不同磷水平下小麦B染色体双端体植株干物质积累和磷效率特征研究
2014-03-30郭程瑾丁长欢李小娟
郭 丽, 郭程瑾, 丁长欢, 赵 妙, 李小娟, 肖 凯*
(1 河北农业大学农学院,河北保定 071001;2 河北农业大学生命科学学院,河北保定 071001;3 华北电力大学科技学院,河北保定 071000)
提高小麦的磷素吸收和利用效率,对于促进磷素资源可持续利用和推动农业可持续发展具有重要实践意义[1-3]。研究证实,小麦等作物基因型和品种在磷素吸收和利用上存在着明显遗传多样性特征[4-5],菜豆、玉米和小麦等植物对磷胁迫的适应和抵御能力,是由多基因控制的数量遗传性状[6]。其中,调控菜豆磷效率性状的多基因间存在着积加和上位遗传效应[7]。此外,植物中众多基因参与了对磷胁迫的应答和抵御过程[8]。
对小麦养分效率与染色体关系的研究发现,小麦D染色体组含有Zn胁迫下的Zn高效利用基因[9]。不同供肥水平下,小麦二倍体染色体组AA、BB或DD、四倍体染色体组AABB和六倍体染色体组AABBDD在养分吸收、利用效率上存在明显差异,随染色体倍数增加,氮、磷和钾的利用效率显著提高。AA、BB和DD染色体组均存在着养分高效利用基因[10]。表明在小麦由二倍体向六倍体的演变和进化过程中,随产量和收获指数显著增加,养分利用效率也明显提高[11]。
B染色体组是小麦3个遗传同源群之一,在调控植株生长、发育和应答及抵御各种生物、非生物逆境中发挥着重要作用[10,12-13]。中国春(CS)及其系列遗传材料是近年来小麦遗传学和外源基因转移等研究的工具和对照材料,在重要农艺性状及抗逆性状主效基因的定位和基因克隆中具有重要理论和实践价值。前期工作中,作者研究室利用中国春遗传背景的B染色体组双端体为材料,开展了小麦光合特性染色体效应的研究[14]。本项研究针对迄今小麦有关磷效率与染色体长、短臂关系报道尚少的现状,以中国春(CS)及整套B染色体组双端体为材料,研究了丰、低磷条件下各双端体和CS的单株干重、磷效率性状、氮钾效率性状及植株形态学特征,旨在为今后小麦高效吸收和利用磷素的遗传改良提供理论依据。
1 材料和方法
1.1 试验设计

选择籽粒均匀的CS和各B染色体组代换系种子,在25℃下萌发。从萌发后种子中,选择芽长约1 cm均匀萌芽种子,摆放至漂浮于MS营养液、间隔均匀的泡沫板孔(行列间距均为10 cm)内。设置丰磷(1.2 mmol/L Pi)和低磷(12 μmol/L Pi )2种处理,其他营养含量不变。参照Sun等2011年的方法[15]进行幼苗培养,时间为3周。CS和各双端体各处理均种植12株,3次重复。培养条件为光周期12 h/12 h,温度20℃/15℃(白天/黑夜),光通量密度200 μmol/(m2·s)。处理期间,微型气泵供气,每3 d更换一次营养液。
1.2 样品测定
选择丰、低磷处理各供试材料6株,测定株高、单株叶面积、根长、单株干重、根冠比和全磷、全氮和全钾含量。其中,用直尺量取分蘖节至叶片拉直后的顶端距离获得获得株高;用叶面积分析仪测定所有叶片叶面积求和获得单株叶面积;用直尺量取各植株所有根系发根基部至根尖长度,求和后除以测定根数计算根长;将植株根系和地上部分别烘至恒重后称量求和获得单株干重;通过根系干重和地上部干重比值计算根冠比。
称取根系和地上部混合粉碎后的样本0.20 g,经浓H2SO4-H2O2消煮后,采用Auto Analyzer3(AA3 )流动注射分析仪测定样本全氮和全磷含量,采用火焰分光光度计法测定样本全钾含量[16]。通过单株干重分别与磷、氮和钾含量乘积获得单株磷、氮和钾累积量。用单株干重分别与单株磷、氮和钾累积量的比值计算磷、氮和钾效率。
