秦岭北坡栓皮栎干基萌苗生长特性研究
2014-03-26薛瑶芹张文辉马莉薇周建云
薛瑶芹,张文辉,马 闯,马莉薇,周建云
(西北农林科技大学 西部环境与生态教育部重点实验室,陕西 杨凌 712100)
森林被采伐或者受到其他干扰后,可以通过伐前更新幼苗的生长、种子库中种子的萌发成苗、伐桩萌发或者根系萌发而不断恢复[1]。主干基部萌苗或者干基萌苗(Stem base sprouts)是在存活的植物个体主干基部产生的萌苗[2-3]。作为伐前更新幼苗的一种类型,许多植物都可以产生干基萌苗,例如,在Burkina Faso的萨瓦纳林地(Savanna-woodland)中,非洲酸枝(BurkeaAfricanaHook.)、AcaciadudgeoniCraib. 和DetariummicrocarpumGuill.& Perr.等植物均可以产生干基萌苗[1],该研究中称其为徒长萌苗(Water sprouts)。另外,在智利南部的雨林中,尤其是不成熟的森林中,干基萌苗是植物较常见的无性繁殖方式,能够产生干基萌苗的植物包括AmaiouaguianensisAubl.、ClethrascabraPers.和ErythroxylumamplifoliumBaill.等[3]。
栓皮栎(QuercusvariabilisBl.)在秦岭和大别山区域数量较多,密度也较大,形成了地带性植被,是关系到当地林业和生态建设的重要树种[4]。然而,为了获取软木、栲胶、木材和薪炭材,多年的过度利用和抚育缺乏,使该区域的栓皮栎林大多沦为残败次生林,种群数量和质量不断下降,生态功能也大幅降低,栓皮栎种群有逐渐转移退缩的趋势[4-5]。在干扰严重的次生林中,无性繁殖是栓皮栎更新和恢复的主要途径[6-7],由伐桩、主干基部或者根系上的不定芽、休眠芽萌发形成的萌生苗,是栓皮栎自然更新幼苗的重要组成部分[8]。自然条件下,栓皮栎能够产生干基萌苗,但其数量较少,对种群更新的贡献较小[7],因此相关研究也较少。然而,在以采伐利用为目的的栓皮栎次生林中,采伐作业后全部林地或者部分林地会有一段裸露时间,而拥有一定数量的干基萌苗则有利于缩短林地裸露时间,并保证栓皮栎能占据原有生态位。
然而,栓皮栎产生的干基萌苗是如何生长的以及如何通过人工措施促进其萌苗,一直未见相关的研究。因此,本研究在位于秦岭北坡的栓皮栎天然次生林中,对不同大小的栓皮栎个体进行环剥,人为造成机械损伤,并通过持续观测和调查,研究栓皮栎干基萌苗的产生及生长特点,以便制定合理的促进栓皮栎林伐前更新的计划和措施。
1 材料与方法
1.1 研究区概况
本研究在陕西省周至县楼观台(107°39′~108°37′E,33°42′~34°14′N)进行。该区位于秦岭北坡,属温带大陆性季风气候,年平均气温为13.2 ℃,年均降水量为650~800 mm,无霜期225 d。土壤为山地褐土。栓皮栎为乔木层的优势树种,伴生植物有20多种,乔木层主要植物有侧柏(Platycladusorientalis(Linn.) Franco)、油松(PinustabulaeformisCarr.)