长白山不同海拔树木生长对气候变化的响应差异
2014-05-05尹云鹤赵东升苑全治吴绍洪
陈 力,尹云鹤,赵东升,苑全治,吴绍洪,*
(1.中国科学院地理科学与资源研究所,北京 100101;2.中国科学院遥感与数字地球研究所,北京 100094)
由于树木年轮具有定年准确、分辨率高、分布广泛和易于获取等特点,因此成为气候变化研究的重要资料之一。研究表明,山地垂直带对气候变化反应的灵敏度远高于水平带[1],在山区树木生长对气候响应可能与海拔梯度相关[2-4]。通常认为高海拔或近林线处的树木对夏季气温有较强的响应,而低海拔的树木对降水有正响应[5-6]。但在中亚干旱和半干旱地区[7-8]以及阿根廷西北部地区[9]等地开展研究所得的结果与普遍认识并不一致,诸如高海拔树线处树木生长受降水而非夏季气温影响[9]。这些案例表明不同海拔树木生长变率还受到区域气候变率的影响。因而充分认识海拔梯度上树木生长对气候的响应机制对树轮气候学研究具有科学意义,同时也有助于评价和预测森林生态系统对未来气候响应。
东北地区是近年来气候变化较为敏感的区域之一[10]。长白山是东北地区最高山,垂直梯度大,目前有针对上限/下限树木生长对气候变化响应研究[11-14],但是针对整个海拔梯度的树木响应研究并不多见。已有研究发现红松分布下限对春季气温响应明显[11],上限长白落叶松对当年气温和降水均有较好响应[13],红松和长白落叶松分布上限/下限对气候响应差异明显[14]。本研究通过获取长白山北坡不同海拔长白落叶松(Larix olgensis)和红松(Pinus koraiensis)树轮资料,研究在海拔梯度上树木生长变化,探讨不同海拔树木生长对气候的响应,以期丰富该地区树轮气候学研究内容。
1 研究区概况与研究方法
1.1 研究区概况
长白山气候特征是具有温带大陆性山地气候,夏季温暖多雨,冬季寒冷漫长。因山体高,气候随海拔变化较大,山下年均温约2.8℃,而山顶年均温约-7.3℃,山下年降雨量为 600—900 mm,山顶为1300—1500 mm。随海拔上升,植被呈现垂直分布带谱,大致分为:低山针阔叶混交林带(500—1100 m)、山地针叶林带(1100—1800 m)、亚高山岳桦林带(1800—2100 m)和高山苔原带(2100 m以上)。
长白落叶松是长白山乡土树种,垂直分布范围在海拔 500—1950 m[15],4 个采样点分别为 L1、L2、L3和L4(其中L4样点的样本源于参考文献[11])。红松是长白山建群树种,垂直分布范围在海拔500—1300 m,6 个采样点分别为 H1、H2、H3、H4、H5 和H6。采样点位于天然次生林,郁闭度达0.7。利用生长锥取样,在每棵树胸高处不同方向取两个树芯作为样本,个别树木取了3个树芯,取样时考虑满足大样本量的要求,每个采样点采集20—30棵树。树芯取出后,放进事先卷好的纸管中,做好编号。采样点概况见表1。
