健康成人和家兔脑电图的比较研究:梦象荒诞离奇特性的实验分析与验证
2014-05-21史均翰渭南师院梦象研究中心渭南714000通讯作者mail09132135116sohucom
史均翰,王 军(渭南师院梦象研究中心,渭南 714000;通讯作者,E-mail:09132135116@sohu.com)
古今中外,梦象一直是吸引人们关注、诱惑人们探索的对象。事实上,由于梦象的五花八门和难以验证,长期以来人类对于梦象的理解与描述几乎一直处于猜测和遐想水平。古埃及和美索不达米亚人相信梦源天意,即上帝旨意通过梦境传达人类[1,2]。这一信念一直延续到古希腊和古罗马时期才逐渐发生改变。
我国古代的王廷相提出“因衍之感”,即人在睡眠中,无意识不自觉地运用“旧之所历,昼之所为”保存在记忆中的材料进行联想、想象和构思,这一过程称为因衍。20世纪初奥地利心理学家弗洛伊德致力于梦象的心理分析,提出所谓梦的六大特征,即原始性、冲动性、非逻辑性、非时空性、非道德性、非语言性。他在《梦象解释》一书中提出了风靡一时的梦象夙愿得偿(wish-fulfillment)的学说[3]。
有关睡眠和梦象的现代神经生物学研究起于20世纪50年代初快眼动(REM)睡眠的发现[4]以及梦境和 REM睡眠关系的确认[5]。半个多世纪以来,有关睡眠和梦象的研究与探讨方兴未艾,成为神经科学、心理学和神经精神学等多学科的共同热点之一。早期的实验研究大多集中于睡眠和梦象产生的神经生物学机制[6-8],后来扩展到梦象的心理现象的分析[9,10]及其与神经和精神疾患的联系[11,12]。另一些学者则收集了大量梦境,对其内容进行分析,得出了影响比较广泛的梦境荒诞(dream bizarreness)学说[13,14]。梦境荒诞离奇几乎人人都有体会,即便所谓有理性、近思维的梦境也常常不合逻辑,与实际相去甚远。但梦象何以如此荒诞离奇只有一些心理方面的分析,还未见神经生物学证据的报告。
迄今为止,脑电图(EEG)仍为梦象研究中最普遍采用的实验手段[6-9]。生理睡眠周期的最后阶段,即 REM 睡眠被证实为有梦睡眠期[4,5]。为了探讨梦境荒诞离奇的根源,本研究采用实验神经生物学手段,对脑神经活动的最基本现象—脑电图进行记录、比较和分析,并从生物进化的角度对梦象的本质进行探讨,而不涉及梦象的具体内容及其可能的心理学意义。
1 材料与方法
1.1 实验记录与观察
实验记录与观察是对人和动物的脑电图和所伴随的行为活动进行连续观察和记录,由渭南师范学院心理梦象研究中心与渭南市中心医院神经内科两方面合作进行。
实验对象分三组:一是渭南市中心医院内科收住的非脑部疾病患者(n=3,年龄15-35岁,每例检测1-3次);二是在渭南师院校大学生志愿者(n=4,年龄18-25岁,每例检测2-3次);三是被试动物,雄性健康家兔(n=4,每例检测2次)。每组观察对象在不同时间重复进行记录观察,单位以人次或次计。全部实验记录观察均在2008-07~2008-11期间完成。非脑部疾病患者脑电图资料在2008-02~2009-10期间收集。
实验记录由脑电图技师使用多道脑电图仪连续记录被试人员和家兔脑电图,以24 h为单位。按文献惯例,我们把睡眠周期简化为非快眼动(NREM)睡眠和快眼动(REM)睡眠期。从清醒转入睡眠,经由一个短暂过渡期(也称昏睡期)进入NREM期,由浅逐渐转深,进入通常所称的熟睡期或慢波睡眠(SWS)期;再转入REM期,即通常所谓的有梦睡眠期。如此周而复始,每晚重复3-5次,其相关参数亦分别统计。我们还同时观察受试对象的行为变化(可逆性意识丧失),并结合脑电图的变化对观察对象的睡眠周期和阶段进行判断。结果分析脑电图由脑电图技师测量;结果由实验设计者进行对比和统计分析。