、槲树(QuercusdentataThunb.)、槲栎(QuercusalienaBlume);主要的灌木有黄栌(CotinuscoggygriaScop.)、绿叶胡枝子(LespedezabuergeriMiq.)、美丽胡枝子(Lespedezaformosa(Vog.) Koehne)、榛(CorylusheterophyllaFisch. ex Trautv.)、黄蔷薇(RosahugonisHemsl)、小花扁担杆(Grewiabilobavar.parviflora(Bunge) Hand.-Mazz.);主要的草本植物有苔草(CarextristachyaThunb. )、紫菀(AstertataricusL.f.)和麦冬(Ophiopogonjaponicus(L.f.) Ker Gawl)等。
该区域的栓皮栎次生林于1985年皆伐后自然形成,郁闭度为75%~80%,栓皮栎占林分组成的90%以上。栓皮栎林林相较为整齐,林分地势较为平坦,栓皮栎个体大小不一,有较大范围的胸径分布。
1.2 环剥母树的选择及环剥处理
2009-03,选择地势平坦、长势较好的栓皮栎次生林(0.5 hm2)进行试验。对林内高度>1.3 m 的栓皮栎进行测量,测量内容包括树高和胸径(DBH)。林中,栓皮栎的胸径为4.5~22.4 cm,树高为1.3~13.5 m。根据胸径测量结果和栓皮栎株数在各胸径上的分布,并参照前人研究对辽东栎(QuercuswutaishanicaBlume)、栓皮栎伐桩径级的划分[9-10],以及Kammesheidt[11]的研究中对多种有萌生能力的树种的径级划分方法,本研究将林地内的栓皮栎个体划分为4个径级,每个径级的胸径范围为:1径级.4.5 cm ≤胸径<10 cm;2径级.10 cm≤胸径<15 cm,3径级.15 cm≤胸径<20 cm,4径级.20 cm≤胸径<25 cm。在每个径级中选择10株母树作为试验对象并进行标记。试验母树概况见表1。试验处理为对树干基部进行环状剥皮(简称环剥,下同),即环状剥去树干基部离地面10 cm处的韧皮部,环剥带宽为5 cm[12]。环剥后对伤口不进行任何处理。
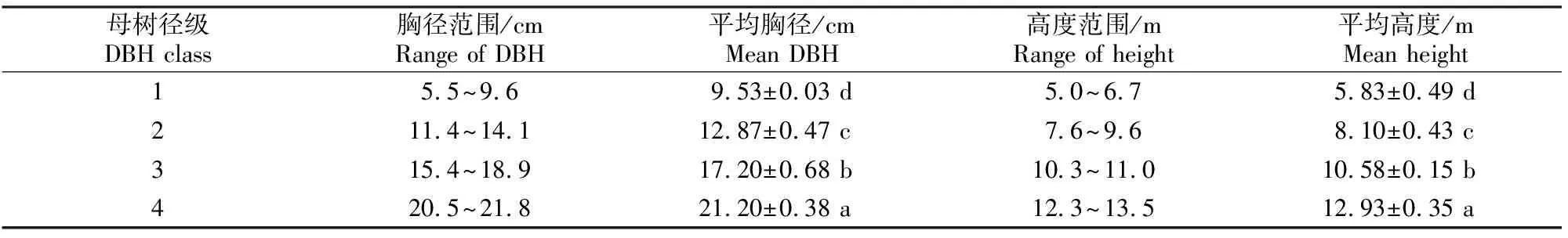
表 1 秦岭北坡不同径级栓皮栎环剥母树的树高及胸径
1.3 干基萌苗的产生及生长观测
观测持续2个生长季。分别于2009-04-15、06-17和09-16及2010-06-19、09-20对环剥母树进行观测,观测及记录内容包括:母树是否产生愈伤组织、是否依然存活、是否产生萌苗;萌苗产生的位置(环剥带上方或者下方)、活萌苗数量、枯死萌苗数量;每株萌苗的年龄(1年生或2年生)、高度、基径和冠幅。萌苗年龄可以根据萌苗主茎树皮的颜色以及其上的芽鳞痕来判断。
1.4 数据的计算与统计分析
综合2009年和2010年的调查数据,计算不同调查时间、不同径级母树中,产生活干基萌苗的母树所占比例(母树萌发率),通过单因素方差分析(One-way ANOVA)比较不同径级母树产生的干基萌苗数量、萌苗枯死率以及干基萌苗的生长状况。
