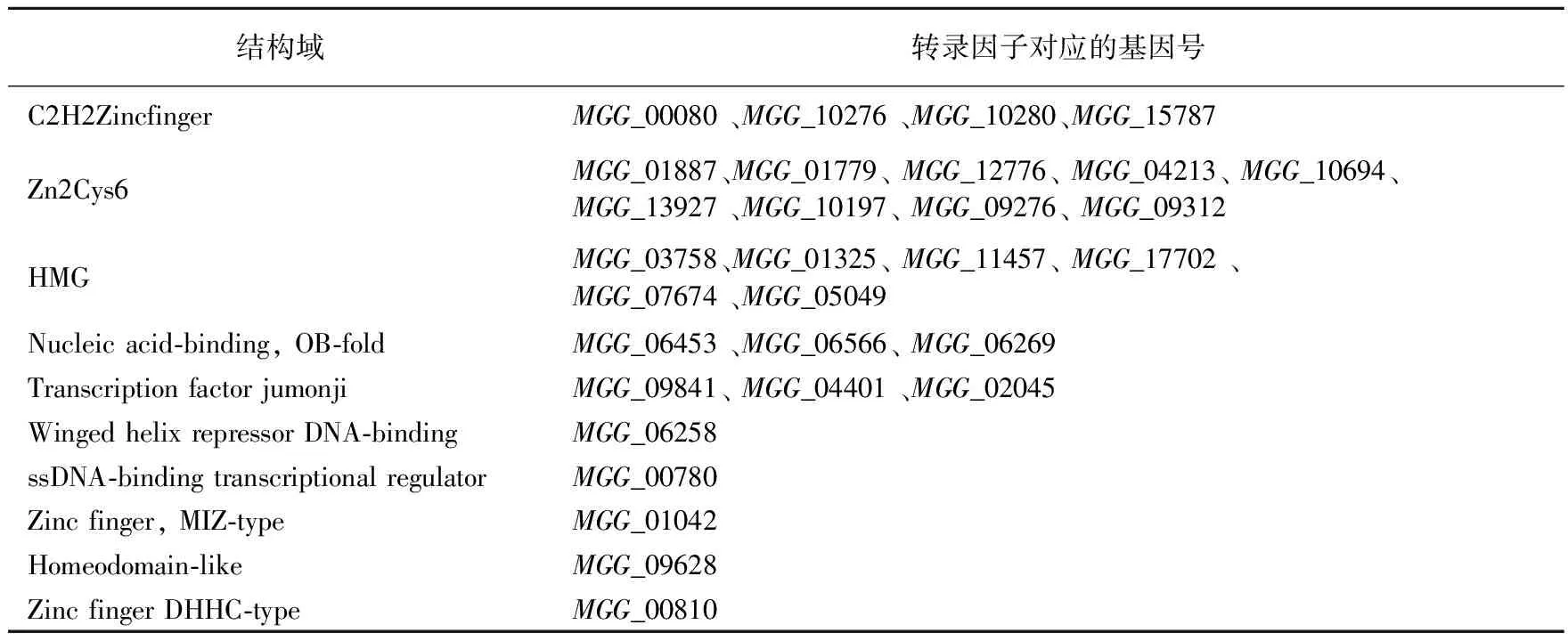
表2 侵染初期特异表达的转录因子及其类型Table 2 Classification for the 30 TFs based on their protein domain contained
3 结果与讨论
本实验通过RT-PCR验证,在稻瘟病菌486个假定的转录因子中,有160个在稻瘟病菌的菌丝生长阶段表达。 从剩余的在菌丝阶段不表达的转录因子中发现了30个在稻瘟病菌侵染洋葱表皮24 h时特异表达的转录因子。这30个转录因子按照结构域的不同可以分为10大类,其中4个含有C2H2 Zinc finger 结构域,9个含有Zn2Cys6 结构域,6个含有HMG 结构域,3个含有Transcription factor jumonji 结构域,还有3个含有Nucleic acid-binding, OB-fold 结构域,含有Winged helix repressor DNA-binding、ssDNA-binding transcriptional regulator、 Homeodomain-like 结构域、zinc finger MIZ-type结构域和 zinc finger DHHC-type结构域的分别含有1个。
本实验只是对这些转录因子进行了初步的筛选,并未进行更细致的研究。这些基因在稻瘟病菌侵染初期特异性表达,它们很可能参与了稻瘟病菌的致病过程。敲除这些转录因子是否会影响稻瘟病菌的致病特性是我们接下来想回答的问题。另外,之前的研究表明,大部分的病原真菌的无毒基因都在其侵染时期特异表达,这30个侵染时期特异表达的转录因子与无毒基因的关系是我们今后的研究重点。
李湘龙,柏斌,吴俊,邓启云,周波.2012.第二代测序技术对于水稻和稻瘟菌互作早期转录组的分析.遗传,34(1):102-112.
张莉林.2013.294个稻瘟病菌转录因子基因的敲除和功能分析.浙江:浙江大学.
赵倩.2010.稻瘟病菌转录因子MoMSN2和MoMYB1 的生物学功能研究.南京:南京农业大学.
邹声浩,黄珊,郑燕梅,方珊如,施碧红.2009.安徽农业通报,15(24):23-25.
Fugal I ,Fudal I,Collemare J,Böhnert HU,Melayah D,Lebrun MH.2007.Expression ofMagnaporthegriseaavirulence gene ACE1 Is connected to the initiation of appressorium-mediated penetration.EUKARYOTIC CELL,6(3):546-554.
Park SY,Choi J,Lim SE,Lee GW,Park J,Kim Y,Kong S,Kim SR,Rho HS,Jeon J,Chi MH,Kim S,Khang CH,Kang S,Lee YH.2013.Global expression profiling of transcription factor genes provides new insights into pathogenicity and stress responses in the rice blast fungus.PTHOGENS,9(6):e1003350.
