退化草地大针茅根系特征对氮素添加的响应
2014-03-26秦洁鲍雅静李政海胡志超高伟
秦洁,鲍雅静,李政海,胡志超,高伟
(1.内蒙古大学生命科学学院,内蒙古 呼和浩特010021;2.大连民族学院环境与资源学院,辽宁 大连116600;3.内蒙古自治区乌海市规划局规划设计院,内蒙古 乌海016000)
草原是我国北方分布最广泛的自然生态系统,构成了我国北方强有力的生态安全屏障。长期的开发利用和巨大的人口压力,使我国北方草原生态系统功能严重退化,生态系统呈现由结构性破坏向功能性紊乱的方向发展,由此引起的水资源短缺、水土流失、沙漠化、生物多样性减少等生态问题持续加剧。锡林郭勒草原是我国北方最典型的草原分布区,是环北京生态防护圈的重要组成部分。由于忽视草地资源的科学管理和合理利用,采取掠夺式经营,导致草地生态系统环境恶化、生产力下降、草畜矛盾日益突出、草场退化加剧[1],严重阻碍了草地畜牧业的可持续发展。
大针茅(Stipagrandis)是亚洲中部草原区特有的蒙古草原种,在我国分布较广,是划分草原植被的指示植物[2-3]。大针茅为多年生密丛型旱生草本植物,是良好的饲用牧草[4],是典型草原区重要的畜牧业资源且在维系草原生态系统平衡方面发挥着重要的作用。以大针茅为建群种的草原是蒙古高原典型草原地带广泛分布的地带性植物群落,具有良好的生产性能。近半个世纪以来,由于自然原因加之对草地不合理的利用,已突破草原生态系统健康阈值,造成草原植物群落的优势种更替,饲用品质恶化,植物个体小型化,光合效率显著下降,氮素、碳素等物质循环的平衡失调,群落生物生产力严重衰减等诸多问题,大针茅草原出现不同程度的退化[5],已经引起国内许多学者的重视,并开展了一系列研究。早在20世纪80年代陈佐忠等[6]就对内蒙古草原大针茅群落地下生物量和降水量的关系进行了研究。董亭等[7]探讨了放牧对大针茅生物量的影响。王炜等[8]研究了有关大针茅群落退化机理。万宏伟等[9]对大针茅叶片功能特征与氮素添加的关系进行了研究。目前,国内外对该植物的研究多集中在地上部分,而对其土壤中根系分布特征的研究相对较少。
生态系统地上与地下的关联主要是通过根系实现的,根系是植被地下部分的重要组成,是植物与土壤环境之间进行物质交流、能量交换的主要桥梁[10]。随着全球变化与生态系统研究的深入,对根系的研究逐渐受到重视[11]。国内外学者对植物根系进行了大量的研究,包括植物根系生长特性[12]、分布特征[13]、生态环境效应[14]以及生理特性[15]等。在草原恢复领域,很多生态理论和植物机理研究[16-17],其目的是为了研究和预测植物多样性以及群落之间的关系[18],这需要很多具体的参数来描述根系空间分布,例如根系直径、根系深度、根长度、根表面积等[19-20]。而长期以来,国内外对根系的研究多集中在林木和农作物方面,但是有关草本植物,特别是草地植被根系的研究相对较少[21]。王艳芬和汪诗平[22]研究了不同放牧率对内蒙古典型草原根系的影响,毛齐正等[23]对植物根系的生态功能及其影响因素进行了研究,Andrén和Paustian[24]研究表明天然草地根系主要受降水和地温的影响。大针茅根系构型为密丛型[25-26],其根系构型在不同退化程度草地中的变化情况能直接反映土壤理化性状、土壤水分和家畜采食、践踏等综合环境因素对大针茅个体及种群的影响。周艳松等[3]通过研究不同退化程度草地中大针茅根系构型,发现中度退化草地中大针茅根系构型小型化特别明显,在群落退化演替进程中大针茅的建群种作用逐渐减弱并最终被其他优势种取代。
