长江口北支水域营养盐的季节性变化
2014-03-25边佳胤吴惠仙薛俊增
吴 双, 边佳胤, 吴惠仙, 薛俊增
(1.上海海洋大学水产种质资源发掘与利用教育部重点实验室,上海 201306; 2.国家海洋局舟山海洋工作站,舟山 316000)
河口是淡水与海水交汇的地带,随时间推移,环流、潮汐进行常规混合过程,使得河口水文环境具有非恒定性及梯度型变化规律,复杂的环境特征保证了该区域内生物的多样性[1]。营养盐被认为是调节初级生产力生物量及种群结构的主要因素[2],在河口生态系统的物质循环与能量传递过程中起传递者作用[3]。已有证据表明河口区限制性营养盐随时间变化[4],且存在营养盐限制现象。Maestini等人研究表明Riga湾受河水影响大的区域呈现磷限制,河水影响小的区域呈现氮限制[5];在珠江口,浮游植物高值区东部,磷及硅相对溶解性无机氮含量较低,磷和硅可能是共同作用的限制因子[6];类似研究如英国Tweed河口,其冲淡水硝酸盐和硅酸盐季节变化显著[7]。
长江口年平均径流总量为9240亿m3[8],持续向河口及近海输送营养盐,为饵料生物生长提供了物质基础,进而促生舟山渔场等近海渔业资源集聚区。有研究表明盐度小于30 ppt的区域存在潜在磷限制[9],长江口同样存在该现象。然而,该区域又具特殊性,由于上游水利工程季节性调节[10],近岸洋流[11,12]等影响,长江口外及向南延伸的杭州湾区域已成为研究热点,有学者对不同时空条件下营养盐分布[3,13,14],营养盐形态特征[8],环境因子[15]以及水动力研究[16]进行调查。对于占径流1%[17]的长江口北支水域,仅见沉积物特征及河道演变等报道[18,19]。基于此,本文以2010年—2013年间在崇明岛北支河口调查资料为基础,对各站点营养盐组分及含量进行分析,并探讨其季节性变化特征以及与各环境因子间的关系,为紧邻该区域的东滩国际湿地及中华鲟保护区生态学研究提供基础数据支持,也为长江口北支水体中长期营养状态评价提出了参考依据。
1 材料与方法


图1 长江口北支采样点分布示意图
2 结果
2.1 溶解性无机氮的变化
2010年—2011年溶解性无机氮浓度在0.114~3.991 mg/L间变化(图2)。最低、最高值分别出现于春季(3月)和夏季(6月)。无机氮全年变化趋势呈单峰型,春季无机氮含量最低,在夏季出现高值,随之在秋冬两季逐渐下降。硝酸盐占DIN的比例为86%~98%,全年均值为90%,为DIN在四季的主要存在形态;亚硝氮所占比例为0.5%~8%,平均值为4%,为所有溶解性无机氮中含量最低的形态;氨氮所占比例为1%~13%,平均值为7%。