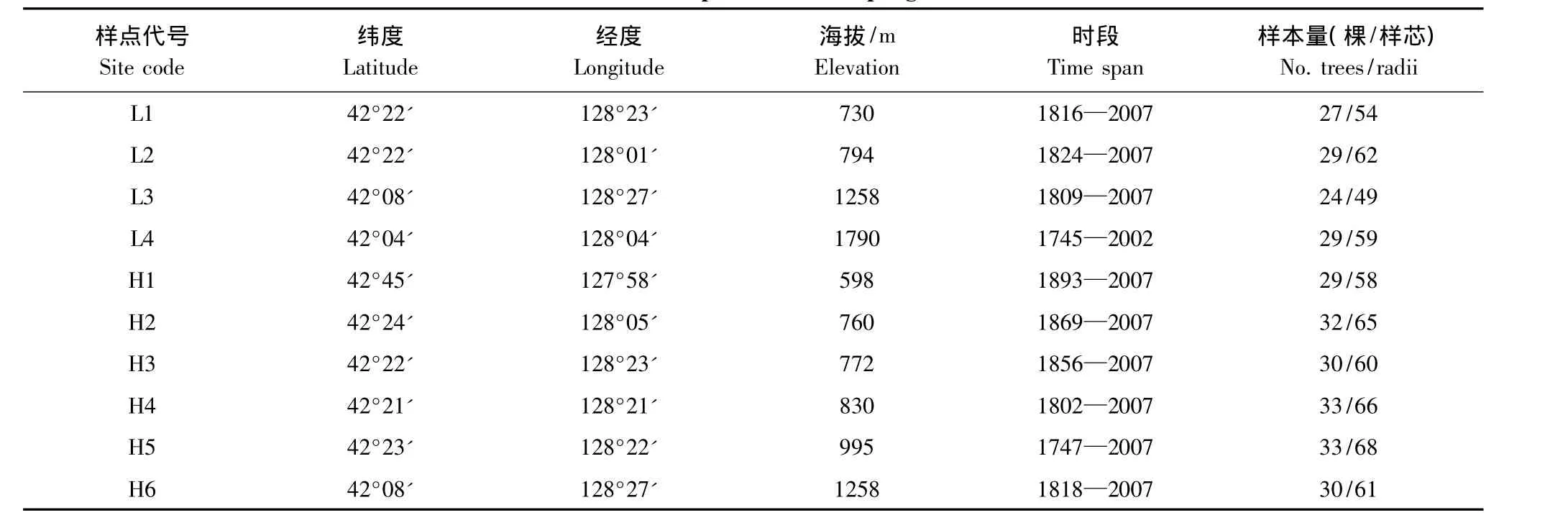
表1 树轮采样点概况Table 1 Description of the sampling sites
1.2 树轮年表建立
将样本晾干、粘贴和打磨,直至达到树木年轮学分析的要求,然后进行交叉定年和树轮宽度量测。量测采用德国Frank Rinn公司生产的LINTAB轮宽量测仪,精度为0.01 mm。为确保定年和量测的准确性,用COFECHA程序[16]对量测结果进行检查,剔除不能交叉定年的序列。
由于本研究采样时间是2008年6月,所以树轮完整宽度的最后一年是2007年。通过ARSTAN程序[17],采用步长为2/3序列长度的样条函数去掉生长趋势和树木之间因竞争造成的低频波动,最终得到标准年表(STD)和差值年表(RES)。为了评价年表的质量,计算了年表的平均敏感度(MS)、标准差(SD)和一阶自相关系数(AC1)等,并对年表进行了共同区间分析,选取时段1900—2002年(长白落叶松,因L4年表截止年份是2002年),1900—2007年(红松)。
1.3 气象资料
本研究采用的气象资料来自二道气象站(42°53'N,128°25'E,海拔 591 m,原为松江气象站,2005年迁站并更名)。在气候分析之前,采用 Mann-Kendall[18]法对气象数据进行均一性检验,经过检验确认数据可靠,气温和降水数据资料变化相对均一。考虑到前一年的气候状况对当年树木生长的影响,分析时选取上年7月到当年9月的逐月平均气温和降水量。
1.4 研究方法
利用统计软件SPSS19分别对长白落叶松年表(1900—2002年)、红松年表(1900—2007年)进行聚类分析,检测树种空间格局的一致性[19]。根据相似系数的距离来衡量采样点之间的亲疏关系,分别选择不转换数据、皮尔逊相关和最远邻法等选项设置数据转换方式、聚类相似性和聚类方法等系统聚类参数。
利用软件DendroClim2002[20]对年表与气候因子进行相关分析和响应分析。响应分析是通过主成分分析去除气候因子间自相关,再与年表做逐步回归,并将各主分量的回归系数转换成对应原始气候资料的回归系数,以回归系数的大小和正负表示树木生长对气候因子的响应程度。显著性检验用Bootstrap方法[21]随机执行 500 次。
2 结果与讨论
2.1 沿海拔梯度的树轮年表统计特征
表2列出长白落叶松和红松差值年表的统计量及共同区间分析结果。