众所周知,氮是蛋白质、核酸、叶绿素等的重要组成部分,是植物重要的营养成分之一,是半干旱草原植物生长的限制性因素。国内外学者对于根系与氮素的关系做了很多研究,包括根系吸收与运输机制[27]、根系生物量[28-29]以及空间分布特征[30]。于占源等[31]主要研究了氮素添加对科尔沁沙质草地生物多样性和生产力的影响,李禄军等[32]对氮素添加与土壤氮素有效性的关系做了研究,但在氮素影响下草原植物根系构型变化的相关研究在国内还鲜为报道。本研究围绕退化草地恢复为主题,试图通过实验,了解氮素添加对典型草原建群种根系特征的影响。目前添加氮素改善草场质量方面的研究多集中于人工草地[33-35],天然草原添加氮素的研究相对较少,退化草地的外界氮输入量很小,因此氮素添加有可能对退化草地的生态系统的结构和功能产生很大的影响。在追求高产量的今天,为了增加产草量盲目施肥,有的不仅没有起到作用还反其道而行,甚至破坏了原本平衡的土壤环境。本研究以锡林郭勒草原轻度退化草地与重度退化草地为研究对象,采用野外定位实验,根据对典型草原不同退化阶段土壤养分状况的已有研究成果,设计养分(N素)添加梯度的实验,模拟土壤的贫养与富养状态,重点探讨在不同退化程度草原土壤环境条件下,群落建群种大针茅根系构型在养分资源梯度上的发展态势与替变规律,以及不同退化草地中是否适合施肥,并且尽可能找到有助于退化草地恢复最适宜的氮素添加量,以便达到低投入高生产量的双赢效果。
1 材料与方法
1.1 研究区概况
锡林郭勒盟属于中温带半干旱大陆性气候。其主要气候特点是风大、干旱、寒冷。年平均气温0~3℃,气温年较差为35~42℃。大部分地区年降水量200~300mm,自东向西递减,降雨多集中在7、8、9三个月内。年平均相对湿度在60%以下,蒸发量在1500~2700mm之间,由东向西递增,蒸发量最大值出现在5—6月份。年日照时数为2800~3200h,日照率64%~73%。由于地势平坦开阔,土质较好,草场类型多,水草丰富,使锡林郭勒盟拥有发展畜牧业经济的得天独厚的优越自然条件。1997年成为国家级保护区,主要有草甸草原、典型草原、沙地疏林草原和河谷湿地生态系统。以1961-2010年的平均降水量为基础,2009年降水量距平均降水量百分率为-28.4,2010年降水量距平均降水量百分率为-40.9,是50年里降水最少的年份。试验区位于内蒙古锡林郭勒盟白音锡勒牧场境内(N:43°38′、E:116°42′),海拔1187m。典型草原群落是本地区分布最广的草原类型。群落内的植物种类组成比较丰富,主要有大针茅、羊草(Leymuschinensis)、糙隐子草(Cleistogenessquarrosa)和一些杂类草等。
1.2 研究方法
1)实验样地的选取:
本研究选择的实验样地位于中国科学院内蒙古草原生态系统定位研究站研究区样地附近,实验样地设置如下。