2012年—2013年,溶解性无机氮浓度在0.383~1.335 mg/L间变动。最低、最高值同样出现在春夏两季。全年无机氮构成中,主要成分硝酸盐所占比例为92%~97%,平均值为95%,高于之前年份;亚硝氮所占比例为1%~2%,全年平均值1%,对无机氮贡献最少;氨氮含量在2%~6%,全年平均值4%。两周年溶解性无机氮年平均值分别为1.24 mg/L和0.77 mg/L;另外溶解性无机氮在两周年中浓度略有下降。

图2 两年各季度间溶解性无机氮浓度变化
2.2 总磷浓度变化
2010年—2011年总磷浓度变化范围在0.09~1.70 mg/L之间(图3),全年平均浓度在0.56 mg/L。最高值出现在夏季,秋季总磷浓度迅速下降,与夏季相差1.6倍,为全年最低,枯水期(春冬两季)总磷浓度稳定在0.12~0.13 mg/L。

图3 两年各季度间总磷浓度变化
2012年—2013年,总磷浓度范围在0.22~0.78 mg/L之间变动。最高值在2012年9月,较前一年推迟一个季度。除最高值外,各季节趋于稳定。在次年3月份降至最低值,随2013年洪期到来,总磷呈回升趋势。2012年,受洪期延迟影响,总磷峰值出现延后,同期叶绿素值也达到最高值4.56 μg/L,该时段初级生产力含量上升,同时悬浮沉积物浓度上升,显著影响磷的相对吸附量[22],造成该季节总磷升高。
2.3 环境因子间的相关性
2010年—2013年,研究区域水体温度平均值为17℃,其变化范围为7.6~24.8℃,两年中均在9月达到最高值;盐度年度间变化较大,两年平均值分别为20.95 ppt和13.85 ppt;水体溶解氧两年均值分别为7.86 mg/L和9.36 mg/L,在2012年—2013年,各季度溶解氧含量均高于上一年同期测定值;在调查期间,pH值在7.30~8.50变动,平均值为8.08,属于偏碱性水,两年无明显差异;浊度平均值为357.6 NTU,2010年—2011年峰值829 NTU,2012年—2013年峰值1015 NTU,分别记录于2011年3月及2013年6月;总固体悬浮物均值为373.6 mg/L,与浊度相似,在调查期间呈现出逐年上升的趋势,两年均值分别为287.7 mg/L及459.5 mg/L,峰值与浊度出现时间一致,分别为642 mg/L及1005 mg/L,这两项均表明携带大量泥沙的长江上游来水过境时间在调查年份间有前后差,结合营养盐在两年中峰值出现时间,进一步说明长江洪期径流是夏秋季节营养盐的主要输入途径(图4)。