1.2 检测验证
检测验证部分包括两个步骤。一是由设计者分别计算出人脑除大脑、小脑和延髓外之脑干复合体和边缘系统(通常所谓的“人脑生梦部位”包括爬虫复合体[15],即脑干复合体、边缘结构和枕叶皮层)的体积数、重量数、神经元数、可容比特数、所含基因数。二是由设计者分别计算出家兔脑相应结构之体积数、重量数、神经元数、可容比特数、所含基因数,并将两组数据进行比较分析。我们对3只福尔马林灌注的成年家兔的脑组织的体积进行了测量,以此计算其信息容量。而成人脑体积容量引自教科书或专著[16,17],并以此为基础进行计算。
1.3 数据处理
脑电图技师测量的健康成人和家兔不同生理状态下的脑电图波形参数(主要包括波形的幅度和频率)以±s表示。数据由实验设计者进行统计学处理和比较分析。测得的两组数据(幅度和频率)先进行双项变异分析(ANOVA)检验,再进行后Dunnett比较。显著性水平(P值)取0.05为界(双侧)。
2 实验结果
2.1 正常人EEG记录
我们观察到,清醒时的脑电图为低幅快波,并混有与外界传入信号相关的高幅慢波。入睡后即进入NREM睡眠。此时的脑电波变得平稳,会出现单一的睡眠梭(时相性快波),并逐渐过渡到SWS期。最后进入REM睡眠期。此时的脑电图以不规则低幅快波为主,其脑电波频率比清醒时慢一些,常混有少量快波,类似思睡期的脑电图。此期间的特征性行为表现是快速眼球运动(REM),并伴有频繁而不自主的肢体抽搐(生理性的睡眠肌阵挛)。
清醒和睡眠状态由实验观察者按被试者的脑电图和行为表现予以区分记录。观察结果表明,由清醒状态进入熟睡期,即脑电图由低幅快波变为高幅慢波,是一个逐渐转变过程,通常需要20-40 min,平均(28.5±4.7)min;n=12人次。随后高幅慢波不断增多,大约30 -40 min,平均(35.7 ±7.8)min后慢波突然消失,转变为低幅快波,即进入REM期,此期持续约5-25 min ,平均(18.1 ±11.7)min,再进入下一个NREM期或转入苏醒状态。这样的睡眠周期每晚重复4次(7人次,70%),5次(2人次,20%)或3次(1人次,10%)。随着睡眠的进程,在每个周期内,NREM时程逐渐减少而REM时程逐渐增加。测量数据显示,正常健康成人的脑电波幅度和频率在清醒状态下分别为(2.2±0.35)μV和(9±2.1)Hz,22人次;非快眼动或无梦睡眠状态为(8.8 ±3.2)μV 和(1.5 ±0.78)Hz,12 人次;快眼动或有梦睡眠状态(2.1 ±1.5)μV 和(5.5 ±1.5)Hz,12人次(见表1)。这些数据与我们以往积累的多例非中枢神经系统患者(n=7人次)的脑电图一致。
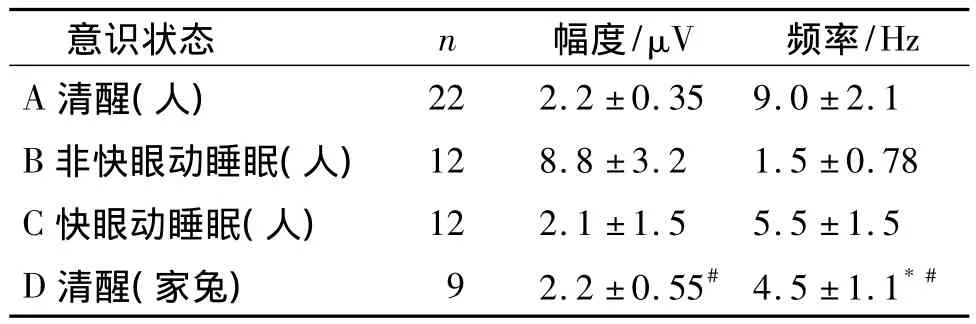
表1 健康成人和家兔不同意识状态下的脑电图参数比较Table 1 Measurements of EEGs of healthy volunteers and rabbits under different conscious status
2.2 健康家兔EEG记录以及与人EEG的比较
进一步测量结果显示,家兔清醒状态下的脑电图波幅和频率分别为(2.2 ±0.55)V 和(4.5 ±1.1)Hz(n=9)。将人和家兔的脑电图测量参数进行统计学分析,表1结果显示,与家兔清醒状态下的脑电图波幅度相比,人清醒和有梦睡眠时与其相当(P>0.05),但熟睡状态则显著偏高(P <0.05);与家兔清醒状态下的脑电图频率相比,人有梦睡眠时与其相当(P >0.05),但清醒时则显著增快(P <0.05),而熟睡时则显著偏慢(P<0.05);如果将脑电图波的幅度和频率综合考虑,只有人有梦睡眠时的脑电图像和家兔清醒时的脑电图像最为相似或无显著差异(P >0.05)。