轻度退化样地:当地牧民打草场,每年秋季打草1次,于2008年年初围封作为实验样地,属于轻度退化草原,简称为:YY样地。
重度退化样地:牧民长期自由放牧样地,已发生严重退化,于2008年年初围封作为实验样地,简称为:YT样地。
2)YY样地与YT样地的植被与土壤概况:
对轻度退化和重度退化样地进行的本底调查表明(表1),YT样地与YY样地相比土壤更为贫瘠,地上、地下生物量均低于YY样地。

表1 轻度退化样地和重度退化样地植被特征与0~10cm土壤概况Table 1 Vegetation characteristics and 0-10cm soil condition of light degradation and heavy degradation
1.3 实验设计
2008年在上述2块研究样地中各选取100m×100m的草场一块,并用围栏进行了围封,作为定位实验样地,在2块样地内进行了本底调查并设计了相同的N素梯度控制实验区。氮素添加于2009年5月1日和2010年5月1日实施(与生长季相对应),氮素添加水平参照潘庆民等[36]氮素添加实验的处理和实验结果设置。包括0、10.5、17.5和28g N/m24个水平,分别代表对照、低氮、中氮和高氮生境,每个氮素水平设置5个重复。在2块样地内采用随机区组方法施加上述氮素水平的硝酸铵(NH4NO3,其中 N占7/20),即0、30、50、80g NH4NO3/m2稀释于少量水中均匀喷洒实验区(共20个2m×2m的实验小区,每个小区中间设置1个1m×1 m的固定样方,每个小区之间留0.5m的过道)。为了保证氮素是唯一限制因子,对所有处理施用适量KH2PO3以排除P,K限制对本实验的干扰。YT样地氮素添加实验设计原则与YY样地相同。2010年8月中旬大针茅生长旺盛期时,对每个样地的每个处理中分别随机选取3个重复处理样地,挖取深度为50cm的30cm×30cm的土方,放置沙袋中冲洗。带回室内后,在每个处理的沙袋中分别随机选取10株完整的大针茅草样,用米尺测量地上部分高度和根系深度,然后使用Delta-T SCAN根系分析系统的扫描仪对大针茅根系进行扫描,样品图像扫描后输送到计算机中,并利用根系分析仪系统中图像分析软件分析大针茅根系的长度、直径、面积、体积等根系构型特征。
1.4 数据统计分析
用Excel软件整理数据,利用SPSS-statistics 20对数据进行统计分析,采用单因素方差(one-way ANOVA)分析氮素添加梯度对根系特征及地上高度的影响,Duncan假定方差齐性检验用于不同处理之间根系特征的多重比较,采用Pearson Correlation相关分析检测根系特征与地上高度和氮素添加梯度之间的关系,采用独立样本T检验比较YY和YT样地之间根系特征对氮素梯度的差异响应。
2 结果与分析
2.1 大针茅根系构型特征对氮素添加梯度的响应
本研究选取了4个根系构型指标,分别为根系面积、根系体积、根系长度和根系直径,研究其在不同退化程度的草地随氮素添加梯度的变化规律,结果表明,单因素方差分析显示重度退化样地中根系面积在不同处理之间差异显著(P<0.05),根系面积在80g/m2处理下显著高于其他3个添加梯度,0,30,50g/m2处理之间无显著差异(图1),相关分析表明(表2),重度退化样地中根系面积与氮素添加梯度呈极显著正相关(r=0.474,P<0.01)。在轻度退化样地中不同处理之间根系面积无显著差异。