对于长白落叶松,由表2可见在共同区间内,相关系数(r1,r2,r3)和样本总体代表性(EPS)值较高,表明树木生长所指示的环境一致性较好。平均敏感度(MS)衡量相邻年轮之间宽度变化,可以反映包括气候因子在内的环境的短期(或高频)变化,MS值高可以指示年表所保留的气候信息丰富。L4年表敏感度高于其他年表,反映生长在高海拔的树木对环境变化的敏感性高。针对马尾松(Pinus massoniana)[22]、青海云杉(Pcicea crassifolia)[23]等树种的年表分析也得出类似结论。年表间信噪比(SNR)差异较大,其中L2年表最高,L1年表最低。各年表pc1范围是34%—50%,表明其共性较强。L2与L4的 EPS、SNR和第一主成分解释方差量(pc1)较大,L1与L3较小。L1与L2样点海拔差异不大,但是二者的统计量(MS、r1、r2、r3 和 SNR)差别明显,可能原因是L1样点位于沼泽苔草地带,小环境造成树木生长同步性不强。总体而言,随海拔增高,长白落叶松年表特征值表现出先下降后增加的趋势。
对于红松,比较表2各样点差值年表可见,平均敏感度(MS)范围是0.141—0.217,其中H5年表最高0.217,其余样点年表MS都不高于0.20,H6年表最低0.141,表示生长在亚高海拔的红松对环境变化的敏感性更高。对于参数SNR、EPS、PC1,H4最高,H2次之,H1最小。总体而言,随海拔增高,红松年表特征值呈先增加再下降的趋势。
由上述分析可知两个树种对环境的敏感性均较强,但在空间尺度上,两个树种年表特征随海拔变化有所不同,分布高海拔的长白落叶松比林内的树木对环境响应更敏感,而红松则是相反的情况。这可能反映了树木遗传学特点不同。
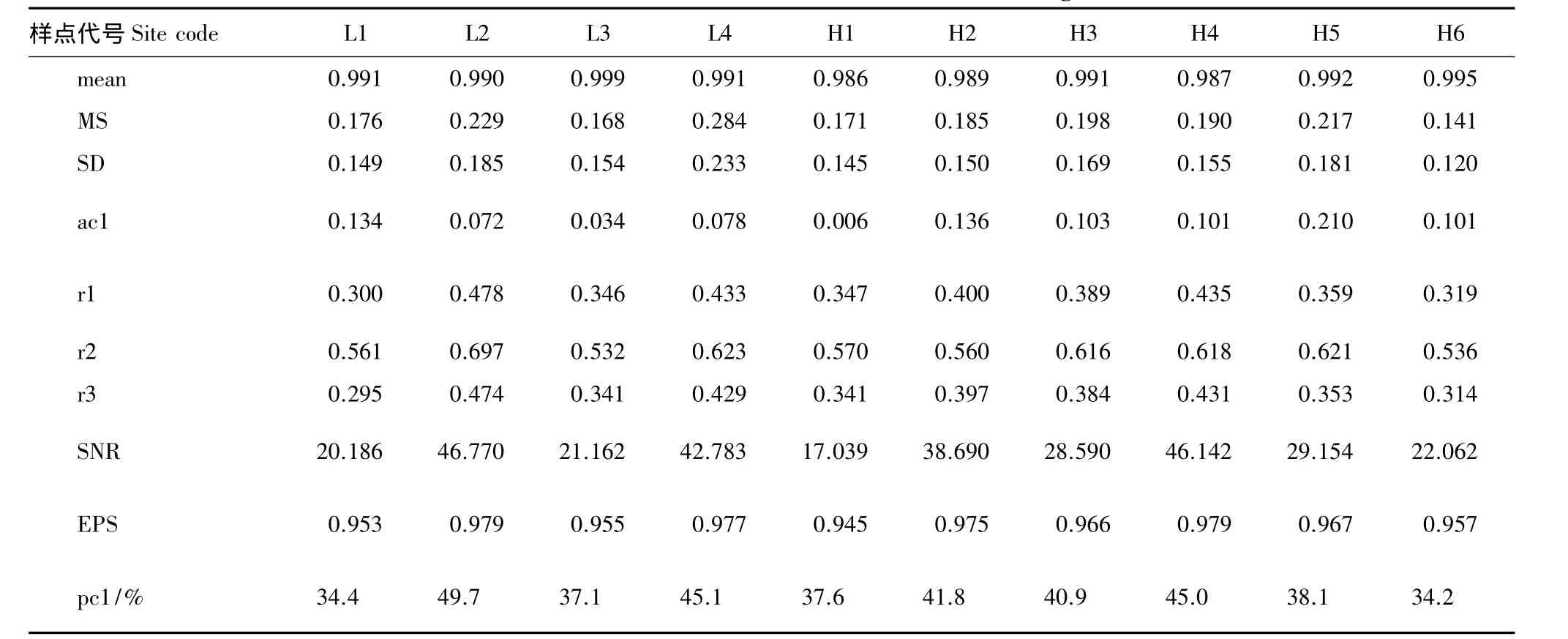
表2 树轮年表及共同区间统计量Table 2 Statistics of common interval of residual chronologies
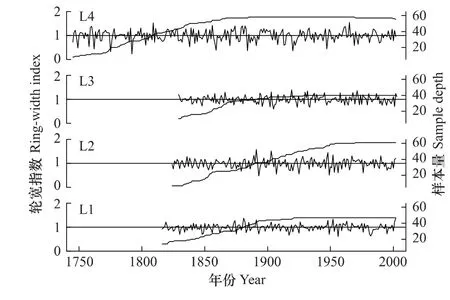
图1 长白落叶松各样点树轮宽度差值年表Fig.1 Ring-width chronologies of Larix olgensis
通常认为EPS值大于0.85的时段为可信时段,图1、图2分别为长白落叶松和红松各样点自EPS>0.85时段起的差值年表及各时期样本量。L4年表同其他样点年表表现出不同的变化特征,各曲线的峰值不一致(图1)。在年际变化上,H3、H4波动幅度较强,而H6波动幅度微弱(图2)。
2.2 树木生长空间格局
海拔变化造成热量与降水的重新分布,可能影响树木生长格局,图3是反映长白落叶松聚类全过程的树形图。4个年表聚为两类时,类间距离较大,表示样点间树木生长亲疏关系不同。第1类是高海拔年表L4,第2类是中低海拔年表L1、L2和L3。高海拔树轮年表序列明显长于低海拔。使用ARSTAN程序将第2类的各样点年表融合,建立新年表。

