Pb2+、Cr6+、Hg2+和Cu2+对碱蓬幼苗生长及生理生化指标的影响
2014-03-29王思瑶
王思瑶,姜 华
(辽宁师范大学 生命科学学院,辽宁 大连 116029)
目前,植物修复技术已被国内外广泛应用,尤其是生长周期短、生物量累积迅速的草本植物在植物修复中显示出较大的研究价值,很多文献也证明了其作为重金属污染土壤修复的潜在能力[1-2],并且筛选出来了一些重金属超积累植物[3-4]。碱蓬是藜科、碱蓬属的一年生草本植物,多生于海滨、荒漠、湖边及盐碱土等非耕地上,营养丰富,富含脂肪、蛋白、粗纤维、矿物质、微量元素和多种维生素等,可作为蔬菜食用。碱蓬籽油富含人体生长发育所必需的脂肪酸、亚油酸和亚麻酸,具有抗肿瘤、抗动脉硬化、抗氧化和降低体内脂肪等医药保健功效;碱蓬还具有耐盐碱、改善土壤生态等功效,被誉为盐碱地改造的先锋植物[5]。前人关于碱蓬做了大量研究,如品种选育与栽培、群落构成、营养成分分析、抗性生理、生态功能、遗传多样性及耐盐基因的利用等[6],但有关碱蓬对重金属的耐、抗性及污染的修复研究报道有限。本文旨为重金属污染的碱蓬修复提供理论依据。
1 材料与方法
1.1 材料
(1)碱蓬:采自辽宁省大连市甘井子区盐场海滨滩涂上的碱蓬(Suaedasalsa)。
(2)器具:722型光栅分光光度计(上海成光仪器厂);紫外分光光度计(Toshiba);TGLL-18K台式高速冷冻离心机(太仓市华美生化仪器厂);电热恒温干燥箱(上海跃进医疗器械厂);恒温水浴。
(3)药品:五水合硫酸铜、硝酸铅、重铬酸钾、氯化汞、氯化钾、磷酸氢二钠、磷酸二氢钠、三氯乙酸、聚乙烯吡咯烷酮、儿茶酚、硼酸、硫代巴比妥酸、碳酸钙粉、愈创木酚、过氧化氢、95%乙醇、牛血清蛋白、考马斯亮蓝G250等均为分析纯。
1.2 方法
(1)重金属浓度的确定:先配制CuSO4·5H2O、Pb(NO3)2、K2Cr2O7和HgCl2母液,再配制成不同浓度。将碱蓬苗放于不同梯度的浓度下培养,观察其生长状况,确定4种重金属对碱蓬植株生长出现明显抑制效果的临界浓度,预实验得知,4种重金属Pb2+、Cr6+、Hg2+和Cu2+的临界浓度依大小排序为45 mg/kg、15 mg/kg、10 mg/kg和5 mg/kg。以此为工作浓度。
(2)碱蓬苗的采集与培育:采集海滨滩涂上自然生长的碱蓬小苗(6~8叶期),洗去根部土壤,培养于装有4种重金属溶液的三角瓶中。为保证重金属浓度,每隔4天更换1次,3次重复,以纯水为对照,培养20天后开始测定。
(3)生长与生化指标的测定:取出小苗,用滤纸吸干水分。分别测定植株的根长和根数、植株鲜重,计算耐性指数(处理组根系平均长度×100/对照组根系平均长度);连续6天采样(叶片),采用分光光度比色法测定叶绿素[7]、可溶性蛋白[8]和丙二醛含量[9],测定过氧化物酶(POD)和过氧化氢酶(CAT)活性[7]。
2 结果与分析
2.1 重金属胁迫对碱蓬生长指标的影响
试验结果表明(见表1):Pb2+、Cr6+、Hg2+和Cu2+对碱蓬植株生长出现明显抑制效果的临界浓度依其大小排序为45 mg/kg、15 mg/kg、10 mg/kg和5 mg/kg,以此浓度水培碱蓬幼苗20天后采样测量,处理组的各项生长指标明显低于对照组。其中,Pb2+处理的植株株高、根数及鲜重分别是对照组的89%、83%和73%;Cr6+处理的分别是对照组的87%、75%和73%,Hg2+处理的分别是对照组的87%、66%和70%,Cu2+处理的分别是对照组的89%、88%和75%。
依据根系长度,计算碱蓬对Pb2+、Cr6+、Hg2+和Cu2+的耐性指数分别为83.18、78.62、80.49和83.71,从中可见,碱蓬对Pb2+耐性较强。

表1 重金属胁迫对碱蓬生长指标的影响
2.2 重金属胁迫对碱蓬叶绿素含量的影响
由表2可知,4种重金属胁迫下的碱蓬叶绿素a、叶绿素b及叶绿素总量均低于对照组,且处理与对照比较叶绿素总量差异显著(P<0.05)。Pb2+处理的叶绿素a、叶绿素b及叶绿素总量分别是对照的65%、52%和61%;Cr6+处理的是对照的66%、42%和59%;Hg2+处理的分别是对照的65%、34%和55%;Cu2+处理的分别是对照的68%、64%和67%。

表2 重金属胁迫对碱蓬光和色素含量的影响
2.3 重金属胁迫对碱蓬可溶性蛋白和丙二醛含量的影响