单因素方差分析显示重度退化样地中大针茅根系体积在不同处理之间差异显著(P<0.05),根系体积在80g/m2处理显著高于0和30g/m2两个处理(图2),而与50g/m2处理差异不显著。相关分析表明(表2),重度退化样地中大针茅根系体积与氮素添加梯度呈极显著正相关(r=0.474,P<0.01),在轻度退化样地不同处理之间的根系体积无显著差异。
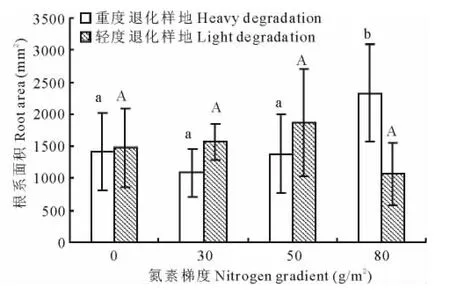
图1 大针茅根系面积对氮素添加梯度的响应Fig.1 The response of S.grandis root surface area to nitrogen addition rate
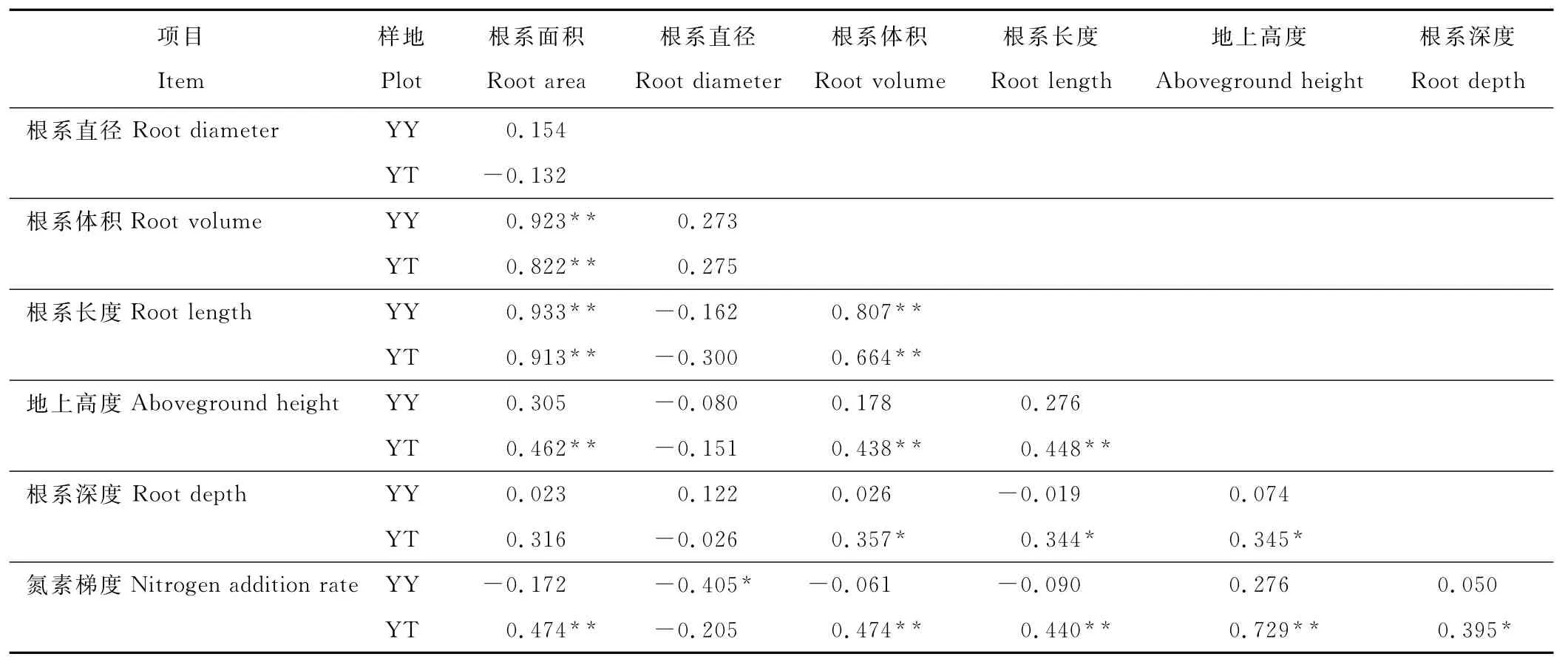
表2 大针茅根系特征与氮素添加梯度的相关性分析Table 2 Correlation analysis between S.grandis root characteristics and nitrogen addition rate
对于大针茅根系长度,不同氮素添加梯度之间,在重度退化样地中有显著差异(P<0.05),在轻度退化样地中无显著差异(图3)。重度退化样地在80g/m2处理下显著高于其他3个添加梯度(0,30,50g/m2)。相关分析表明(表2),重度退化样地中大针茅根系长度与氮素添加梯度呈极显著正相关(r=0.440,P<0.01),而在轻度退化样地中无显著相关性。
不同氮素添加梯度之间比较,大针茅根系直径在轻度退化样地和重度退化样地中均没有显著差异(图4)。但相关分析显示(表2),轻度退化样地中大针茅根系直径与氮素添加梯度呈显著负相关(r=-0.405,P<0.05),重度退化样地中根系直径与添加氮素梯度无显著相关关系。
相关分析表明(表2),2种退化程度样地中,根系长度与根系面积、体积三者之间都呈极显著正相关关系,根系直径与面积、体积、长度之间没有显著相关关系。根系图像分析软件主要是通过根系的直径和长度来自动换算根系体积和面积,这说明根系面积、体积的变化主要是由根系长度的变化引起的。
2.2 大针茅地上高度与根系深度对氮素添加梯度的响应
不同氮素添加梯度之间,大针茅地上高度在重度退化样地中有显著差异(P<0.05),在轻度退化样地中差异不显著(图5)。重度退化样地中,大针茅地上高度在80g/m2处理下显著高于其他3个添加梯度(0,30,50 g/m2),30g/m2处理也显著高于0g/m2;相关分析表明(表2),大针茅重度退化样地中地上高度与氮素添加梯度呈极显著正相关(r=0.729,P<0.01),在轻度退化样地中无显著相关性。
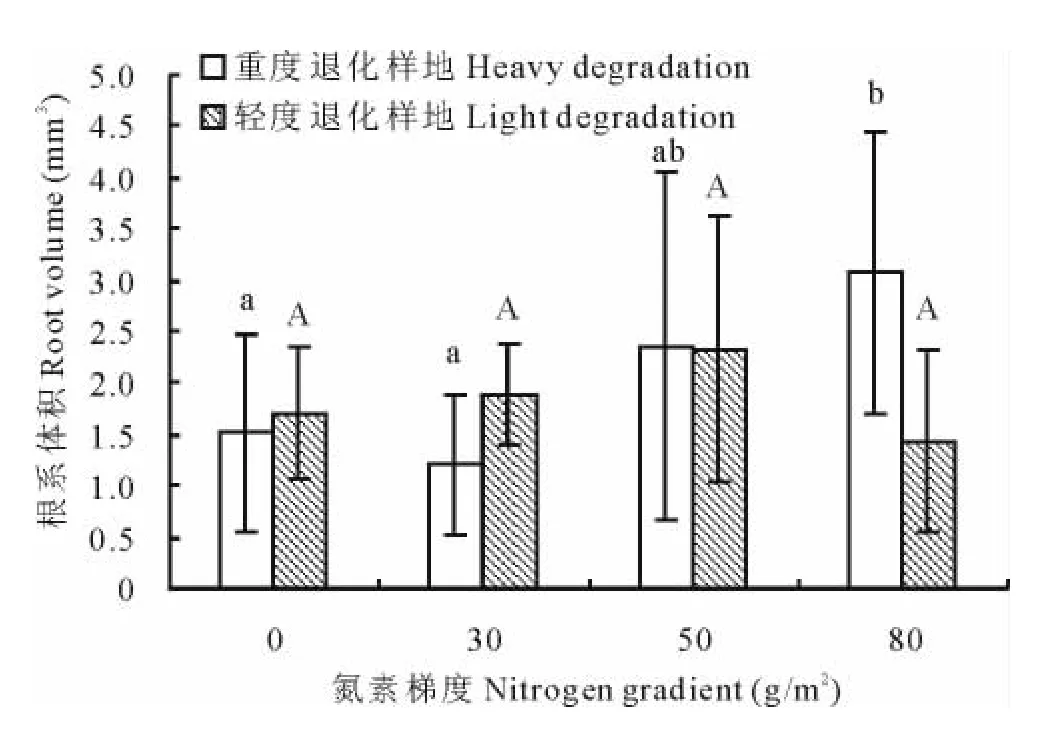
图2 大针茅根系体积对氮素添加梯度的响应Fig.2 The response of S.grandis root volume to nitrogen addition rate
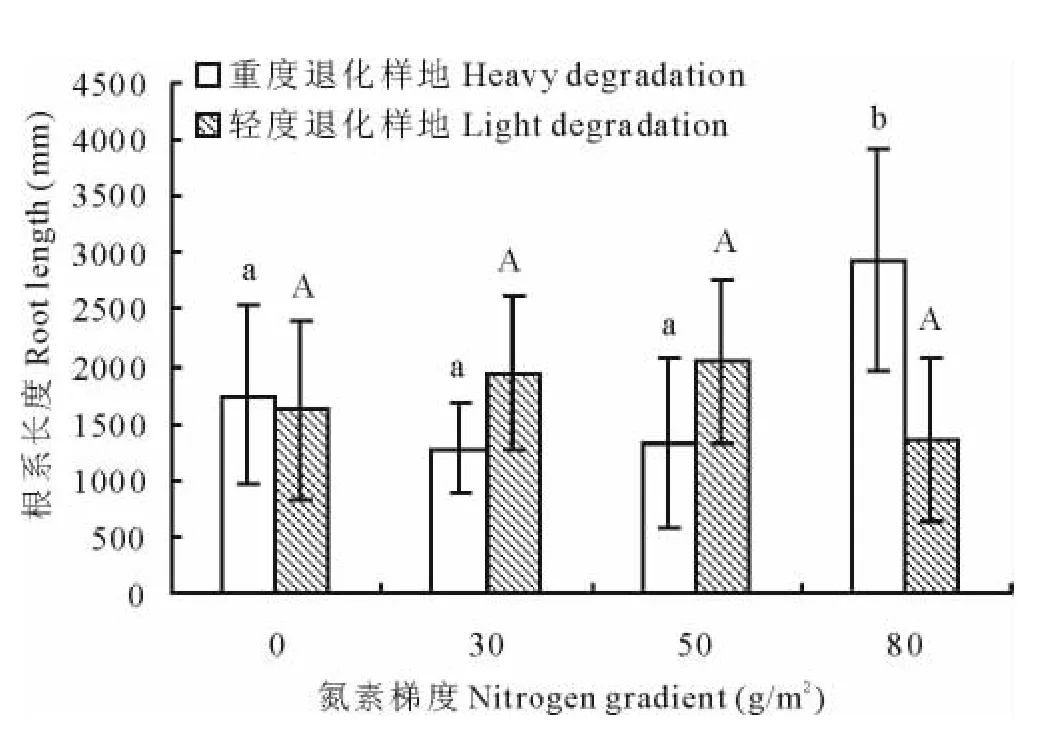