图2 红松各样点树轮宽度差值年表Fig.2 Ring-width chronologies of Pinus koraiensis
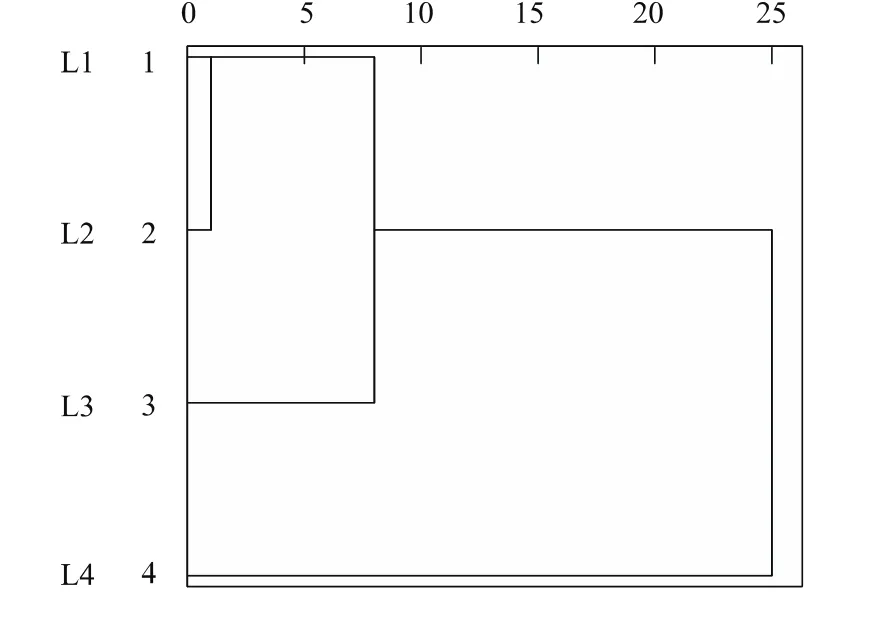
图3 长白落叶松年表聚类分析树形图Fig.3 Dendrogram of hierarchical clustering analysis of chronologies of Larix olgensis
图4是反映红松聚类全过程的树形图。6个年表聚为三类时,类间距离较大。各类组成为:第1类是高海拔年表H5和H6;第2类是中海拔年表H3、H4、H2;第3类是低海拔年表H1。采用ARSTAN程序分别将第1、2类的各样点年表融合,建立新年表。
2.3 不同海拔树木生长对气候变化的响应
长白落叶松两类年表与气候要素响应分析结果如图5所示。第1类年表(即高海拔长白落叶松)与当年3月、6月气温分别呈显著负、正相关(图5)。上年气候要素与年表的相关系数均未达到显著水平。这与以往研究[12-13]发现生长在高海拔的长白落叶松包含较强的温度信号的结果相符。在高海拔样点处,3月是冬末,气温在-10℃左右,冬季气温增加使得树木呼吸作用增加,养分消耗增加,不利于物质累积。6月气温在8℃左右,气温增高促进形成层活动能力,增加光合产物,形成宽轮。
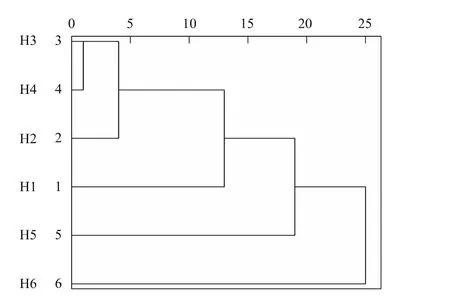
图4 红松年表聚类分析树形图Fig.4 Dendrogram of hierarchical clustering analysis of chronologies of Pinus koraiensis
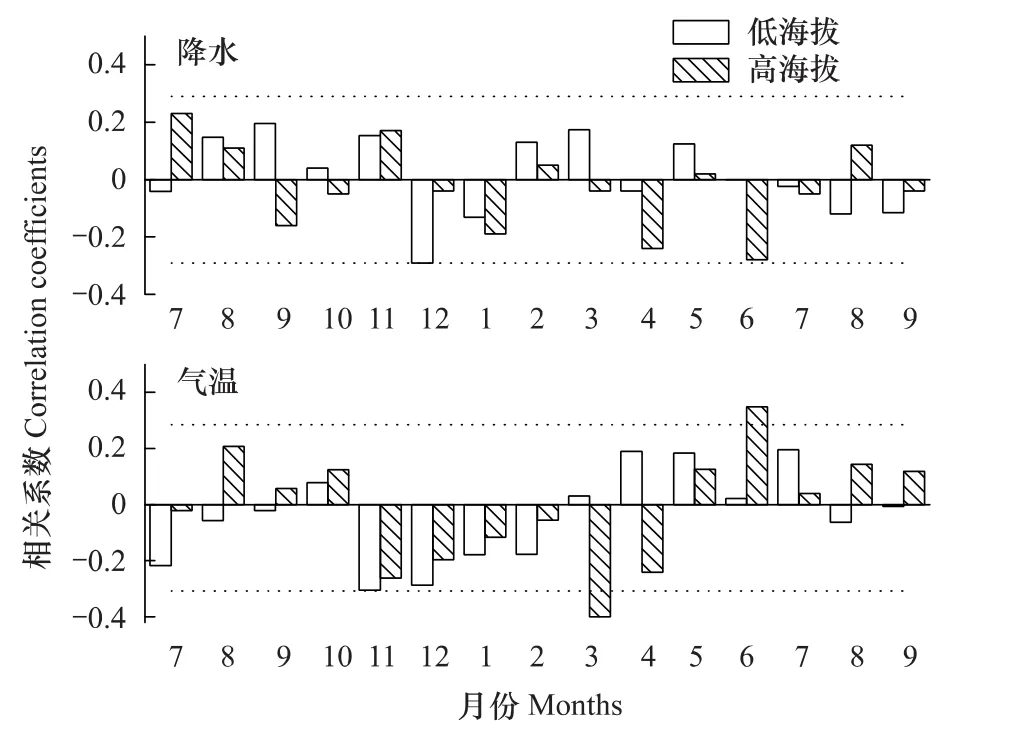
图5 长白落叶松年表与气候要素相关分析结果Fig.5 Results of correlation analysis for the relationships between the residual chronologies of Larix olgensis and monthly climatic factors图中虚线:P<0.05;7—9:上年7月—当年9月
第2类年表(即低海拔长白落叶松)与上年11月气温、上年12月降雪均呈显著负相关(图5)。年表与当年气温、降水的相关系数均未达到显著水平。由于长白山冬季寒冷多雪,如果11月气温偏高,则呼吸作用加剧,消耗累积光合产物,不利于来年径向生长[24]。同理,12月降雪偏多,树木呼吸作用增加,使冬季处于休眠状态的树木消耗增加,不利于营养物质存积,形成窄轮,这可能也体现长白落叶松耐旱性。