由图1可知,4种重金属胁迫下碱蓬植株的可溶性蛋白含量均低于对照,而MDA含量均高于对照。Pb2+、Cr6+、Hg2+和Cu2+处理的可溶性蛋白含量分别是对照的66.7%、58.3%、41.7%和75%;Hg2+处理与对照比差异显著(P<0.05);而Pb2+、Cr6+、Hg2+、Cu2+处理的丙二醛含量分别是对照的167%、147%、114%、和131%,Pb2+处理与对照比差异显著(P<0.05)。
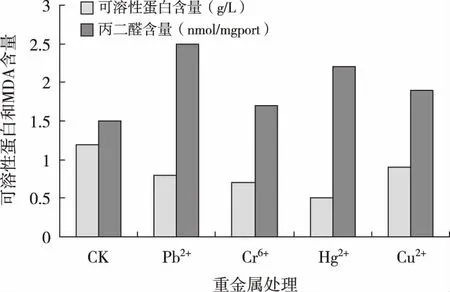
图1 重金属胁迫对碱蓬可溶性蛋白和丙二醛含量的影响
2.4 重金属胁迫对碱蓬过氧化物酶(POD)和过氧化氢酶(CAT)活性的影响
重金属处理20天后,连续6天采样测定POD和CAT活性。结果得知,4种重金属胁迫下的碱蓬植株体内POD活性均高于对照,而CAT活性均低于对照(图2和图3)。Pb2+、Cr6+、Hg2+、Cu2+处理植株的POD酶活性6天总和分别是对照的116%、165%、135%和153%,而CAT活性分别是对照的73.8%、66.7%、74%和61.5%。

图2 重金属胁迫对碱蓬POD活性的影响
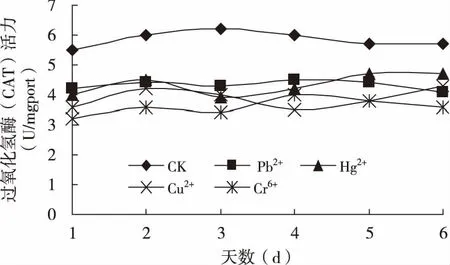
图3 重金属胁迫对碱蓬CAT活性的影响
3 结论与讨论
3.1 结论
重金属Pb2+、Cr6+、Hg2+和Cu2+胁迫对碱蓬植株生长均有抑制作用,出现明显抑制效果的临界浓度分别为:45 mg/kg、15 mg/kg、10 mg/kg和5 mg/kg;以植株株高、根数及鲜重排序,Pb2+处理的植株株高、根数及鲜重分别是对照组的89%、83%和73%,Cr6+处理的是87%、75%和73%,Hg2+处理的是87%、66%和70%,Cu2+处理的是89%、88%和75%;4种重金属胁迫下碱蓬植株的叶绿素、可溶性蛋白含量均低于对照,而MDA含量均高于对照。Pb2+、Cr6+、Hg2+、Cu2+处理的叶绿素总量分别是对照的61%、59%、55%和67%,可溶性蛋白含量分别是对照的66.7%、58.3%、41.7%和75%,而丙二醛含量分别是对照的167%、147%、114%、和131%。4种重金属胁迫下的碱蓬植株体内POD活性均高于对照,而CAT活性均低于对照。
3.2 讨论
本文查明了使碱蓬幼苗生长出现明显抑制效果的4种重金属(Pb2+、Cr6+、Hg2+和Cu2+)的临界浓度,以及此浓度胁迫下的生长及生理生化指标的变化。可溶性蛋白含量是一个重要的生理生化指标,是构成酶的重要组成部分,参与生理生化代谢过程的调控,与植物的生长发育、抗逆性等密切相关。光合色素是植物光合作用的物质基础,其含量高低是判断植物光合作用强弱的一个重要指标[10]。本实验结果表明,4种重金属的胁迫均可使碱蓬蛋白质和叶绿素合成受阻,在临界浓度时蛋白含量降低近25%~60%、叶绿素总量降低33%~45%。尤其是Hg2+可使蛋白含量降低近60%、叶绿素近45%。重金属可能与叶绿素合成的相关酶的肽链部分结合,抑制了酶活性、阻碍了叶绿素的合成[11]。植物组织在遭受胁迫时,往往发生膜脂过氧化作用,产生多种多样的产物,丙二醛是主产物之一,其数量的多少能代表膜脂过氧化程度,也可间接反应植物抗氧化能力的强弱[12]。逆境胁迫下,丙二醛含量往往会增加,并随胁迫作用的加大含量升高[13]。本实验结果表明,在4种重金属胁迫下,MDA含量与对照相比升高31%~67%,尤其是Pb2+处理的使之升高近67%。
植物根系是最先接触重金属的部位,对其进行吸收或排斥,其耐性指数是用来反映植物对重金属耐性大小的一个重要指标[14]。重金属影响植物根尖细胞有丝分裂,造成细胞分裂速度减慢,并通过改变植物的生理生化过程而影响其生长发育[15]。本实验结果表明,4种重金属均可使碱蓬植株的鲜重减轻、根数减少。在临界浓度时,碱蓬植株鲜重降低25%~30%、根数减少12%~34%,叶片长减少17%~32%,叶片数降低11%~42%尤其是Hg2+可使碱蓬植株鲜重降低30%、根数减少34%,叶片长缩短32%,叶片数减少42%。
POD是植物体内的一类氧化酶,对环境变化反应灵敏,植物在受到逆境胁迫时会诱导产生,在植物呼吸代谢中具有重要作用。前人研究表明,植物在受到污染或逆境胁迫时,POD酶活性有增加趋势[16]。CAT普遍存在于植物组织中,能保护植物免受过氧化氢积累的危害,其活性的高低亦可反映植物抗性的强弱,酶活性降低是植物对逆境的一种适应性表现[17]。许多研究发现植物在受到逆境或污染胁迫时,CAT活力随逆境或污染胁迫强度增加会出现不同程度的下降[18]。本实验4种重金属胁迫下的碱蓬植株体内POD酶活性升高、而CAT酶活性降低,这与前人的研究结果一致。