图3 大针茅根系长度对氮素添加梯度的响应Fig.3 The response of S.grandis root length to nitrogen addition rate
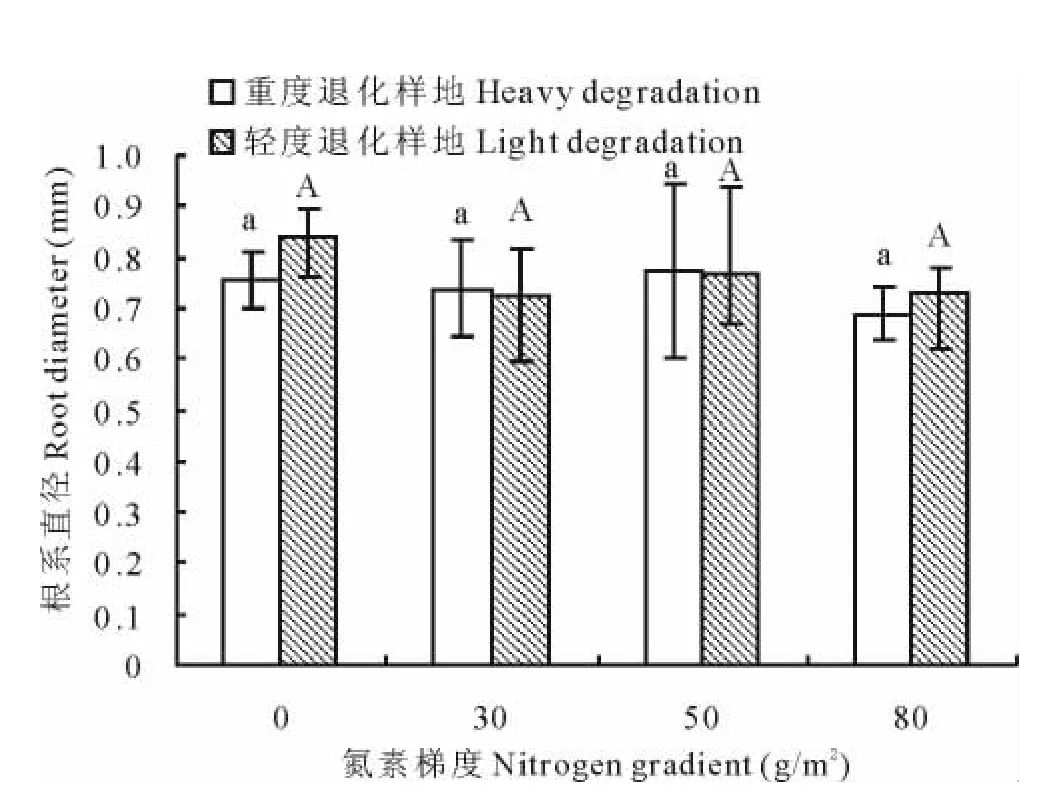
图4 大针茅根系直径对氮素添加梯度的响应Fig.4 The response of S.grandis root diameter to nitrogen addition rate

图5 大针茅地上高度对氮素添加梯度的响应Fig.5 The response of S.grandis height to nitrogen addition rate
对于大针茅根系深度而言,不同氮素添加梯度之间比较,重度退化样地中有显著差异(P<0.05),在轻度退化样地中差异不显著(图6)。重度退化样地中,大针茅根系深度在80g/m2处理显著高于0g/m2处理,而与30和50g/m2处理差异不显著。相关分析(表2)显示,大针茅重度退化样地中根系深度与氮素添加梯度呈显著正相关(r=0.395,P<0.05),在轻度退化样地中无显著相关性。

图6 大针茅根系深度对氮素添加梯度的响应Fig.6 The response of S.grandis root depth to nitrogen addition rate
地上高度在重度退化样地中与根系长度、面积、体积都呈极显著正相关关系,与根系深度呈显著正相关关系,轻度退化样地中这种相关性不显著(表2)。根系深度在重度退化样地中与根系长度、体积都呈显著正相关关系,说明氮素添加有利于根系在土壤中深度的伸长以及侧根的生长。
2.3 轻度退化样地和重度退化样地大针茅根系特征比较
YY样地和YT样地比较(图1~6),根系体积、面积、长度、根系深度和地上高度在前3个添加梯度(0,30,50g/m2)均未达到显著差异,80g/m2处理时 YT样地显著高于YY样地。根系直径在后3个添加梯度(30,50,80g/m2)中无显著差异,在0g/m2处理时YY样地显著高于YT样地。
说明在没有添加氮素的自然情况下,大针茅的根系直径在轻度退化草地要高于重度退化草地,根系体积、面积、长度、根系深度和地上高度没有显著差异,氮素的添加更有利于重度退化草地中大针茅根系的增粗生长,在低氮素添加条件下(30,50g/m2),2个样地大针茅的伸长生长差异不显著,但在高氮素添加的条件下(80g/m2),重度退化样地中大针茅的伸长生长显著高于轻度退化样地。
3 讨论
土壤系统与植被是一个相互作用相互影响的整体,不同退化程度的草地对氮素添加的响应不同。其中重度退化草地的响应更明显,随氮素添加量的增加,大针茅根系变长,面积、体积也随之增加,地上高度和根系深度也相应增加,但是根系直径则无明显变化,这表明土壤中氮素的增加,促使大针茅根系通过增加根系长度和侧根的生长扩大根系空间分布使之能够吸收足够的养分供自身生长。研究表明在高氮素添加(80g/m2)情况下,重度退化草地中大针茅根系长度、面积、根系深度和地上高度均显著高于轻度退化草地。对于轻度退化草地,土壤中氮素添加的变化没有引起大针茅根系的变化,这表明在轻度退化草地中,大针茅根系的生长处于稳定状态。