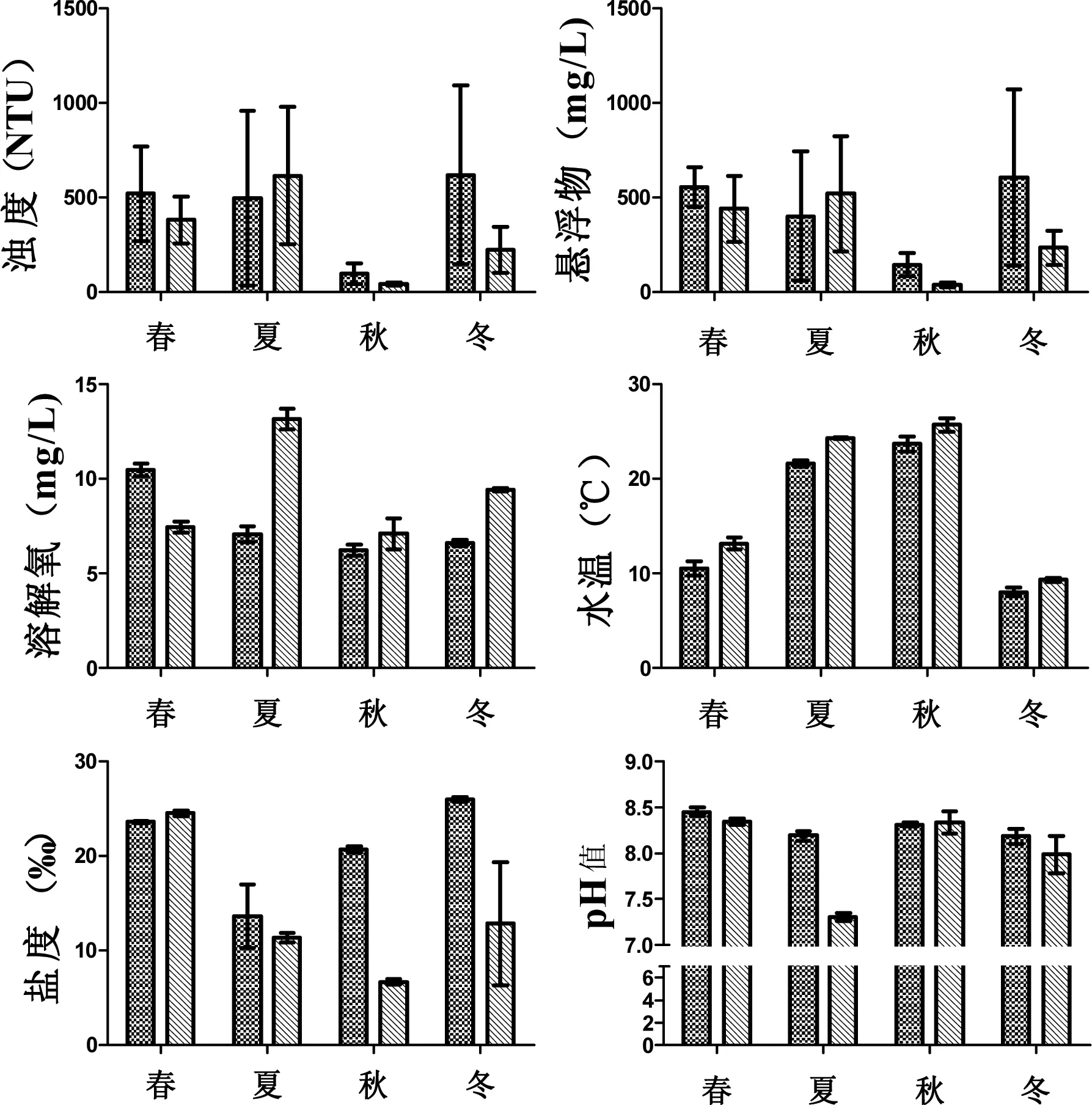
图4 两年中各季度环境因子变化

表1 不同环境因子间的Spearman相关系数(n=40)
*—在P<0.05时(双测),相关性显著;**—在P<0.01时(双测),相关性显著。
环境因子经SPSS 20.0标准化处理后进行Spearman相关性分析,结果显示:pH与盐度呈正相关,与叶绿素含量呈显著正相关,而与溶解性无机氮、总磷呈显著负相关;悬浮物与盐度、总磷呈显著正相关;总磷分别与温度、溶解性无机氮呈负相关、显著正相关见表1。
3 讨论
长江口北支溶解性无机氮浓度季节性变化显著,具体表现为夏季浓度远高于其余各季,全年变化趋势表现为单峰型,该现象在两年中趋势相似,具有稳定性。而特殊性体现在季节差异上,2011年夏季与2013年夏季溶解性无机氮浓度最大值相差3倍,远高于其他季节,可能由径流及大气沉降作用共同导致。结合各月降水量分布资料,发现该年长江口上游降水集中在6、7月,降水量高于其他季节4~7倍[23],在调查期内居高;径流受季节变化影响,且长江径流输入是口内无机氮高值来源[8,13,24],而北支流量仅占长江入海径流极少部分,因而水文变化对北支营养盐影响较南支更显著。盐度是影响营养盐平面分布的重要因素[25],枯季,北支河段受径流影响减小,口外冲淡水随潮流涌入,以物理混合主导的稀释过程促使溶解性无机氮含量进一步稳定,本研究中盐度与溶解性无机氮具显著负相关性,与已有研究一致[26]。北支硝酸盐约占溶解性无机氮总量的90%,占无机氮含量90%以上[25],从一方面反映了无机氮变化趋势。由表2可知,近年来硝酸盐组分均在90%左右,进一步证明无机氮分布具规律性。

表2 历年来无机氮的化学形态构成
总磷含量也呈现出季节性变化规律,即在夏秋之际浓度达到最低,冬春两季持平。然而两年间峰值出现时段不同。受2011年夏季强降雨过程影响[27],总磷含量提前进入上升期。研究证实总磷除受陆源输入外,生物活动及口外洋流等多过程控制[28],如悬浮颗粒物与水共同控制磷的吸附-释放过程[29],故总磷浓度在8月后含量大于5月[8],该现象与2012年—2013年度调查结果基本一致。
参考文献:
[1]梁 翠.长江口陆源输入变化及河口生态环境响应的初步研究[D].青岛:中国科学院海洋研究所,2013.