图1为节选的受试人员枕叶与被试家兔额叶的脑电图图像。其中图A,B分别显示受试学生在清醒状态(AWAKE)、熟睡状态(NREM或SWS)和有梦睡眠状态(REM)期间的脑电图。图C,D分别为节选的2例家兔清醒状态之脑电图。将这些具有代表性的脑电图图像进行直接目测法比较发现,家兔AWAKE时的脑电图和正常健康成人AWAKE时的脑电图幅度相当但频率偏慢,与正常健康成人NREM睡眠期相比,幅度明显偏小而频率较快,但与正常健康成年人REM睡眠期,即有梦睡眠状态时脑电波幅度和频率均十分接近。
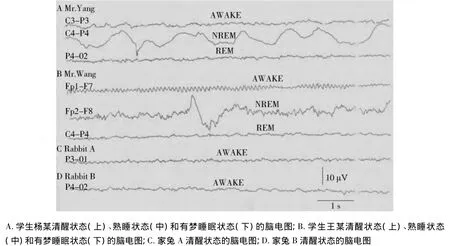
图1 健康成人和家兔不同生理状态下的脑电图变化Figure 1 Changes of EEG of healthy volunteers and rabbits under different conscious status
2.3 实验检测
通过测量3只福尔马林灌注的成年家兔的脑组织体积,成人脑体积容量引自教科书或专著[16,17],以此计算出的信息容量如下:①人脑体积如不把小脑(主管运动的协调与控制)和丘脑(感觉传入)计算在内大约有100亿神经元并含有大约10万亿比特。而大脑皮质占整个人脑皮质的96%,因而人脑的边缘系统与爬虫复合体再减去延髓部分,其神经元约为3.5亿,其信息容量约为3 500亿比特。②成年家兔脑的体积大约是13 cm3,拥有几亿神经元和几千亿比特。③人的遗传基因数与哺乳动物(包括家兔)基因数均为10亿比特左右。
3 讨论
动物实验发现,REM期的特征性脑电活动是起源于脑干复合体的脑桥并经丘脑外侧膝体到达大脑枕叶皮层的特征性电位波,即所谓脑桥-外膝体-枕叶皮层(PGO)波,这被认为是触发REM期梦境产生的神经生物学基础[6-8]。在睡眠及睡梦的实验观察中,脑电图记录一般是多道连续记录,但在观察结果和测量分析时大多选取受试人枕叶皮层(即视觉皮层)的图像[8]。动物实验中,家兔脑电图的记录方法亦相似—多道连续记录。资料分析则多选取大脑前额叶的图像[18]。本实验在收集人和动物脑电图资料时亦采用多道连续记录;结果测量和分析也分别节选人脑枕叶区和家兔大脑前额叶区的记录图像。此外,脑电图记录得以普及应用的原因之一在于其无创伤性。但通常由颅外记录到的脑电信号幅度较小且个体差异较大,但其频率比较稳定,个体差异不大,能比较可靠地反映受试者(人和动物)记录部位脑区的神经电生理活动状态。所以,我们所选用脑电图及其记录方法,包括受试人和动物,都具有恰当的代表性,符合本实验的目的和文献惯例。需要指出的是,家兔脑电图亦随其睡眠周期而变化,并伴有相应的行为改变[15];但我们所感兴趣的是该动物清醒状态下的脑电图。有趣的是,家兔在清醒安静状态下的脑电图和REM睡眠时的脑电图不仅图像相似,其脑电波能谱亦几乎完全相同[19]。
在现代神经科学研究中,对神经元形态的观察已深入到树突棘和轴突终扣(二者均小于1 μm)以及亚细胞水平。对于一个结构均匀的神经核团的神经元数目也能做出比较准确的估计。但大脑是一个结构十分复杂的庞大器官,对其神经元的总数目至今未有一个令人信服的统计数据。相反,现有的有关大脑总体及其相关结构和某一种神经元的数目及其所形成的突触数也都处于“估计”水平,各家所持数据差异甚大。我们的分析验证主要是从宏观水平着手,对大脑中与梦象相关的神经结构以及家兔相应中枢神经结构及其信息容量予以大体估算,并借以窥测这两个不同种属大脑中相应结构在从生物进化过程的平行和接近程度。比特(bit,binary digit)于20世纪60年代用于神经信息编码的量化描述。