1.3 数据分析
采用SAS统计学分析软件进行数据平均值、标准差、显著性测定和相关分析。
2 结果与分析
2.1 丰、低磷处理后中国春和各双端体的植株形态和单株干重
与CS相比,部分双端体的植株形态和单株干重与CS差异较小,如丰磷处理的4BS、6BS、3BL及7BL和低磷处理的4BS、6BS、7BS和7BL(图1、图2)。表明上述特定B染色体组长、短臂缺失对不同供磷下植株形态和单株干重未表现明显遗传效应。除上述双端体外,丰、低磷下其他双端体植株形态和单株干重与CS相比表现出明显差异,呈植株形态变劣(图1)和单株干重降低特征(图2)。其中,丰磷下以3BS与CS差异最大,低磷下以3BS和1BL与CS差异最大(图1、图2)。表明3B长臂和1B短臂在调控植株丰、缺磷条件下的植株干物质积累中发挥重要作用。

图1 丰、低磷处理后中国春(CS)和各双端体的植株形态Fig. 1 The plant phenotypes of Chinese spring (CS) and each ditelosomic line in the treatments with sufficient- and low-Pi[注(Note): A,丰磷 Sufficient-Pi; B,低磷 Low-Pi.]
2.2 丰、低磷处理后中国春和各双端体的植株磷累积量、磷含量和磷效率
表1表明,丰、低磷条件下,各双端体单株磷累积量与CS相比多呈减少趋势。丰磷下,单株磷累积量仅4BS较CS显著增加,而1BS、2BS、3BS、5BS、6BS、7BS和2BL较CS显著降低,其中3BS的降低幅度达到极显著水平;低磷下,单株磷累积量仅5BS较CS显著增加,而3BS、1BL和3BL较CS显著降低,其中3BS和1BL的降低幅度达到极显著水平。丰、低磷下各双端体的全磷含量与CS相比变幅相对较小,丰磷下表现为4BS和4BL较CS显著增加,5BS较CS显著降低;低磷下5BS较CS显著提高,3BL和5BL较CS显著降低。与CS相比,各双端体植株的磷效率在丰、低磷下表现较大变化,丰磷下,3BS和5BS较CS显著增加,4BS、1BL和4BL较对照显著减少;低磷下,1BL和3BL较CS显著增加,1BS、2BS和5BS较CS显著减少(表1)。表明缺失特定的B染色体长、短臂,通过改变植株在丰、低磷下的植株磷素吸收和利用能力,对植株全磷含量和干物质积累能力产生较大影响。
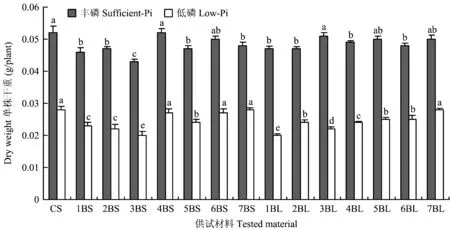
图2 丰、低磷处理后中国春(CS)和双端体的单株干重Fig. 2 The plant dry weights of Chinese spring (CS) and ditelosomic lines with sufficient- and low-Pi supplies[注(Note): 相同磷水平不同字母表示达到0.05显著水平 Different letters in the same P level mean significant difference at 0.05 level.]
注(Note): *和**表示与CS相比分别达到0.05和0.01显著水平 Mean significant difference compared with CS at 0.05 and 0.01 levels, respectively.