整体而言,水分能满足长白落叶松的生理要求,气温成为树木生长的主要限制因子,高海拔长白落叶松生长受当年气候影响,低海拔长白落叶松生长对气候存在“滞后响应”。长白落叶松采样点最高海拔1790 m,接近云冷杉林植被类型的上限,树轮中包含较强温度信号,这对重建历史温度也有一定的借鉴意义。
红松三类年表与气候要素响应分析结果如图6所示。第1类年表(即高海拔红松)与当年6月降水呈显著正相关,与上年10月气温呈显著正相关(图6)。6月份红松进入生长期,充沛的雨水能促进其生理活动能力,产生宽轮。上年10月气温(在2℃左右)对红松生长起限制作用,这可能表明秋季气温越高,光合作用越强,形成的光合产物就越多,有利于细胞壁物质的形成。
第2类年表(即中海拔红松)与当年4月、9月气温显著正相关,与上年7月降水显著负相关(图6)。在中海拔处,4月高温加速地温升高,促进根系活动和地上部分萌动;9月是红松晚材形成时期,高温有利于红松生长;7月进入生长季,是全年降水最多的月份,多雨影响光合作用的效率,相应缩短了生长时间,树体内存储的营养物质少,对来年生长不利。
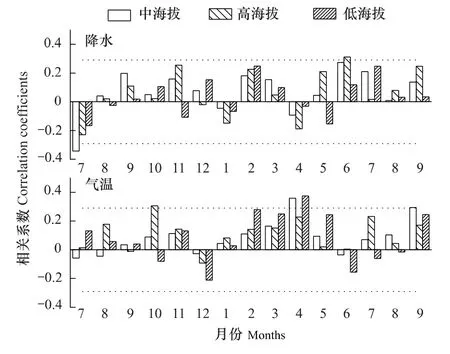
图6 红松年表与气候要素相关分析结果Fig.6 Results of correlation analysis for the relationships between the residual chronologies of Pinus koraiensis and monthly climatic factors
第3类年表(即低海拔红松)与当年4月气温显著正相关(图6)。结合中海拔红松响应结果,反映红松对初春气温较为敏感。王晓明等[25]发现4月最高气温升高有助于红松树轮生长。
整体看,气候影响整个海拔红松生长,高海拔红松不仅受降水限制,还对气温有“滞后响应”,中海拔红松不仅受气温限制,还对降水有“滞后响应”,低海拔红松生长主要受气温限制。红松采样点最高海拔1258 m,是针阔混交林植被类型的上限,但不是整个森林上限,因此研究结果不适用“高海拔或近林线处的树木对夏季气温有较强的响应”结论。
高海拔长白落叶松样点比红松的高约532 m,该样点的长白落叶松与气温显著相关,而对应的红松与气温、降水显著相关。低海拔长白落叶松样点与红松的高度相似,该样点长白落叶松生长与气温、降雪显著相关,红松生长与气温显著相关。该现象在一定程度上反映了长白落叶松耐寒,红松喜温的特点。
3 结论
基于树木年轮气候学分析方法,针对长白山地区的长白落叶松和红松生长对气候变化响应随海拔梯度变化进行了初步研究,得出如下结论:
(1)长白山北坡整个坡面各个采样点树木包含丰富的环境信息。随海拔升高,长白落叶松年表特征值呈先下降后增加的趋势,红松年表特征值呈先增加后下降的趋势。