萌苗枯死率=每株母树上枯死萌苗数/总萌苗数。
计算每株母树上1年生和2年生干基萌苗的比例,然后在同一径级母树间计算1年生和2年生干基萌苗比例的平均值,并进行单因素方差分析(One-way ANOVA)和多重比较,绘制不同径级母树的干基萌苗年龄结构图。
所有数据的统计分析均在SPSS 13.0和Excel 2003软件上进行,并在P=0.05水平上检验统计差异的显著性,用Origin 8.0软件制图。
2 结果与分析
2.1 环剥后栓皮栎母树愈伤组织的产生状况
2009-04-15(环剥后1个月)调查发现,所有处理母树均能在环剥伤口处形成愈伤组织,伤口没有腐烂。环剥后2个生长季中,虽然有的母树未能产生干基萌苗,但是环剥伤口一直有愈伤组织保护,因此未发现伤口腐烂或者母树死亡的情况。
2.2 环剥后栓皮栎母树的萌发率
环剥后1个月,所有环剥母树均未产生干基萌苗,因此,只针对2009-06-17、09-16和2010-06-19、09-20的观测记录干基萌苗进行统计分析。
据观测,78%的环剥栓皮栎均能在跟踪观测期间产生干基萌苗,而且所有萌苗均出现在环剥带下方,分布在环剥伤口下边缘到根颈这一范围内;70%的母树能在环剥后2年内保存有活萌苗。
由表2可知,不同径级母树产生萌苗的时间有差别,产生干基萌苗的母树所占比例(母树萌发率)也在环剥后2年内表现出一定波动:1径级母树在环剥3个月(2009-09-16)后产生萌苗,而且母树萌发率随着环剥后时间的延长而有所下降(从100.0%降低到60.0%);2径级和3径级母树萌发率较为稳定,基本保持在80.0%;4径级母树在环剥后第1年没有产生萌苗,环剥后第2年60.0%的环剥个体产生了干基萌苗。总体而言,4个径级的环剥母树中,2径级和3径级母树的萌发率最高。
2.3 环剥后栓皮栎母树干基萌苗的枯死率
干基萌苗枯死率可以反映同一株母树上萌苗间的竞争状况以及母树对于萌苗的保存能力。由表2可知,萌苗枯死发生在环剥后第2年,环剥当年产生干基萌苗的母树均能够保存萌苗。4个径级母树中,2径级(胸径≥10~<15 cm)母树的萌苗枯死率在环剥后2年内均为0.0%;1径级、3径级和4径级母树的萌苗枯死率随着环剥时间的延长而不断上升,但3径级母树的萌苗枯死率较低,只有18.7%~24.7%。
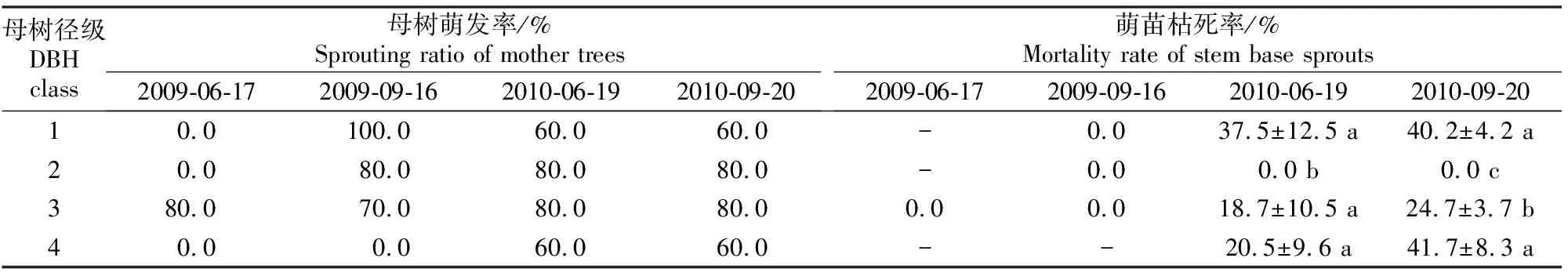
表 2 秦岭北坡不同径级栓皮栎环剥母树的萌发率与干基萌苗枯死率
2.4 环剥栓皮栎母树干基萌苗的数量
图1显示,无论哪个径级的母树,环剥后第2年(2010年)单株母树的干基萌苗数量均多于环剥后当年(2009年)。在4个调查时间中,单株母树的干基萌苗数量在各径级间有所波动,2径级和3径级母树的干基萌苗数量相对较多。2~4径级母树中,干基萌苗数量随母树径级增大总体呈减少趋势,这可能与植物组织的分生能力随年龄增长而下降有关。单因素方差分析结果表明,母树径级(年龄)造成干基萌苗数量存在显著差异(P<0.05)。