2.3 丰、低磷处理后中国春和各双端体的植株氮钾累积量、氮钾含量和氮钾效率
表2表明,丰、低磷条件下,各双端体的植株氮累积量与CS相比差异较小,均未达到统计学显著水平。丰磷下各双端体全氮含量与CS的差异也均不显著;低磷下,7BS、1BL和2BL的全氮含量较CS显著降低,其他双端体全氮含量与CS差异不显著。丰磷下,2BS、5BS、6BS和7BS氮效率较CS显著降低;低磷下,3BS氮效率较CS显著降低(表2)。表明缺失部分B染色体长短臂对低磷下植株全氮含量(7BS、1BL和2BL)、氮效率(3BS)和丰磷下氮效率(2BS、5BS、6BS和7BS)也具有明显调控效应。
表3显示,丰、缺磷条件下,各双端体全钾含量和钾效率均与CS无显著差异,单株钾累积量也与CS差异较小。其中,仅3BS在上述供磷处理下的单株钾累积量较CS显著降低。表明供磷水平对双端体钾素吸收和利用的调控效应较小,与不同供磷下各双端体与CS在磷氮含量、单株磷氮累积量和磷氮效率上多表现明显差异的结果不同。3BS在丰、低磷下单株钾累积量均较CS显著降低,表明该染色体短臂含有钾高效吸收调控基因。

表2 不同供磷水平下各双端体的植株氮累积量、全氮含量和氮效率Table 2 The N accumulation amount, total N contents and N usage efficiencies in Chinese spring (CS) in plant and ditelosomic lines with sufficient- and low-Pi supplies
注(Note): *和**表示与CS相比分别达到0.05和0.01显著水平 Mean significant difference compared with CS at 0.05 and 0.01 levels, respectively.
2.4 丰、低磷处理后中国春和各双端体的植株形态学性状