严格地讲,只有“全”或“无”式的动作电位符合这种二进制的数学描述;神经元之间的信息传递主要通过突触完成,而突触之间的信息传递符合概率规律。这样,比特只能用于描述神经网络信息活动的特殊状况—最大信息容量,大致相当于神经网络的突触数量。我们的检测数据表明,人脑生梦部位,即脑干复合体(亦称爬虫复合体)、边缘系统和相关大脑皮层与家兔相应脑结构体积数类似、重量数类似,神经元数类似,信息容量类似,基因容量类似。这些相似性提示,类似的神经功能或活动过程也可能在这两个完全不同的种属相应的中枢结构系统中分别进行、发生。
本实验记录观察了人脑清醒、无梦睡眠和有梦睡眠三种生理状态下的脑电图和家兔清醒状态下的脑电图,并将其进行测量比较。结果发现,人清醒状态下的脑电图与家兔清醒时的脑电图明显不同;人无梦睡眠时的脑电图与家兔清醒时之脑电图差别很大;而人有梦睡眠时的脑电图与家兔清醒之脑电图像十分相似(图1)。我们的统计数据也表明这一点。家兔清醒时的脑电图与人有梦睡眠时的脑电图十分类似,而与另外两种状态下人脑电图明显不同,这些观察结果和我们实验假设相符,即“人在梦境中的决断具有动物性”。需要指出的是,本文的“决断”是指认知与抉择,即现代汉语中拿主意、做决定之意。所谓“动物性”是指生物学公认的主宰动物生存所必需的行为,即食欲、性欲、生存欲,与文献中广泛使用的梦境荒诞离奇所指相近。
现已清楚,梦境不仅发生在REM睡眠期,亦可在NREM睡眠阶段产生。但两个不同睡眠阶段所发生的梦境的内容和频率都不同。NREM睡眠期的梦境往往带有一些理性的想法而没有明显的人物与情节;最新研究还表明,NREM睡眠与学习和记忆密切相关[20]。与此相反,REM睡眠期的梦境大多荒诞杂乱,不符合常理。后者则是本文所研究和探讨的对象。这些资料提示,如果不把梦境的神经生物学基础阐明,不把梦象按其发生的睡眠期进行合理分类,而试图对所有的梦象内容进行心理学解释,必然得出相互矛盾、无法统一的结果。
梦象通常被视为一种心理现象,能否用神经生物学手段进行观察和研究曾一直没有定论。20世纪50年代初REM睡眠与梦境活动的关系得到确认[4,5],随后大量的动物实验研究结果和临床观察表明,参与梦境活动的中枢神经结构包括脑干脑桥复合体(不含延髓)、丘脑、部分大脑皮层及边缘系统等中枢神经结构[16]。与清醒状态下的大脑意识活动在大脑皮层前额叶的协调和控制下进行不同,睡眠期间大脑前额叶处于深度抑制状态;在REM睡眠期,起源于脑干中部脑桥的PGO波经丘脑外膝体到达枕叶皮层的同时,直接或间接地激活边缘系统的下丘脑、杏仁核等边缘结构。这一上行神经系统的活动被认为是梦境发生的神经生理学基础[6-8]。Llinás等认为,睡眠时大脑与外界的联系中断,但大脑是一个从不“休息”的器官,梦境则是大脑自主活动的表现[21]。Hobson等认为,梦境是大脑在不受外界“干扰”情况下自主活动、合成的“副产物”,不具任何生物学意义[6]。Cavallero等也认为,梦境并不创造任何新东西,只是把原有的记忆信息以原始的方式结合在一起而已[22]。这些观点都从神经生物学角度对梦境的发生原理进行了剖析,认为梦境不是大脑正常活动的生理性产物;而我们的研究则是采用实验的手段,提供了梦境荒诞离奇的神经生物学证据。总之,这些资料表明,失去大脑意识活动的协调和控制作用是人梦境离奇荒诞的神经生理学基础。