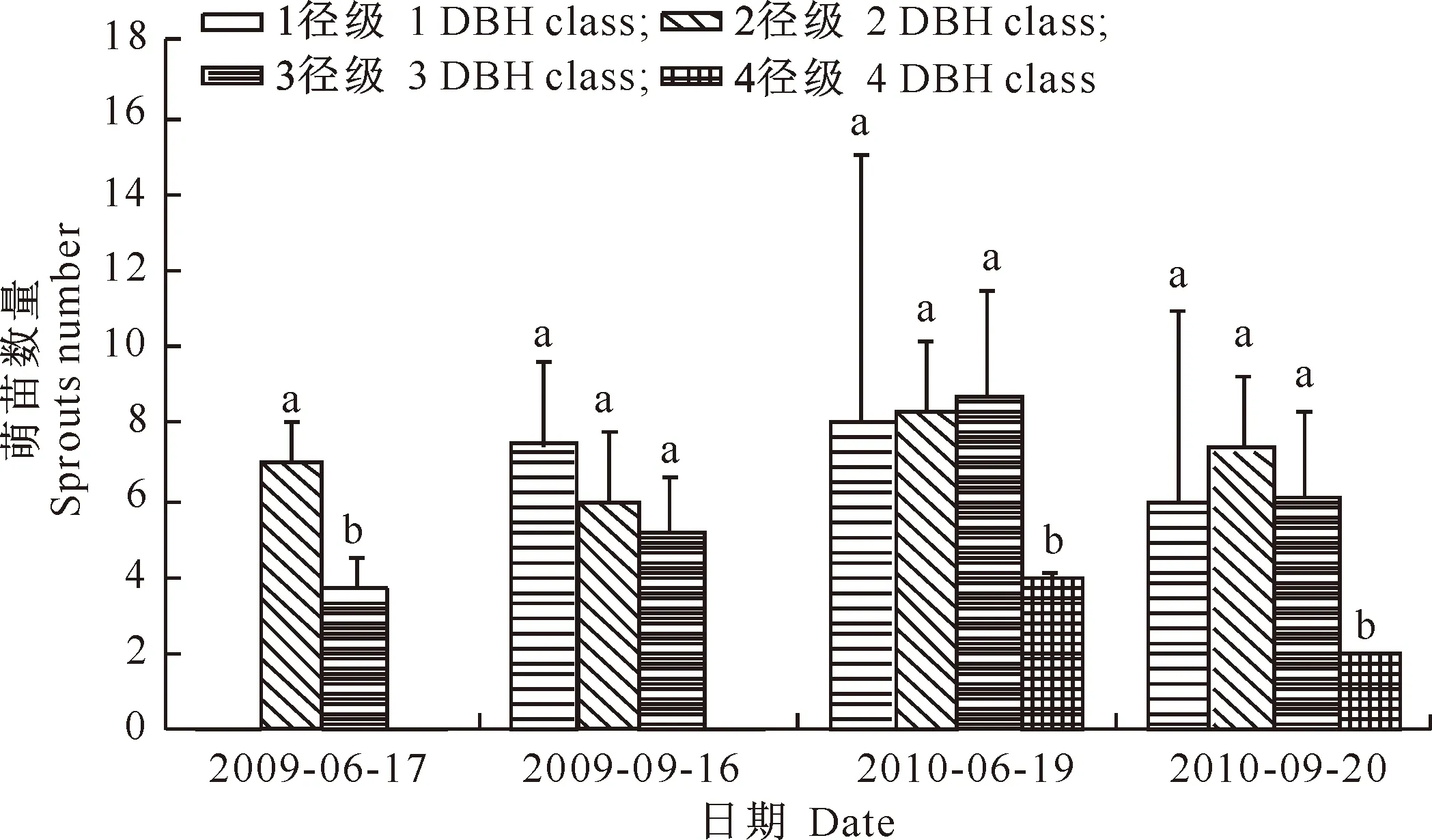
图 1 秦岭北坡不同径级栓皮栎单株母树产生的干基萌苗数量
2.5 环剥后栓皮栎母树干基萌苗的生长
由图2可知,2径级母树的萌苗高度和冠幅均大于其他径级母树的萌苗(P<0.05),而且在环剥后2年中,该径级母树上萌苗的高度和冠幅生长一直优于其他母树上的萌苗(图2-A,B)。不同时期的萌苗基径在4个径级母树间有所不同,2径级母树的萌苗基径在环剥初期(1年内)与其他母树萌苗的基径基本一致,但环剥后第2年,该径级母树的萌苗基径明显大于其他径级母树(图2-C)。
2.6 栓皮栎母树干基萌苗的年龄结构
图3显示了环剥后第2年不同径级母树上1年生和2年生干基萌苗的比例。从图3可以发现,各径级母树在环剥后第2年仍然能够产生当年生萌苗,说明其休眠芽的萌发能力至少能保持到环剥后第2个生长季。不同径级母树对萌苗的保存能力有所不同,除1径级母树外,其他径级母树所产生的1年生和2年生干基萌苗比例差异较大,2径级母树的2年生萌苗远多于1年生萌苗,3径级母树则相反,4径级母树只有1年生萌苗。另外,1年生萌苗所占比例在各径级母树间差异显著(P<0.05),2年生萌苗比例也表现出相同趋势(P<0.05)。

图 2 秦岭北坡不同径级栓皮栎母树上干基萌苗的高度(A)、冠幅(B)和基径(C)

图 3 秦岭北坡不同径级栓皮栎母树的干基萌苗年龄结构
3 讨 论
萌苗一般是植物体对破损或者环境条件的巨大改变而作出的应激反应[13-14]。干基萌苗由树干破损后根颈处被压制的休眠芽或者不定芽萌发产生[3,15],本研究中环剥造成了树干基部的机械损伤,因而大部分栓皮栎(78%)对这一损伤迅速作出反应,即在环剥伤口处产生愈伤组织并在树干基部产生萌苗,这一现象与前人在其他树种中(A.guianensis,C.scabra,E.amplifolium,QuercusmongolicaFisch.)观测到的结果一致[3,16]。根据观测,栓皮栎的干基萌苗全部出现在环剥带下方。丁国华等[12]发现,杉木(CunninghamialanceolataHook.)环剥后萌苗的出现位置与此相同;Mwavu等[15]的研究也指出,树体损伤激发的植物萌苗一般会出现在破损位置的下方。造成这种现象的原因可能是:1)靠近破损位置的休眠芽或者不定芽的活力最强[17];2)本研究中环剥位置位于根颈的上部,而根颈上有大量休眠的子叶芽[14];3)植物内源激素中的细胞分裂素是休眠芽初始萌发的先决条件[18],而细胞分裂素在根系中合成,环状剥皮去掉了树干的韧皮部,阻断了地上部分和地下部分运输营养物质的通道,因此细胞分裂素在环剥带下方累积,促进了主干基部的休眠芽萌发。