CS和各双端体植株株高、单株叶面积、平均根系长度和根冠比见表4。与CS相比,各双端体丰、低磷条件下的株高、叶面积、根长和根冠比均具有不同程度改变。其中,丰、低磷下各双端体根长和根冠比与CS相比的差异均未达到显著水平。丰、低磷下,3BS单株叶面积均较CS显著减小,与丰低磷下该双端体较CS植株形态明显变劣和单株干重显著降低相吻合。除3BS外,其他双端体单株叶面积与CS相比均无显著差异。丰磷下,1BS和3BS株高较CS显著减小,其他双端体株高与CS差异不显著;低磷下,3BL和4BL株高较CS显著降低,其他双端体株高与CS无显著差异。
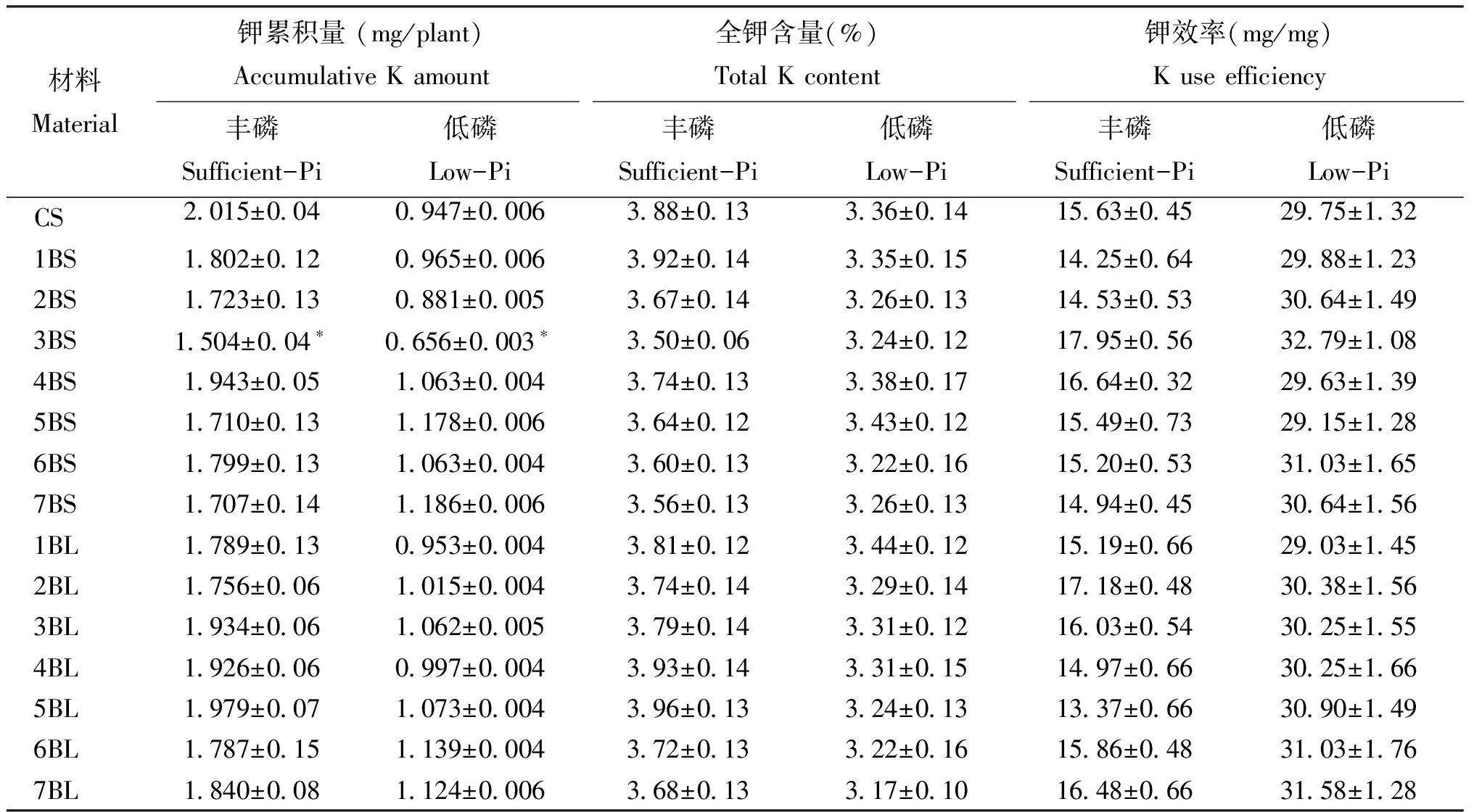
表3 不同供磷水平下各双端体的植株钾累积量、全钾含量和钾效率Table 3 The plant accumulation amount of K, total K contents and K usage efficiencies in Chinese spring (CS) and ditelosmic lines with sufficient- and low-Pi supplies
注(Note): *和**表示与CS相比分别达到0.05和0.01显著水平 Mean significant difference compared with CS at 0.05 and 0.01 levels, respectively.

表4 不同供磷水平下各双端体的植株株高、叶面积、根长和根冠比Table 4 The plant heights, leaf area per plant, root lengths and ratios of root weight and aerial weight of Chinese spring (CS) and ditelosmic lines with sufficient- and low-Pi supplies
注(Note): *和**表示与CS相比分别达到0.05和0.01显著水平 Mean significant difference compared with CS at 0.05 and 0.01 levels, respectively.