参考文献:
[1] Alkorta I, Garbisu C.Phytoremediation of organic contaminants[J].Bioresource Technology,2001,79:273-276.
[2] Baker A J M, McGrath S P, Sidoli C M D, et al.The possibility of in situ heavy metal decontamination of polluted soils using crops of metal-accumulating plants[J].Resource Conservation and Recycling,1994,11:41-49.
[3] Chaney R L, Malik M, Li Y M, et al.Phytoremediation of soil metals[J].Current Opinion Biotechnology,1997,8:279-284.
[4] Raskin I,Smith RD,Salt DE.Phytoremediation of metals: usingplants to remove pollutants from the environment[J].Current Opinion Biotechnology,1997,8:221-226.
[5] 任伟重,姜华,郑音,等.碱蓬资源的开发价值[J].辽宁农业科学,2011(5):51-53.
[6] 张洛艳.沿海地区盐地碱蓬的分子标记研究[D].山东师范大学硕士学位论文,2010:5.
[7] 王学奎.植物生理生化实验原理和技术[M].2版.北京:高等教育出版社,2006.
[8] 李玲.植物生理学模块实验指导[M].北京:科学出版社,2009.
[9] 郝再彬,苍晶,徐仲,等.植物生理实验[M].哈尔滨:哈尔滨工业大学出版社,2004.
[10] 杜天庆,杨锦忠,郝建平,等.菰和菖蒲对重金属的胁迫反应及其富集能力[J].生态学报,2007,27(1)281-287.
[11] Somashekaraiah B V, Padamajaes K, Prasad R K.Phytotoxicity of cadmium ions on germination seedlings of mung bean(Phaseolus vulgartze); Involvement of liquid peroxides in chlorophyll degradation[J].Plant Physiology,1992(65):85-89.
[12] Clijsters H,Van Assche F.Inhibtion of photosynthesis by heavy metals[J].Photosynthesis Research,1985(7):31-40.
[13] 陈明林,王友保.水分胁迫下外来种铜锤草和本地种酢浆草的生理指标比较研究[J].草业科学,2008,17(6):52-59.
[14] 严明理,刘丽莉,王海华,等.3种植物对红壤中镉的富集特性研究[J].农业环境科学学报,2009,29(8):4475-4482.
[15] Seregin I V, Kozhevnikova A D.Physiological role of nickel and its toxic effects on higher plants[J].Russian Journal of Plant Physiology,2006,53(2):257-277.
[16] 郑福丽,江丽华,刘兆辉,等.石油污染物对油菜产量及生理指标的影响[J].中国农学通报,2009,25(19):279-282.
[17] 刘祖祺,张石诚.植物抗性生理学[M].北京:中国农业出版社,1994.
[18] 王雪峰,陈桂珠,徐夏玲.白骨壤对石油污染的生理生态响应[J].生态学报,2005,25(5):1095-1100.