本研究中,环剥母树的胸径(年龄)对干基萌苗产生的时间和萌苗数量有较大影响。栓皮栎母树产生干基萌苗的能力随着胸径增大总体上先增加后减小,2径级和3径级(胸径为10~20 cm)的栓皮栎母树在环剥后容易产生干基萌苗,母树萌发率为70.0%~80.0%,而且在环剥后2个月就能够产生萌苗,单株母树干基萌苗数量也比1径级和4径级母树多。有关母树产生干基萌苗能力的研究较少,但类似的规律可以在其他树种的伐桩萌发过程中发现,如胡桃楸(JuglansmandshuricaMaxim.)、白色桦树(BetulapubescensEhrh.)、黄檗(PhellodendronamurenseRupr.)、银色桦树(BetulapendulaRoth)、水曲柳(FraxinusmandschuricaRupr.)和樱皮栎(QuercuspagodaRaf.)等树种的伐桩萌苗能力均随着伐桩的增大而呈现先增大后减小的规律[19-21],即直径较大的伐桩或者母树的萌苗能力较弱,这主要是因为萌苗由休眠芽和不定芽萌发产生,起源于原生分生组织的休眠芽数量有限,其数量随着树木的生长而逐渐减少[22],其活力也随着母树的衰老而下降[23-24],而不定芽起源于次生分生组织,直径较大的母树产生不定芽的能力与机会则较少[22]。本研究中,胸径最大的4径级母树在环剥后第1年完全没有产生萌苗,在第2年只有60.0%的个体产生干基萌苗,大叶栎(QuercusgriffithiiHk.) 的伐桩萌发也表现出相同的规律[23],这可能是因为随着树木直径和年龄的增大,树皮变得更加厚实和坚硬[23],树皮对休眠芽萌发的机械阻碍变大[20],因而休眠芽萌发的时间较晚,母树产生萌苗的数量也较少。
本研究中,环剥后第1年干基萌苗的枯死率为0.0%,而到了第2年,萌苗的枯死率为0.0%~41.7%,萌苗的数量则呈相反的趋势,即第1年单株母树产生的萌苗数量少于第2年。萌苗数量的增加导致了萌苗间的激烈竞争,发生了自疏,这与前人在栓皮栎、辽东栎伐桩萌苗中观测到的现象一致[10,25-26]。萌苗的产生和生长需要消耗大量营养物质[10,12],这些营养物质全部来源于母树根系,而环剥破坏了树干的韧皮部,阻碍了碳水化合物通过韧皮部维管束从叶向根的流通,减少了根系中淀粉的含量[27-28]。王文杰等[28]的研究显示,环剥后红松(PinuskoraiensisSiebold et Zuccarini)根中的可溶性糖的总量以及单种糖(葡萄糖、蔗糖和山梨糖)和淀粉含量均比对照树 (未环剥)低。但是,环剥没有破坏主干的木质部,根系还需要地上部分提供水分和营养物质[29-30],根系的呼吸和新根形成也需要动用根系储藏的碳水化合物[28]。然而,根系所储存的营养物质比较有限,环剥后根系所消耗的营养物质从地上部很少得到甚至无法得到补给,所以导致萌苗间对营养物质的激烈竞争。另外,主干的存在也造成了对干基萌苗的遮荫。干基萌苗得不到充足的光照、水分和营养物质,因此萌苗的枯死率增加。本研究在野外观察也发现,环剥后第2年,枯死的萌苗大部分为当年生萌苗,这说明休眠芽虽然能够萌发,但是在生长过程中由于养分、水分和光照供应不足,限制了能够存活的萌苗数量。