研究根系的特征具有重要意义:根系面积直接反映了根与土壤的结合面积,结合面积越大则越利于根系对营养物质的吸收[3];根系长度反映了根系生长及其在土壤中的拓殖能力[3];根系直径大小与根导水阻水有直接关系,直径越大,阻力越大[37]。梅莉等[38]通过对水曲柳(Fraxinusmandshurica)根系的研究表明,根系吸收机制是在比较肥沃的土层中尽可能的吸收较多的碳水化合物,扩大根系与土壤的接触面积,这与重度退化草地对氮素添加的研究结果相一致。但在轻度退化草地中,大针茅根系特征与地上高度对氮素添加的响应不显著。但施氮量过高则影响植被生长,造成生物量减少[39],对轻度退化草地的研究结果也证明了一种假说:高氮添加的植物根系不再需要更大的能力来汲取养分[40],故其地下生物量会减少,这也说明植被在适应氮素上存在一个阈值。
不同退化草地的研究结果差异,反映了大针茅根系形态在不同退化状态下对氮素的响应机制不同。在重度退化草地中对氮素的响应显著,可能是由于重度退化草地土壤中养分不足,氮素的添加充实了土壤中的养分,根系通过增加其长度,扩大表面积,提高根系活力,使其能够从土壤中吸收更多的营养物质。轻度退化草地对氮素添加的响应不显著,只有根系直径与氮素添加保持负线性关系。轻度退化草地中,由于土壤退化滞后于植被退化,土壤保水性良好,含充足养分,大针茅根系对于土壤中养分的变化具有较好的自我调节能力,所以对氮素添加的响应不明显,氮素的添加对其没有显著意义。随着草地退化程度的恶化,大针茅根系面积、体积、长度以及地上高度、根系深度之间的联系更加紧密,相互作用进一步增强。综合来看土壤特性的变化与根系特征的变化是相辅相成的,一方面是大针茅根系会根据土壤养分变化作出适应性反应,另一方面大针茅根系改变了其生存的土壤条件,两者相互作用相互影响达到了平衡的对大针茅自身生长有益的生长状态,这也是大针茅对环境进一步适应的最终结果。
适宜的水肥条件会促进根系发育,提高根系生理活性,促进水分养分的吸收[41]。研究发现,一年生植物是高度顺应降水波动的特殊类群[42],而不良的水肥供应如只施氮不施水抑制了根系的发育,降低水分养分的吸收。在水分胁迫严重情况下,施肥会增加土壤中养分浓度,降低土壤水势,进一步加剧水分胁迫,这种严重的水分胁迫是氮素添加表现出不良作用的根本原因。植被根系对于水分和养分的吸收是相互的,而本研究只做了氮素单一因子添加对根系的影响,只单一添加养分可以提高植被生产力,但并不能使植物生产达到最佳状态。水分与养分对根系的共同作用以及影响的显著程度,以及不同的退化草地中差异响应都需要进一步的研究。综合推测在不同水分条件下土壤中氮素的添加对根系的影响程度也是不同的。
大针茅是典型草原的主要物种,研究大针茅根系对土壤养分的响应可为退化草地人工恢复以及提高大针茅产草量提供依据和借鉴。在不同退化程度的草地,需要补充养分的量是不同的,对于轻度退化草地不需要添加养分或者添加少量养分便会为植被地下部分的生长提供最佳生长条件,而在重度退化草地中,则需要补充一定量的养分才会满足植被生长所需。随着土壤中养分的增加,大针茅根系逐渐扩大,以及地上高度的增加,最终导致整个植物体的大型化,并在群落演替过程中逐渐取得优势地位。从这种意义上讲,对于严重退化的草地可以采用施一定量的氮肥,保护和减少退化草地的进一步退化,并会促进群落的恢复演替。