(2)气候影响长白落叶松生长,其中气温作用更大。高海拔长白落叶松生长受当年气候要素影响,低海拔长白落叶松对气候要素有“滞后响应”。
(3)气候影响红松生长,高海拔红松不仅受降水限制,还对气温有“滞后响应”;中海拔红松不仅受气温限制,还对降水有“滞后响应”;低海拔红松生长主要受气温限制。
[1] WalterH.Vegetation ofthe earth.New York:Spring-Verlag,1984.
[2] Fritts H C,Smith D G,Cardis J W,Budelsky C A.Tree-ring characteristics along a vegetation gradient in northern Arizona.Ecology,1965,46(4):393-401.
[3] Hughes M K,Funkhouser G.Frequency-dependent climate signal in upper and lower forest border tree rings in the mountains of the Great Basin.Climate Change,2003,59(1/2):233-244.
[4] Tardif J,Camarero J J,Ribas M,Gutierrez E.Spatiotemporal variability in tree growth in the Central Pyrenees:climatic and site influences.Ecological Monographs,2003,73(2):241-257.
[5] Dittmar C,Zech W,Elling W.Growth variations of Common beech (Fagussylvatica L.)underdifferentclimatic and environmental conditions in Europe—a dendroecological study.Forest Ecology and Management,2003,173(1/3):63-78.
[6] Leal S,Melvin T M,Grabner M,Wimmer R,Briffa K R.Treering growth variability in the Austrian Alps:the inf l uence of site,altitude,tree species and climate.Boreas, 2007,36(4):426-440.