本研究中,胸径为≥10~<15 cm(2径级) 的母树上的干基萌苗在2个生长季中长势均优于其他径级母树的萌苗,主要是因为在环剥后第1年,该径级母树的萌苗数量少于其他径级,因此萌苗竞争不激烈,这一结果与薛瑶芹等[10]对栓皮栎伐桩萌发研究得到的萌苗数量与萌苗生长呈负相关的结论一致。环剥后第2年,该径级母树萌苗年龄结构的统计结果显示,2年生萌苗显著多于1年生幼苗,因此该径级母树上萌苗的平均高度、平均基径和平均冠幅均高于1年生萌苗所占比例较高的其他径级母树。另外值得关注的是,2径级母树的萌苗枯死率在环剥后2年内均为0.0%,而其他径级母树的萌苗至少有20%的个体枯死,笔者在野外监测时发现,在环剥后第2年,枯死萌苗中当年生萌苗占大部分,有的萌苗甚至是当年6月萌发,到了9月再调查时就已经枯死,另外个别2年生萌苗也没有成活,出现这种现象的原因可能是,根系储存的碳水化合物能够支撑新萌苗的产生,但是不能支持萌苗的生长,因为不成熟的幼苗在营养和水分竞争中处于劣势;而与此相反的是,2径级母树环剥后第2年产生的当年生萌苗较少,2年生萌苗全部成活,所以造成萌苗枯死率的差别。
本研究中,栓皮栎干基萌苗与母树的关系及其生长规律,与该树种的伐桩萌发规律有相似之处[7],但是环剥后干基萌苗的数量和萌苗生长远小于伐桩萌苗[7]。例如,本研究中每株母树能产生的干基萌苗为2~8个,2年生萌苗的高度为36.6~45.2 cm,而同一区域中栓皮栎伐桩萌苗的数量经过自然稀疏后为2~12个/桩, 2年生萌苗平均高度则为45.5~65.6 cm[7],出现这种现象的原因可能是伐桩上面没有遮荫的主干,伐桩根系的营养物质可全部供于萌苗生长;而产生干基萌苗的母树其主干和萌苗对根系营养物质的竞争较为激烈。虽然栓皮栎干基萌苗的数量和生长明显小于伐桩萌苗,但与其他树木一样,栓皮栎林即使没有受到大规模干扰,其仍然有可能遭受到人为小面积采伐、采样或者树倒、树枝掉落造成的树干损伤[13,15,31]。因此,栓皮栎的树干萌发能力对于个体存活必不可少,而且产生干基萌苗的母树主干被砍伐或者死亡后,被压迫的干基萌苗或许能迅速生长,从而保证母树的生态位不被其他树种占据。
本研究中,胸径为10~20 cm的栓皮栎母树在环剥后容易产生干基萌苗,且萌苗数量稳定,萌苗基径和高度也优于其他径级母树产生的萌苗,萌苗枯死率(0.0%~24.7%)小于其他径级母树。在栓皮栎次生林改造和经营过程中,可以提前1~2年在次生林中选择间伐对象,对胸径为10~20 cm的间伐目标以带宽为5 cm的标准环状剥去其基部离地面10 cm处的韧皮部,待其产生干基萌苗后再进行砍伐,能够缩短间伐林地土壤的裸露时间和促进林地的快速恢复。
[参考文献]
[1] Ky-Dembele C,Tigabu M,Bayala J,et al.The relative importance of different regeneration mechanisms in a selectively cut savanna-woodland in Burkina Faso,West Africa [J].Forest Ecology and Management,2007,243:28-38.