2.5 丰、低磷下单株干重、磷效率性状与氮钾效率性状和植株形态学性状的相关分析
丰磷下,单株干重与磷累积量、全磷含量、氮效率、钾累积量、钾效率、株高呈显著正相关,与其他测试性状和指标相关不显著;磷效率与磷累积量和全磷含量分别呈显著和极显著负相关。低磷下,单株干重与磷累积量、氮累积量、钾累积量、钾效率和叶面积;磷效率与磷累积量和全磷含量呈极显著负相关,与根长呈显著正相关(表5)。上述结果表明,不同供磷水平下的植株干重和磷效率与部分养分含量、效率参数和植株形态学性状具有紧密联系。
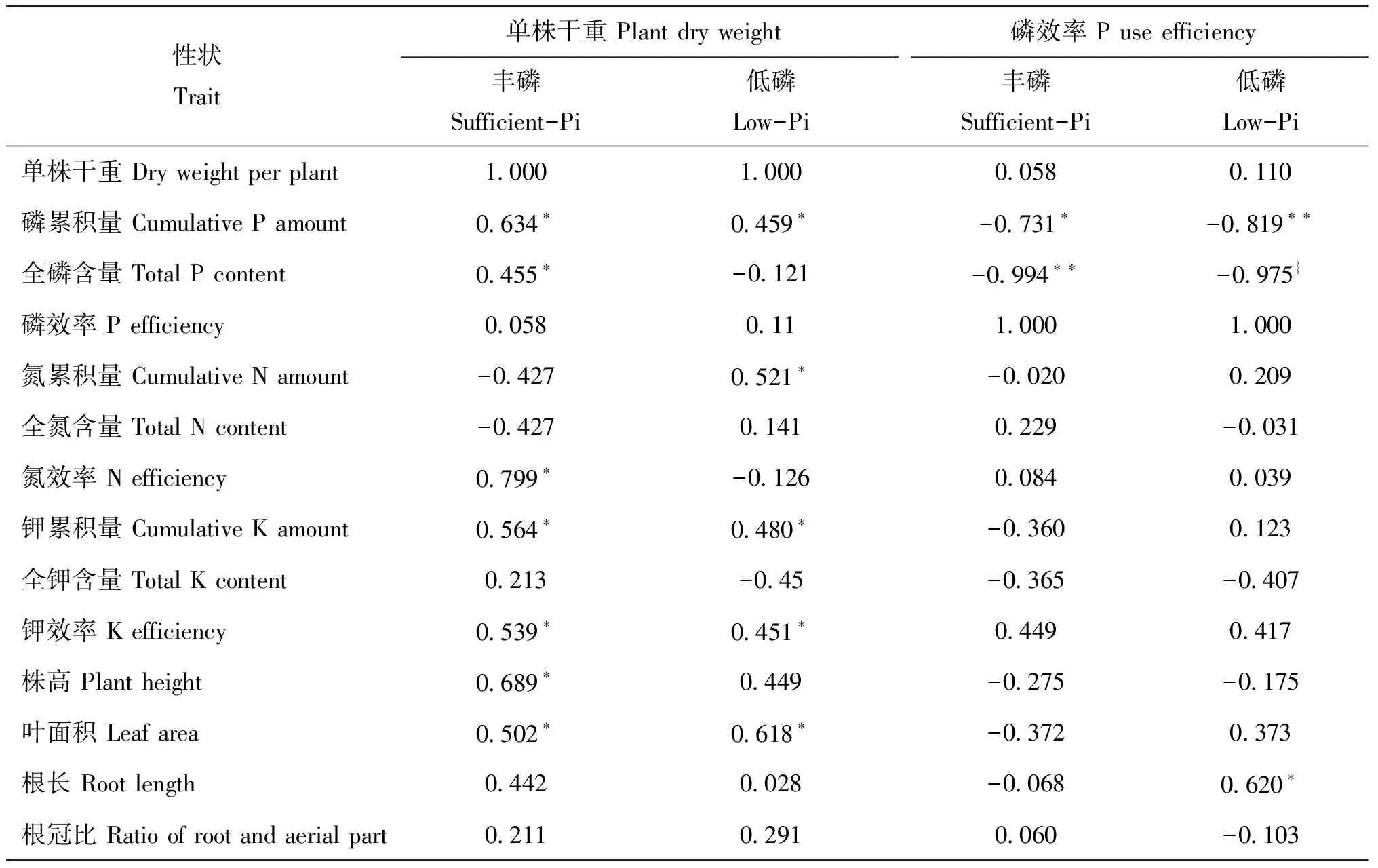
表5 单株干重、磷效率与氮钾养份效率性状和植株形态学性状间的相关系数Table 5 The correlation coefficients between plant dry weight and P use efficiency with N, and K efficiency and some plant phenotype parameters under sufficient- and low-Pi conditions
注(Note): *和**表示相关程度分别达到0.05和0.01显著水平 Mean significant difference compared with CS at 0.05 and 0.01 levels, respectively.