[7] Liu L S,Shao X M,Liang E Y.Climate signals from tree ring chronologies of the upper and lower treelines in the Dulan region of the Northeastern Qinghai–Tibetan Plateau.Journal of Integrative Plant Biology,2006,48(3):278-285.
[8] Esper J,Frank D C,Wilson RJS,Buntgen U,Treydte K.Uniform growth trends among central Asian low-and high-elevation juniper tree sites.Trees,2007,21(2):141-150.
[9] Morales M S,Villalba R,Grau H R,Paolini L.Rainfallcontrolled tree growth in high-elevation subtropical treelines.Ecology,2004,85(11):3080-3089.
[10] Wang Z Y,Ding Y H,He J H,Yu J.An updating analysis of the climate change in China in recent 50 years.Acta Meteorogical Sinica,2004,62(2):228-236.
[11] Zhu H F.A Temperature Reconstruction for Eastern Part of Northeast China Based on a Tree Ring Width Network[D].Beijing:Beijing Normal University,2006.
[12] Wu X D,Shao X M.A preliminary study on impact of climate change on tree growth using tree ring-width data.Acta Geographica Sinica,1996,51(Supplement):92-101.
[13] Yu D P,Wang S Z,Tang L N,Dai L M,Wang Q L,Wang S X.Relationship between tree ring chronology of Larix olgensis in Changbai Mountains and the climate change.Chinese Journal of Applied Ecology,2005,16(1):14-20.