[2] Kammesheidt L.Forest recovery by root suckers and above-gr-ound sprouts after slash-and-burn agriculture,fire and logging in Paraguay and Venezuela Anglais [J].Journal of Tropical Ecology,1999,15:143-157.
[3] Simões C G,Marques M C M.The role of sprouts in the restoration of Atlantic Rainforest in Southern Brazil [J].Restoration Ecology,2007,15:53-59.
[4] 张文辉,卢志军.栓皮栎种群的生物学生态学特性和地理分布研究 [J].西北植物学报,2002,22(5):1093-1101.
Zhang W H,Lu Z J.A study on the biological and ecological property and geographical distribution ofQuercusvariabilispopulation [J].Acta Bot Boreal-Occident Sin,2002,22(5):1093-1101.(in Chinese)
[5] 韩照祥,山 仑.栓皮栎种群变异与适应对策研究 [J].林业科学,2005,41(6):16-22.
Han Z X,Shan L.Variation and adaptive countermeasures ofQuercusvariabilispopulation in Shaanxi Province [J].Scientia Silvae Sinicae,2005,41(6):16-22.(in Chinese)
[6] 吴明作,刘玉萃,姜志林.栓皮栎种群生殖生态与稳定性机制研究 [J].生态学报,2001,21(2):225-230.
Wu M Z,Liu Y C,Jiang Z L.The reproductive ecology and stable mechanism ofQuercusvariabilis(Fagaceae) population [J].Acta Ecologica Sinica,2001,21(2):225-230.(in Chinese)
[7] 薛瑶芹,张文辉,马莉薇,等.不同生境下栓皮栎伐桩萌苗的生长特征及在种群更新中的作用 [J].林业科学,2012,48(7):23-29.
Xue Y Q,Zhang W H,Ma L W,et al.The growth features ofQuercusvariabilisstump sprouts and its contribution to population regeneration in different habitats [J].Scientia Silvae Sinicae,2012,48(7):23-29.(in Chinese)
[8] 卢志军.栓皮栎种群生殖生态学研究 [D].陕西杨凌:西北农林科技大学,2002.
Lu Z J.Studies on reproductive ecology ofQuercusvariabilispopulation [D].Yangling,Shaanxi:Northwest A&F University,2002.(in Chinese)
[9] 高贤明,王 巍,杜晓军,等.北京山区辽东栎林的径级结构、种群起源及生态学意义 [J].植物生态学报,2001,25:673-678.
Gao X M,Wang W,Du X J,et al.Size structure,ecological significance and population origin ofQuercuswutaishanicaforest in Beijing Mountainous area [J].Acta Pkytoccdogica Sinica,2001,25:673-678.(in Chinese)
[10] 薛瑶芹,张文辉,周建云,等.秦岭南坡不同生境条件下栓皮栎伐桩萌芽特性 [J].林业科学,2011,47(7):57-64.
Xue Y Q,Zhang W H,Zhou J Y,et al.Sprout initiation and growth characteristics ofQuercusvariabilisstump after cutting in different habitats of the southern slope of Qinling Mountains,China [J].Scientia Silvae Sinicae,2011,47(7):57-64.(in Chinese)
[11] Kammesheidt L.The role of tree sprouts in the restoration of stand structure and species diversity in tropical moist forest after slash-and-burn agriculture in Eastern Paraguay [J].Plant Ecology,1998,139:155-165.
[12] 丁国华,程淑婉,叶镜中.杉木不同季节采伐伐桩萌芽的内源激素动态 [J].福建林学院学报,1996,16(2):109-113.
Ding G H,Cheng S W,Ye J Z.Dynamics of endogenous hormones in sprouting buds of Chinese Fir stump [J].Journal of Fujian College of Forestry,1996,16 (2):109-113.(in Chinese)
[13] Paciorek C J,Condit R,Hubbell S P,et al.The demographics of resprouting in tree and shrub species of a moist tropical forest [J].Journal of Ecology,2000,88:765-777.
[14] Del Tredici P.Sprouting in temperate trees:A morphological and ecological review [J].Botanical Review,2001,67:121-140.