脑生物电是大脑生理功能活动的基础,深入研究人脑的生物电活动规律和机制是破译人脑功能密码的必要途径,因而脑电图是研究人脑科学奥秘的重要工具之一。通常所谓的脑电图就是通过记录电极在头皮某一部位记录到相应位置深部大脑皮层细胞群的自发性、节律性突触后电活动情况。不同脑区或同一脑区不同功能状况下的脑电图不同,脑结构相似的动物种类之间的脑电图也可能相似。人和家兔及猫等哺乳动物的大脑皮层均为5-6层的板层结构,其脑电波的基本图像也相似,相似的图像变化过程也反映了同样或相似的神经生物活动现象。同理,家兔清醒之脑电图与人有梦睡眠之脑电图相似,说明家兔脑与人脑生梦部位(被称之为古皮质、旧皮质的边缘系统与爬虫复合体[15])的结构复杂程度以及功能的进化程度类似,而与此相关的信息整合的内容和方式亦应类似或接近[16]。因之,人有梦睡眠期间参与梦境发生的脑区的神经生物活动过程具有动物性,像动物那样整合的储存记忆与现有的体内外刺激而形成梦是很自然的。
动物不能用语言表达其梦象的内容,甚至难以判定其有无梦境发生。但哺乳动物,包括猫、狗、家兔和鼠类的睡眠周期性和阶段性以及REM睡眠时的脑电、肌电、眼球运动及体温的动态变化几乎和人体完全一致[7,8],提示哺乳动物在REM睡眠时很可能有梦境产生。最近有人指出至少狗会做梦[23]。众所周知,动物实验是理解和阐明人体功能活动及其机制的最基本最常用手段,前人桑代克研究人的心理机制、巴甫洛夫研究人脑神经机能都是从动物实验着手。我们研究人梦像的特性用动物实验(家兔)进行观察比较,从原则上讲也应该是可行的。按逻辑思维方法,人脑生梦部位和家兔脑相关结构是可以进行比较的。我们根据和人梦境形成有无联系的标准,把和人梦有关的脑结构与家兔脑相关部位进行对照比较,得出二者在神经形态方面的相似性,从而在此方面属于同类,并得出人生梦部位的脑结构与家兔相应的脑部在功能方面的相似性,继而得出人脑生梦部位具有动物性属性的结论。这样,我们不仅按比较的要求对事物内部矛盾的各方面进行了一般的比较,而且进行了事物内部的深入到细胞水平的微观结构有关方面对比,如可容信息数、可容基因数等方面的对比,以增加我们实验假说、验证方法和所得结论的可靠性。卡尔·萨根指出:“梦态中类似动物所做的事可以理解是爬虫复合体[17]和边缘系统的活动,偶尔也有新皮质右半脑活动的强烈直观见解……从进化来看,做梦是哺乳动物的原始功能”。这一观点和本研究的假设、方法和结论十分吻合。
最后需要指出的是,上述起源于脑干的PGO波既触发梦象发生,也能在大脑的某些区域诱发可塑性变化[24,25]。已知神经元可塑性变化是大脑学习记忆活动在细胞水平的反映[20,26];已有实验资料表明,有梦睡眠期间,当日的程序性训练,即运动技巧学习,得到加强和巩固;如果人为地压抑PGO波的发生,其运动技巧的获得也变得缓慢[27,28]。但目前尚无梦境本身和运动技巧的获得有任何联系的直接实验证据。
致谢:参与本研究的实验技术工作的有渭南师院陈兰萍、梁艳玲、孟宗宏、马雅菊、王亚娟和赵姝萍;渭南市中心医院张志俊、张进社和程月芳。谨对为本实验与研究提供帮助的渭南师院科研处王双喜教授、孙红梅干事,物理系王菊侠教授,化学系刘亚强教授,生命科学系卢爱岗主任、张万海副教授,教育科学系傅俊贤教授、秦文孝教授、董建稳副教授,体育系张虎主任、秦华副教授,图书馆原馆长刘志涛先生、杨晓云副教授,数学系王云山副教授、张泉林副教授,艺术系王新惠教授、外语系王仰义副教授致以衷心的感谢。感谢西雅图儿童研究所韩中胜博士和浙江大学心理系翁旭初教授惠阅本文初稿并提出修改意见。
[1] Noegel S.Dreams and dream interpreters in mesopotamia and the hebrew bible(old testament)[M]//Bulkeley K,ed.Dreams:a reader on the religions,cultural and psychological dimensions of dreaming.New York:Palgrave,2001:45 -71.