[14] Chen L,Wu S H,Dai E F.Analysis of the tree-ring width chronologies of Pinus koraiensis and Larix olgensis on Changbai Mountains,Northeast China.Geographical Research,2011,30(6):1147-1155.
[15] Wang Z,Xu Z B.Major forest types and the community structure characteristics in the northern slope of Changbai Mountain(I)//Wang Z,ed.Forest Ecosystem Study.Shenyang:China Forestry Publishing House,1980.
[16] Holmes R L.Computer assisted quality control in tree-ring dating and measurements.Tree-ring Bulletin,1983,43:69-78.
[17] Cook E R,Holmes R L.Guide for computer program ARSTAN//Grissino-Mayer H D, Holmes R L, Fritts H C, eds.The International Tree-ring Data Bank Program Library Version 2.0 User’s Manual.Tucson:University of Arizona,1996.
[18] Kendall M G,Gibbons J D.Rank Correlation Methods.London:Edward Arnold,1990.
[19] FriedrichsD A, Neuwirth B, Winiger W, Löffler L.Methodologically induced differences in oak site classifications in a homogeneous tree-ring network.Dendrochronologia,2008,27(1):21-30.
[20] Biondi F,Waikul K.DENDROCLIM2002:a C++program for statistical calibration of climate signals in tree-ring chronologies.Computers and Geosciences,2004,30(3):303-311.
[21] Guio J.The bootstrapped response function.Tree-Ring Bulletin,1991,51:39-41.
[22] Lei J P,Xiao W F,Huang Z L,Zeng L X,Wang H Q,Li L J.Analysis of the tree-ring width chronology of Pinus massoniana in Yunyang County.Forest Research,2009,22(2):269-273.
[23] Gou X H,Chen F H,Yang M X,Peng J F,Qiang W Y,Chen K.Analysis of the tree-ring width chronology of Qilian Mountains at different elevation.Acta Ecologica Sinica,2004,24(1):172-176.
[24] Chen L, Wu S H, Pan T.Variabilityofclimate-growth relationshipsalong an elevation gradientin the Changbai Mountain,northeastern China.Trees,2011,25(6):1133-1139.
[25] Wang X M,Zhao X H,Gao L S,Jiang Q B.Age-dependent growth responses of Pinus koraiensis to climate in the north slope of Changbai Mountain,North-Eastern China.Acta Ecologica Sinica,2011,31(21):6378-6387.
参考文献:
[1] 沃尔特H.世界植被:陆地生物圈的生态系统.中国科学院植物研究所生态室,译.北京:科学出版社,1984.
[10] 王遵娅,丁一汇,何金海,虞俊.近50年来中国气候变化特征的再分析.气象学报,2004,62(2):228-236.
[11] 朱海峰.利用树轮宽度网络重建我国东北地区东部温度变化[D].北京:北京师范大学,2006.
[12] 吴祥定,邵雪梅.采用树轮宽度资料分析气候变化对树木生长量影响的尝试.地理学报,1996,51(增刊):92-101.
[13] 于大炮,王顺忠,唐立娜,代力民,王庆礼,王绍先.长白山北坡落叶松年轮年表及其与气候变化的关系.应用生态学报,2005,16(1):14-20.
[14] 陈力,吴绍洪,戴尔阜.长白山红松和落叶松树轮宽度年表特征.地理研究,2011,30(6):1147-1155.
[15] 王战,徐振邦.长白山北坡主要森林类型及其群落结构特点(之一)//王战.森林生态定位研究.沈阳:中国科学院长白山森林生态系统定位站,1980.
[22] 雷静品,肖文发,黄志霖,曾立雄,王怀情,李良俊.云阳马尾松树轮宽度年表特征研究.林业科学研究,2009,22(2):269-273.
[23] 勾晓华,陈发虎,杨梅学,彭剑锋,强维亚,陈拓.祁连山中部地区树轮宽度年表特征随海拔高度的变化.生态学报,2004,24(1):172-176.
[25] 王晓明,赵秀海,高露双,姜庆彪.长白山北坡不同年龄红松年表及其对气候的响应.生态学报,2011,31(21):6378-6387.