[15] Mwavu E N,Witkowski E T F.Sprouting of woody species following cutting and tree-fall in a lowland semi-deciduous tropical rainforest,North-Western Uganda [J].Forest Ecology and Management,2008,255:982-992.
[16] Suh M H,Lee D K.Stand structure and regeneration ofQuercusmongolicaforests in Korea [J].Forest Ecology and Management,1998,106:27-34.
[17] Burns R M,Honkala B H.Silvics of North America:Vol.2 [M].Washington,U S:Forest Service Handbook,1990.
[18] 程淑婉,姜紫荣,杨 伦.杉木萌芽中内源细胞分裂素的分离鉴定 [J].林业科学,1987,23(1):79-84.
Cheng S W,Jiang Z R,Yang L.Isolation and identification of endogenous cytokinins in sprouts of Chinese Fir [J].Scientia Silvae Sinicae,1987,23(1):79-84.(in Chinese)
[19] 李景文,聂绍荃,安滨河.东北东部林区次生林主要阔叶树种的萌芽更新规律 [J].林业科学,2005,41(6):72-77.
Li J W,Nie S Q,An B H.Stump sprouting of the main broad-leaved tree species of secondary forest in eastern area of northeast China [J].Scientia Silvae Sinicae,2005,41(6):72-77.(in Chinese)
[20] Lockhart B R,Chambers J L.Cherrybark oak stump sprout survival and development five years following plantation thinning in the lower Mississippi alluvial valley,USA [J].New Forests,2007,33:183-192.
[21] Johansson T.Sprouting ability and biomass production of do-wny and silver birch stumps of different diameters [J].Biomass and Bioenergy,2008,32:944-951.
[22] 李景文,刘世英,王清海,等.三江平原低山丘陵区水曲柳无性更新研究 [J].植物研究,2000,20(2):215-220.
Li J W,Liu S Y,Wang Q H,et al.Study ofFraxinusmandshuricaasexual regeneration in the hills area of Sanjiang plain [J].Bulletin of Botanical Research,2000,20(2):215-220.(in Chinese)
[23] Khan M L,Tripathi R S.Tree regeneration in a disturbed sub-tropical wet hill forest of north-east India:Effect of stump diameter and height on sprouting of four tree species [J].Forest Ecology and Management,1986,17:199-209.
[24] Randall C K,Duryea M L,Vince S W,et al.Factors influencing stump sprouting by pondcypress (Taxodiumdistichumvar.nutans(Ait.) Sweet) [J].New Forests,2005,29:245-260.
[25] Rong L,Wenhui Z,Jingfeng H,et al.Survival and development of Liaodong oak stump sprouts in the Huanglong Mountains of China six years after three partial harvests [J].New Forests,2013,44(1):1-12.
[26] Xue Y Q,Zhang W H,Zhou J Y,et al.Effects of stump diameter,stump height,and cutting season onQuercusvariabilisstump sprouting [J].Scandinavian Journal of Forest Research,2013,28(3):223-231.
[27] Schneider H.Effect of trunk girdling on phloem of trunk of sweet orange trees on sour orange rootstocks [J].Hilgardia,1954,22:593-601.
[28] 王文杰,胡 英,王慧梅,等.环剥对红松(Pinuskoraiensis) 韧皮部和木质部碳水化合物的影响 [J].生态学报,2007,27(8):3472-3481.
Wang W J,Hu Y,Wang H M,et al.Effects of girdling on carbohydrates in the xylem wood and phloem bark of Korean pine (Pinuskoraiensis) [J].Acta Ecologica Sinica,2007,27(8):3472-3481.(in Chinese)
[29] Greene D W,Lord W J.Effect of dormant pruning,summer pr-uning,scoring and growth regulators on growth,yield and fruit quality of delicious and cortland apple trees [J].Journal of the American Society for Horticultural Science,1983,108:590-595.
[30] 李保国,王永蕙.增施氮肥和环剥对枣树光合速率的影响 [J].河北农业大学学报,1991,14(3):33-37.
Li B G,Wang Y H.Effects of some cultural methods on photosynthesis of Chinese jujube [J].Journal of Agricultural University of Hebei,1991,14(3):33-37.(in Chinese)
[31] Clark D B,Clark D A.The impact of physical damage on canopy tree regeneration in tropical rainforest [J].Journal of Ecology,1991,79:447-457.
