苏姜猪及其杂交后代HSL基因PCR-RFLP分析
2014-03-26任善茂徐椿慧董晓君赵旭庭
任善茂, 陶 勇, 徐椿慧, 庄 勋, 董晓君, 赵旭庭
(1. 江苏农牧科技职业学院, 江苏 泰州 225300; 2. 江苏省淮安市淮安区畜牧兽医站, 江苏 淮安 223200)
猪激素敏感脂肪酶(HSL)基因是影响脂肪代谢的重要候选基因之一,基因全长10~11 kb,有9个外显子,编码171个氨基酸[1],定位于6q1.2[2]。猪HSL基因在不同组织中的表达存在较大差异性,在心肌和骨骼肌中HSL基因mRNA含量极少[3]。Otsu等认为HSL基因所在的染色体区域可以作为影响猪瘦肉率、肌内脂肪等屠宰性状的重要区段[4],曲亮等将HSL基因作为影响猪胴体重要性状的候选基因进行相关研究[5]。因此,本试验利用PCR-RFLP方法研究了苏姜猪及其杂交后代群体内HSL基因外显子I的多态性,为进一步分析HSL基因与猪背膘厚、瘦肉率等胴体性能的关系奠定分子生物学基础。
1 材料与方法
1.1 材料
试验选用江苏姜曲海种猪提供的苏姜猪、长苏猪(长白猪×苏姜猪)、大苏猪(大白猪×苏姜猪)各60头,90 kg左右。每头猪采集10 mL血液,ACD抗凝。采用常规的酚/氯仿/异戊醇法抽提血液DNA,乙醇沉淀,溶于TE缓冲液。使用BD-100超微量核酸蛋白分析仪测定浓度后,稀释至50 ng/μL分装,-20℃保存备用。
1.2 主要设备与试剂
ABI梯度PCR扩增仪(美国),DYY-6C恒压恒流电泳仪(北京六一),垂直板电泳槽装置(北京东方),Eppendorf冷冻离心机(USA),GDS-8000型凝胶成像系统(美国UVP公司)。
TaqDNA聚合酶、dNTP购自上海华美生物工程公司,DNA Marker、BsaH I购自上海生工生物工程技术服务有限公司,丙烯酰胺(Acrylamide,Acr)、N-N′亚甲基双丙烯酰胺(Bisacrylamide,Bis)购自南京基天生物技术有限责任公司。
1.3 引物设计与合成
根据猪HSL基因序列(Genbank登录号:AJ000483),参照文献[6]设计一对引物,序列为:上游引物(PF)5′-CGCACAA TGACACA BT CA CT GGT-3′,下游引物(PR)5′-AGGCAGCGGCCGT AGAAGCA-3′。扩增片段长度为497bp,位于HSL基因外显子I区域。引物由上海生工生物工程有限公司合成,去离子水溶解,-20℃保存。
1.4 PCR-RFLP分析
PCR 反应体系20 μL:10 × buffer 2 μL,dNTPs 1.5 μL,Taq DNA 聚合酶0.1 μL( 2U/μL),上下游引物各0.5 μL,基因组DNA 1 μL,ddH2O补足至20 μL。
PCR 反应条件: 94℃预变性10 min; 94℃变性30 s, 59.5℃退火30 s, 72℃ 延伸30 s, 32个循环; 72℃延伸5 min; 4℃保存。扩增产物以2% 琼脂糖凝胶电泳检测。