4 结论
随着氮素添加量的增加,轻度退化样地的大针茅根系特征没有显著变化,而重度退化样地中变化显著。在重度退化样地中,根系长度、面积、体积均随氮素的增加而显著增加,三者之间呈极显著正相关关系;根系直径对氮素添加却没有明显变化;地上高度与根系深度也随氮素添加的增加而显著增加,且地上高度与根系深度呈显著正相关关系。这表明重度退化样地中土壤氮素的增加,促使大针茅根系主要通过增加根系长度扩大根系在土壤的空间分布。
比较不同退化程度的草地,在无氮素添加的条件下,大针茅根系直径在重度退化样地显著低于轻度退化样地;在高氮素添加下(80g/m2),大针茅根系长度、面积、体积以及地上高度和根系深度均在重度退化样地中显著高于轻度退化样地。说明在低氮素添加处理下,不同退化程度大针茅的根系特征几乎没有明显差异,当氮素添加到一定量时,重度退化样地的根系主要通过伸长生长与轻度退化样地表现出明显差异。
[1] 宝音陶格涛,王静.退化羊草草原在浅耕翻处理后植物多样性动态研究[J].中国沙漠,2006,26(2):232-237.
[2] 中国科学院内蒙古宁夏综合考察队.内蒙古植被[M].北京:科学出版社,1985.
[3] 周艳松,王立群,张鹏,等.大针茅根系构型对草地退化的响应[J].草业科学,2011,28(11):1962-1966.
[4] 内蒙古植物志编辑委员会.内蒙古植物志第五卷[M].呼和浩特:内蒙古人民出版社,1994.
[5] 李博.中国北方草地退化及其防治对策[J].中国农业科学,1997,30(6):2-10.
[6] 陈佐忠,黄德华,张鸿芳.内蒙古锡林河流域羊草草原和大针茅草原地下生物量与降水量关系模型探讨[A].草原生态系统研究(第2集)[M].北京:科学出版社,1988:20-25.
[7] 董亭,李群,赵萌莉,等.放牧对大针茅根系生物量影响的研究[J].草地学报,2011,19(2):237-241.
[8] 王炜,梁存柱,刘钟龄,等.羊草+大针茅草原群落退化演替机理的研究[J].植物生态学报,2000,24(4):468-472.
[9] 万宏伟,杨阳,白世勤,等.羊草草原群落6种植物叶片功能特性对氮素添加的响应[J].植物生态学报,2008,32(3):611-621.
[10] 李鹏,李占斌,澹台湛.黄土高原退耕草地植被根系动态分布特征[J].应用生态学报,2005,16(5):849-853.
[11] 韦兰英,上官周平.黄土高原不同演替阶段草地植被细根垂直分布特征与土壤环境的关系[J].生态学报,2006,26(11):3740-3748.
[12] Baddeley J A,Watson C A.Seasonal patterns of fine-root production and mortality inPrunusaviumin Scotland[J].Canadian Journal of Forest Research,2004,34(7):1534-1537.
[13] 赵忠,李鹏.渭北黄土高原主要造林树种根系分布特征及抗旱性研究[J].水土保持学报,2002,16(1):96-99,107.