此外,丰磷下,单株干重与氮效率、钾累积量、钾效率、株高和叶面积均呈显著正相关,磷累积量与氮累积量、钾累积量钾效率和单株叶面积均呈正相关,全磷含量与氮效率呈正相关;低磷下,植株干重与氮累积量、钾累积量、钾效率和叶面积呈正相关,磷累积量与株高和叶面积呈正相关、与根长呈负相关;全磷含量和磷效率均与根长呈负相关(表5)。表明CS和双端体植株磷素吸收和利用能力,与部分氮、钾效率相关参数和植株形态学性状共同作用,影响着不同磷水平下植株的干物质生产能力。单株叶面积与植株干重和单株磷累积量均呈显著正相关,表明前者可作为评价不同磷水平下供试小麦双端体及其他遗传学材料磷效率的形态学指标。
3 讨论与结论
小麦对矿质元素吸收、利用及效率特征在很大程度上受到遗传因子的调控[6-8,17-18]。六倍体栽培小麦中的B染色体组源于拟斯卑尔脱山羊草,在小麦的生长发育、产量和品质形成及植株的抗逆性中具有重要的作用[12]。研究表明,与A、D染色体组相比,小麦B染色体组在调控CO2同化速率和增加叶绿素的含量上具有更重要的遗传调控效应[12]。其中,2B 染色体对对光合速率和叶绿素含量具有正向调控效应,1B 和3B 染色体对光合速率和叶绿素含量具有负向调控效应[13]。
李小娟等(2006)以小麦B染色体组双端体为材料,研究了B染色体组长短臂对光合效率的影响。发现染色体臂1BL和3BS对叶绿素含量分别具有正、负效应;2BL和7BL对光反应活性分别具有正、负效应,表明1BL和2BL含有高光效主效基因[14]。本研究对丰、低磷不同磷水平下中国春及其遗传背景B染色体组整套双端体的植株干重和磷效率相关性状研究表明, 3BS含有调控丰磷下植株干物质积累主效基因,3BS和1BL含有调控低磷逆境下植株干物质积累主效基因。此外,在植株磷累积量上,丰磷条件下,1BS、2BS、3BS、5BS、6BS、7BS和2BL较CS显著降低,其中3BS的降低幅度达到极显著水平;低磷条件下,3BS、1BL和3BL较CS显著降低,其中3BS和1BL的降低幅度达到极显著水平。表明上述B染色体组长、短臂上含有调控小麦磷素吸收和累积的主效基因。上述部分结果与李玉京等[19]利用以中国春-长穗偃麦草二体异附加系和二体异代换系为材料,证实小麦3B和5B染色体携带磷高效基因的结果相吻合。有关不同磷水平下植株干物质积累和磷效率性状的相关生化和分子机制及其内在联系有待进一步探讨。
本研究发现,丰、缺磷不同供磷条件下,与中国春相比,部分供试B染色体组双端体氮、钾效率性状也发生不同程度改变。表明部分B染色体组长、短臂在调控不同磷水平下植株磷效率的同时,对氮、钾吸收和利用效率也产生较大调控效应。有关不同氮、钾供应条件下中国春(CS)和各B染色体组双端体的氮素吸收、利用和干物质积累特征有待进一步研究。
不同供磷水平下,小麦植株在根系形态、地上部株高及叶面积、生理生化特征上均发生明显变化[20,21],在调控植株的磷素吸收、利用能力及磷效率上发挥重要作用[5,22]。本研究对不同供磷水平下中国春及供试B染色体组双端体植株干重和单株累积量的相关分析发现,上述两个性状之间在丰、低磷条件下均呈显著正相关,表明植株干物质累积数量与植株根系吸收的磷素数量之间具有密切联系。较多的磷素吸收,是部分双端体低磷逆境下缓解磷胁迫压力、改善植株体内的能量合成和生化代谢,进而增强植株光合物质积累的重要原因。这与丰、低磷条件下单株干重和单株磷累积量均与单株叶面积呈显著正相关的结果相吻合。表明植株磷素吸收数量增多,通过调控的植株光合面积,在较大程度上影响着植株的干物质累积量。因此,可用单株面积作为小麦染色体双端体及特定小麦染色体分析遗传材料的磷效率评价指标。
综上,本研究表明,与中国春相比,丰磷下4BS、6BS、3BL及7BL和低磷下4BS、6BS、7BS和7BL植株形态和单株干重无明显变化,其他B染色体组双端体植株形态变劣、单株干重降低。丰、低磷下,各双端体单株磷累积量与CS相比多呈减少趋势;磷效率大体呈与单株磷累积量相反趋势特征。丰、低磷条件下,单株干重与磷累积量及磷累积量与全磷含量显著相关,单株干重和磷累积量均与单株叶面积显著相关,因此,可用单株叶面积作为丰、低磷下植株磷效率的形态学评价指标。
参考文献:
[1] Wissuwa M. How do plants achieve tolerance to phosphorus deficiency? Small causes with big effects[J]. Plant Physiol., 2003, 133(4): 1947-1958.