酶切体系15 μL:10 × buffer 2 μL,BsaH I内切酶3U,PCR 产物10 μL,ddH2O补足至15 μL, 37 ℃酶切3 h。酶切产物经8% 聚丙烯酰胺电泳检测,电压6 V/cm,电泳8 h,银染显带、照相、记录并保存。
2 结果与分析
2.1 PCR-RFLP结果
PCR扩增结果见图1,没有发现非特异性条带,PCR扩增产物大小约500 bp,产物长度与预测结果一致,可以用于进行RFLP分析。
PCR-RFLP结果见图2所示,PCR扩增产物经BsaH I限制性内切酶消化后,表现出了多态性。根据条带数和位置,判定有3种基因型,分别为AA(190 bp、307 bp)、AB(67 bp、190 bp、240 bp和307 bp)和BB(67 bp、190 bp和240 bp)基因型。其中,因片断67 bp较小,电泳时间较长而发生弥散消失。
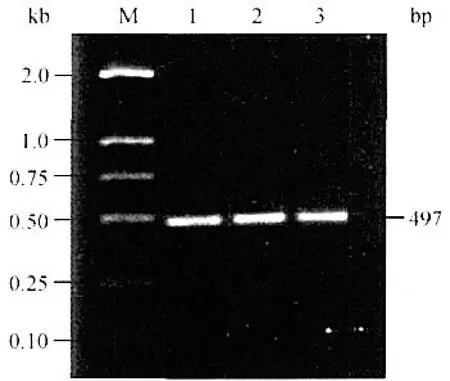
M:DL2000 DNA marker;1-3:PCR products
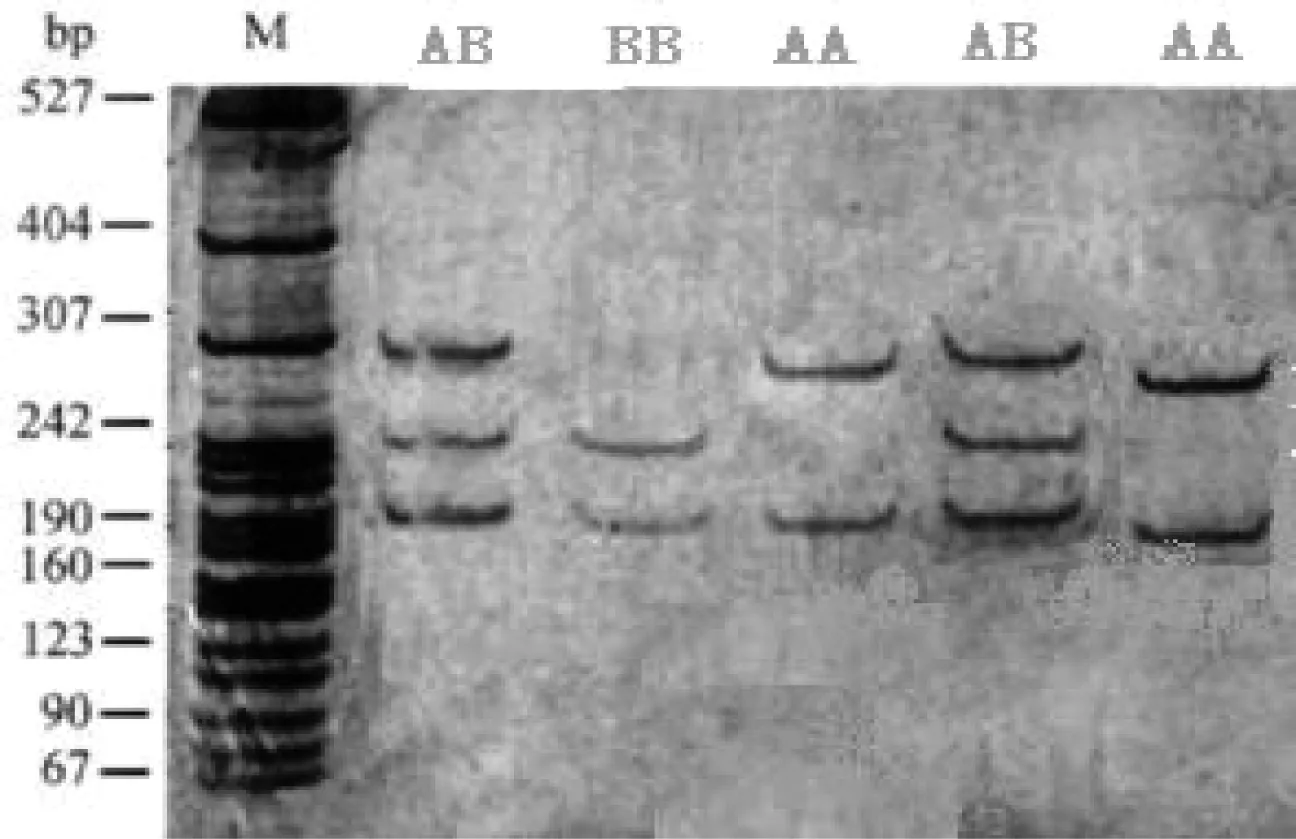
M:pBR322/MspⅠ marker;1,5:AB型;2:AA型;3,4:BB型
2.2 HSL基因外显子I基因型和等位基因频率
苏姜猪及其杂交后代不同基因型检出数和等位基因频度见表1。由表1可见,3种基因型在苏姜猪及其杂交后代中均检测出,苏姜猪有更多的等位基因A。

表1 猪HSL基因的基因型及等位基因频率
注:括号内为样本数。
3 讨论
猪HSL基因为调控猪体内激素敏感脂肪酶的基因,外显子I区域内存在G→A的碱基转换,改变了BsaH I酶识别位点,最终导致编码氨基酸Val替换Ile,其不同等位基因在中外猪种群体中的分布差异很大[6]。强巴央宗等[7]在藏猪群体中检测到HSL基因外显子ⅠG→A突变位点,出现3种基因型,表现出多态性,等位基因A和G频率分别为62.50%和37.50%,突变位点的基因分布均处于瘦肉型外种猪和脂肪性地方猪之间。陈利荣等[8]研究发现瘦肉型大白猪、长白猪全部表现为GG基因型,山西黑猪表现为AA、AG和GG 3种基因型,华北杂种野猪、东北纯种野猪及杂种野猪表现为AG和GG 2种基因型,研究结果丰富了国内外对野猪的分子生物学研究,为野猪遗传资源的合理开发利用提供了依据。
本试验中,在苏姜猪及其与长白猪、大白猪的杂交后代群体中均检测到了HSL基因外显子I区域内G→A碱基突变产生的3种基因型,基因型频率分布介于瘦肉型外种猪和脂肪型地方猪种之间,这与HSL的功能特性及苏姜猪的品种来源有一定的关系。HSL主要是将白脂肪中的甘油三酯分解成游离脂肪酸和甘油[9],因此可以推测,HSL基因外显子I的A→G转换而导致的Ile→Val的替换可能影响HSL的活性,最终影响到猪的背膘厚、瘦肉率或肌内脂肪含量。苏姜猪是以地方猪种姜曲海猪、枫泾猪与国外猪种杜洛克猪为亲本、历经10余年培育而成的瘦肉型新品种猪,2013年通过了国家畜禽种质资源委员会品种审定,获得了新品种证书[10]。因此,今后将开展HSL基因外显子I区域内的G→A碱基突变多态与苏姜猪背膘厚、瘦肉率等胴体性状的相关性研究。
参考文献:
[1]Harbitz I, Langset M, Ege A G, et al. The porcine hormone-sensitive lipase gene: sequence, structure, polymorphism and linkage mapping[J]. Animal Genetics, 1999, 30:10-15.
[2]Mellink C H M, Lahbib-Mansais Y, Yerle M, et al. Localization of four new markers to pig chromosomes 1, 6 and 14 by radioactive in situ hybridization[J]. Cytogenet Cell Gnet, 1993, 64:256-260.
[3]Egan J J, Greenberg A S, Min-Kun C, et al. Mechanism of hormone-stimulated lipolysis in adipocytes: translocation of hormone-sensitive lipase to lipid storage droplet[J]. Proc Natl Acad Sci USA, 1992, 89:8534-854.
[4]Otsu K, Khanlla V K, Arhibald A L, et al. Cosegregation of porcine malignant hypertheha and a probable causal mutation in the skeletal musele ryanone receptor gene in back cross tamilies[J]. Genomies, 1991, 11:744-750.
[5]曲 亮,黄瑞华,李开桢,等. H-FABP和HSL基因多态性对苏淮猪胴体性状的影响[J]. 南京农业大学学报, 2008, 31 (3): 107-112.
[6]Knoll A, Stratil A, Nebola M, et al. characterization of a polymorphism in exon I of the porcine hormone-sensitive lipase (LIPE) gene[J]. Animal Genetics, 1998, 29:460-477.
[7]强巴央宗,商 鹏,杨 涛,等. 藏猪激素敏感脂肪酶(HSL)基因遗传多样性分析[J].西南农业学报,2012,25(5):1904-1906.
[8]陈利荣,贾艳梅,韩俊文.BsaH I对猪激素敏感脂肪酶基因外显子I的PCR-RFLP分析[J].生物学杂志,2009,26(3):19-21.
[9]苗志国,李国旺,谢红兵. 金华猪与长白猪脂肪组织中HSL mRNA丰度的差异研究[J]. 广东农业科学,2011(4):128-130.
[10]唐现文,韩大勇,周春宝,等. 苏姜猪四至六世代雌激素受体(ESR)基因PvuⅡ多态性分析[J]. 浙江农业学报,2013,25(5):965-970.