[2] Szpakowska K.Through the looking glass.Dreams in ancient egypt[M]//Bulkeley K,ed.Dreams:a reader on the religions,cultural and psychological dimensions of dreaming.New York:Palgrave,2001:29 -43.
[3] Freud S.The interpretation of dreams[M].Modern library edi.Toronto:Random House Inc,1994.
[4] Aserinsky E,Kleitman N.Regular occurring periods of eye motili-ty and concomitant phenomena,during sleep[J].Science,1953,118:273-274.
[5] Dement W,Kleitman N.The relation of eye movements during sleep to dream activity:an objective method for the study of dreaming[J].J Exp Psychol,1957,53(5):339 -346.
[6] Hobson JA,McCarley RW.The brain as a dream state generator:an activation - synthesis hypothesis of the dream process[J].Am J Psychiatry,1977,134(12):1335 -1348.
[7] Hobson JA.REM sleep and dreaming:towards a theory of protoconsciousness[J].Nat Rev Neurosci,2009,10(11):803 -813.
[8] Staunton H.Mammalian sleep[J].Naturwissenschaften,2005,92(5):203-220.
[9] Staunton H.The function of dreaming[J].Rev Neurosci,2001,12(4):365-371.
[10] Reiser MF.The art and science of dream interpretation:Isakower revisited[J].J Am Psychoanal Assoc,1997,45(3):891 -905.
[11] Solms M.New findings on the neurological organization of dreaming:implications for psychoanalysis[J].Psychoanal Q,1995,64(1):43-67.
[12] Blagrove M,Pace-Schott E.Trait and neurobiological correlates of individual differences in dream recall and dream content[J].Int Rev Neurobiol,2010,92:155 -180.
[13] Limosani I,D’Agostino A,Manzone ML,et al.Bizarreness in dream reports and waking fantasies of psychotic schizophrenic and manic patients:Empirical evidences and theoretical consequences[J].Psych Res,2011,189:195 -199.
[14] Hobson JA,Hoffman SA,Helfand R,et al.Dream bizarreness and the activation - synthesis hypothesis[J].Hum Neurobiol,1987,6(3):157-164.
[15] Newman JD,Harris JC.The scientific contribution of Paul D MacLean(1913 -2007)[J].J Nerv Ment Dis,2009,197(1):3 -5.
[16] Brodal P.The central nervous system-structure and function[M].2nd Ed.Oxford:Oxford University Press,1998:553 -620.
[17] Sagan C.The dragons of eden:Speculations on the evolution of human intelligence[M].2nd Ed.New York:Ballantine Books,1986:19-50.
[18] Pivik RT,Bylsma FW,Cooper P.Sleep-wakefulness rhythms in the rabbit[J].Behav Neural Biol,1986,45(3):275 -286.
[19] Yamamoto J.Roles of cholinergic,dopaminergic,noradrenergic,serotonergic and GABAergic systems in changes of the EEG power spectra and behavioral states in rabbits [J].Jpn J Pharmacol,1988,47(2):123 -34.
[20] Plihal W,Born J.Effects of early and late nocturnal sleep on declarative and procedural memory [J].J Cogn Neurosci,1997,9(4):534-547.
[21] Llinás RR,Paré D.Of dreaming and wakefulness[J].Neuroscience,1991,44(3):521 -535.
[22] Cavallero C,Cicogna P,Bosinelli M.Mnemonic activation in dream production[M]//Koella WP,Obal F,Schulz H,et al.Sleep.London:Gustav Fisher,1986:91 -94.
[23] Miller K.Do animals dream[J]?Popular Science,2012,(2):71.
[24] Datta S.Avoidance task training potentiates phasic pontine-wave density in the rat:A mechanism for sleep-dependent plasticity[J].J Neurosci,2000,20(22):8607 -8613.
[25] 房季冬,冯平福.睡眠和觉醒—神经科学[M].北京:北京大学医学出版社,2009:904-919.
[26] Rasch B,Born J.About sleep’s role in memory[J].Physiol Rev,2013,93(2):681 -766.
[27] Legault G,Smith CT,Beninger RJ.Post-training intra - striatal scopolamine or flupenthixol impairs radial maze learning in rats[J].Behav Brain Res,2006,170(1):148 -155.
[28] 韩中胜,史均翰.睡眠、梦境的机制及其与学习记忆的关系[J].生理科学进展,2013,44(6):409 -414.