[2] Vance C P, Uhde S C, Allan D L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource[J]. New Phytol., 2003, 157: 423-447.
[3] Bieleski R L. Phosphate pools, phosphate transport, and phosphate availability[J]. Annu. Rev. Plant Physiol., 2007, 24: 225-252.
[4] 严小龙, 张福锁. 植物营养遗传学[M]. 北京: 中国农业大学出版社, 1997, 1-30.
Yan X L, Zhang F S. Plant nutrition and genetics[M]. Beijing: Chinese Agricultural University Press, 1997, 1-30
[5] 郭程瑾, 李宾兴, 周彦珍, 等. 不同磷效率小麦品种的磷吸收特性[J]. 植物遗传资源学报, 2006, 7(1): 49-53.
Guo C J, Li B X, Zhou Y Zetal. Characteristics of phosphorus acquisition in wheat varieties with different P efficiencies[J]. J Plant Gen. Resourc., 2006, 7(1): 49-53.
[6] 严小龙, 卢永根. 普通菜豆的起源、进化和遗传资源[J]. 华南农业大学学报, 1994, 15(4): 111-116.
Yan X L, Lu Y G. The origin, evolution and genetic resources of kidney bean[J]. J. South China Agric. Univ.,1994,15(4): 111-116.
[7] 严小龙. 全国植物营养与肥料学术年会论文集[M]. 北京: 中国农业科技出版社, 1994. 15-18.
Yan X L. Annual proceedings of plant nutrition and fertilizer[M]. Beijing: Chinese Agricultural Science and Technology Press, 1994. 15-18.
[8] Misson J, Raghothama K G, Jain Aetal. A genome-wide transcriptional analysis usingArabidopsisthalianaAffymetrix gene chips determined plant responses to phosphate deprivation[J]. Proc. Natl. Acad. Sci. USA, 2005, 102: 11934-11939.
[9] Cakmak I, Torun B, Erenoglu Betal. Morphological and physiological differences in cereals in response to zinc deficiency[J]. Euphytica, 1998, 100: 349-357.
[10] Gill B S, Friebe B, Endo T R. Standard karyotype and nomenclature system for the description of chromosome bands and structural aberrations in wheat (Triticumaestivum)[J]. Genome, 1991, 34: 830-839.
[11] Batten G D. The uptake and use of phosphorus and nitrogen by diploid, tetraploid and hexaploid wheats (Triticumspp. )[J]. Ann Bot., 2006, 58: 49-59.