[14] 李勇,徐晓琴,朱显谟.黄土高原植物根系提高土壤抗冲性机制初步研究[J].中国科学(B辑 化学 生命科学 地学),1992,22(3):254-259.
[15] 蒋乔峰,陈静波,宗俊勤,等.盐胁迫下磷素对沟叶结缕草生长及Na+和K+含量的影响[J].草业学报,2013,22(3):162-168.
[16] Leadley P W,Reynolds J F,Chapin F S I.A model of nitrogen uptake byEriophorumvaginatumroots in the field:ecological implications[J].Ecological Monographs,1997,67(1):1-22.
[17] Somma F,Hopmans J W,Clausnitzer V.Transient three-dimensional modeling of soil water and solute transport with simultaneous root growth,root water and nutrient uptake[J].Plant and Soil,1998,202(2):281-293.
[18] Caldwell M,Manwaring J,Durham S.Species interactions at the level of fine roots in the field:influence of soil nutrient heterogeneity and plant size[J].Oecologia,1996,106(4):440-447.
[19] Buysse J,Smolders E,Merckx R.Modelling the uptake of nitrate by agrowing plant with an adjustable root nitrate uptake capacity[J].Plant and Soil,1996,181(1):19-23.
[20] Grant R.Simulation in ecosys of root growth response to contrasting soil water and nitrogen[J].Ecological Modelling,1998,107(2):237-264.
[21] Carroll C,Merton L,Burger P.Impact of vegetative cover and slope on runoff,erosion,and water quality for field plots on a range of soil and spoil materials on central Queensland coal mines[J].Soil Research,2000,38(2):313-328.
[22] 王艳芬,汪诗平.不同放牧率对内蒙古典型草原地下生物量的影响[J].草地学报,1999,7(3):198-203.
[23] 毛齐正,杨喜田,苗蕾.植物根系构型的生态功能及其影响因素[J].河南科学,2008,26(2):172-176.
[24] Andrén O,Paustian K.Barley straw decomposition in the field:a comparison of models[J].Ecology,1987,68(5):1190-1200.
[25] 陈世鍠,张昊,王立群.中国北方草地植物根系[M].长春:吉林大学出版社,2001.
[26] Staswick P,Serban B,Rowe M,etal.Characterization of anArabidopsisenzyme family that conjugates amino acids to indole-3-acetic acid[J].Plant Cell,2005,17(2):616-627.
[27] Hopmans J,Bristow K.Current capabilities and future needs of root water and nutrient uptake modeling[J].Advances in Agronomy,2002,(77):103-183.
[29] Frank D,Groffman P.Plant rhizospheric N processes:what we don’t know and why we should care[J].Ecology,2009,90(6):1512-1519.
[30] Iversen C,Bridgham S,Kellogg L.Scaling plant nitrogen use and uptake efficiencies in response to nutrient addition in peatlands[J].Ecology,2010,91(3):693-707.
[31] 于占源,曾德慧,艾桂艳,等.添加氮素对沙质草地土壤氮素有效性的影响[J].生态学杂志,2007,26(11):1894-1897.
[32] 李禄军,曾德慧,于占源,等.氮素添加对科尔沁沙质草地物种多样性和生产力的影响[J].应用生态学报,2009,20(8):1838-1844.
[33] 张耀生,赵新全,黄德清.青藏高寒牧区多年生人工草地持续利用的研究[J].草业学报,2003,12(3):22-27.
[34] 李志坚,周道玮,胡跃高.不同施肥水平与组合对饲用黑麦生产性能的影响研究 Ⅱ对饲用黑麦质量的影响[J].草业学报,2005,14(4):72-81.
[35] 钟小仙,江海东,曹卫星,等.施肥和刈割日期对杂交狼尾草钙、磷、镁含量的影响及其与家畜需要的关系[J].草业学报,2005,14(5):87-91.
[36] 潘庆民,白永飞,韩兴国,等.内蒙古典型草原羊草群落氮素去向的示踪研究[J].植物生态学报,2004,28(5):665-671.
[37] 严小龙.根系生物学原理与应用[M].北京:科学出版社,2007.
[38] 梅莉,王政权,韩有志,等.水曲柳根系生物量、比根长和根长密度的分布格局[J].应用生态学报,2007,17(1):1-4.
[39] 马霞,王丽丽,李卫军,等.不同施氮水平下接种根瘤菌对苜蓿固氮效能及种子生产的影响[J].草业学报,2013,22(1):95-102.
[40] 樊维,蒙荣,陈全胜.不同施氮水平对克氏针茅草原地上地下生物量分配的影响[J].畜牧与饲料科学,2010,31(2):74-76.
[41] 宋海星,李生秀.水、氮供应和土壤空间所引起的根系生理特性变化[J].植物营养与肥料学报,2004,10(1):6-11.
[42] 闫建成,梁存柱,付晓玥,等.草原与荒漠一年生植物性状对降水变化的响应[J].草业学报,2013,22(1):68-76.