[12] Planchon C, Fesquet J. Effect of the D genome and of selection on photosynthesis in wheat[J]. Theor. Appl. Genet., 1982, 61: 359-365.
[13] Watanabe N, Ogawa A, Kitaya Tetal. Effects of substituted D genome chromosomes on photosynthetic rate of durum wheat (TriticumturgidumL. var. durum)[J]. Euphytica, 1994, 72(1/2): 127-131.
[14] 李小娟, 蓝岚, 李雁鸣, 等. 小麦B染色体组双端体旗叶光合能量转换参数的研究[J]. 华北农学报, 2006, 21(5): 9-12.
Li X J, Lan L, Li Y Metal. Studies on photosynthetic parameters related to energy transformation of flag leaf in wheat B ditelosomic lines[J]. Acta Agric. Boreali-Sin., 2006, 21(5): 9-12.
[15] Sun Z H, Ding C H, Li X Jetal. Molecular characterization and expression analysis ofTaZFP15, a C2H2- type zinc finger transcription factor gene in wheat (TriticumaestivumL.)[J]. J. Integr. Agric., 2012, 11: 31-42.
[16] 张英利, 许安民, 尚浩博, 等. AA3型连续流动分析仪测定土壤和植物全氮的方法研究[J]. 西北农林科技大学学报, 2006, 34(10): 128-132.
Zhang Y L, Xu A M, Shang H Betal. Studies on the assay method of total nitrogen in soil and plants using AA3 continuous flow analyzer[J]. J. Northwest A&F Univ., 2006, 34(10): 128-132.
[17] 郭程瑾, 张立军, 崔喜荣, 等. 氮胁迫条件下中国春-代换系小麦苗期耐低氮特征研究[J]. 植物营养与肥料学报,2011, 17(1): 29-37.
Guo C J, Zhang L J, Cui X Retal. Characterization of low-nitrogen tolerance capacities of Chinese Spring substitution lines of wheat under deficient-N condition at the seedling stage[J]. Plant Nutr. Fert. Sci., 2011, 17(1): 29-37.
[18] Guo C J, Li J C, Chang W Setal. Effects of chromosome substitution on the utilization efficiency of nitrogen, phosphorus, and potassium in wheat[J]. Front. Agric. China, 2011, 5(3): 253-261.
[19] 李玉京, 刘建中, 李滨, 等. 普通小麦各基因组有效利用土壤磷基因的遗传分析[J]. 西北植物学报, 1999, 19(1): 1-6.
Li Y J, Liu J Z, Li Betal. Genetic analysis of each wheat genome on effectively using the soil available phosphorus[J]. Acta Bot. Boreali-Occid. Sin., 1999, 19(1): 1-6.
[20] 郭丽, 龙素霞, 赵芳华, 等. 小麦不同品种磷效率比较和评价的生化指标研究[J]. 植物遗传资源学报, 2008, 9(4): 506-510.
Guo L, Long S X, Zhao F Hetal. Comparison and evaluation of biochemical criteria for phosphorus efficiency in wheat[J]. J. Plant Gen. Resourc., 2008, 9(4): 506-510.
[21] 郭程瑾, 李宾兴, 王斌, 等. 小麦高效吸收和利用磷素的生理机制[J]. 作物学报, 2006, 32(6): 827-832.
Guo C J, Li B X, Wang Betal. Physiological mechanisms of absorption and use of phosphorus with high efficiency in wheat cultivars[J]. Acta Agron. Sin., 2006, 32(6): 827-832.
[22] 张建恒, 李宾兴, 王斌, 等. 不同磷效率小麦品种光合碳同化和物质生产特性研究[J]. 中国农业科学, 2006, 39(11): 2200-2207.
Zhang J H, Li B X, Wang Betal. Studies on the characteristics of photosynthesis and dry matter production in wheat varieties with different P efficiency[J]. Sci. Agric. Sin., 2006, 39(11): 2200-2207.