[2]Mallin M A, Cahoon L B, Mciver M R, et al. Alternation of factors limiting phytoplankton production in the cape Fear River Estuary[J]. Estuaries, 1999, 22(4): 825-836.
[3]李 磊,夏培艳,唐峰华,等.舟山附近海域富营养化的时空分布及其与环境因子的关系[J].生态学杂志,2011,30(4): 771-777.
[4]Fisher T R, Hagy III J D, Boynton W R, et al. Cultural eutrophication in the Choptank and Patuxent estuaries of Chesapeake Bay[J]. Limnol Oceanogr, 2006, 51(1): 435-447.
[5]Maestrini S Y, Berland B R, Bréret M, et al . Nutrients limiting the algal growth potential (AGP) in the Po River plume and an adjacent area, Northwest Adriatic Sea: Enrichment bioassays with the test algaeNitzschiaclosteriumandThalassiosirapseudonana[J]. Estuaries, 1997, 20(2): 416-429.
[6]Yin K, Qian P Y, Chenl J C, et al. Dynamics of nutrients and phytoplankton biomass in the Pearl River estuary and adjacent waters of Hong Kong during summer[J]. Marine Ecology Progress Series, 2000, 194: 295-305.
[7]Lohrenz S E,Fahnenstiel G L,Redalje D G,et al. Nutrients,irradiance,and mixing as factors regulating primary production in coastal waters impacted by the Mississippi River plume[J]. Continental Shelf Research, 1999,19: 1113-1141.
[8]全为民,沈新强,韩金娣,等.长江口及邻近海域氮、磷的形态特征及分布研究[J].海洋科学,2010,34(3):76-81.
[9]Chai C, Yu Z, Shen Z, et al. Nutrient characteristics in the Yangtze River Estuary and the adjacent East China Sea before and after impoundment of the Three Gorges Dam[J]. Science of The Total Environment, 2009,407(16): 4687-4695.
[10]Yang S L, Zhang J, Xu X J. Influence of the Three Gorges Dam on downstream delivery of sediment and its environmental implications, Yangtze River[J]. Geophysical Research Letters, 2007, D01:10.1029/2007GL029472.
[11]焦念志,王 荣,李超伦.东海春季初级生产力与新生产力的研究[J].海洋与湖沼,1998,29(2): 135-140.
[12]刘兴泉. 东海P、N 断面夏季温盐及化学要素的分布特征[J].海洋与湖沼,2001,32(2): 204-212.
[13]王 奎,陈建芳,金海燕,等.长江口及邻近海域营养盐四季分布特征[J].海洋学研究,2011,29(3): 18-35.
[14]李 峥,沈志良,周淑青,等.长江口及其邻近海域磷的分布变化特征[J].海洋科学,2007,31(1): 28-42.
[15]卢 勇,李洪亮,陈建芳,等.长江口及邻近海域表层水体溶解氧饱和度的季节变化和特征[J]. 海洋学研究,2011,29(3): 71-77.
[16]孙 霞,王宝栋,王修林,等.东海赤潮高发区营养盐时空分布特征及其控制要素[J].海洋科学,2004,28(8): 28-32.
[17]高 倩,徐兆礼,庄 平.长江口北港和北支浮游动物群落比较[J].应用生态学报,2008,19(9): 2049-2055.
[18]贾海林,刘苍宇,杨 欧.长江口北支沉积动力环境分析[J].华东师范大学学报:自然科学版,2001(1): 90-96.
[19]陈宝冲.长江口北支河势的变化与水、沙、盐的输移[J].地理科学,1993,13(4): 346-352.
[20]国家质量技术监督局.海洋监测规范[S].2007.
[21]宁修仁,孙 松,等.海湾生态系统观测方法[M].北京:中国环境科学出版社,2005.
[22]李 敏,韦鹤平,王光谦,等.长江口、杭州湾水域沉积物对吸附行为的研究[J].海洋学报,2004,26(1): 132-136.
[23]长江流域及西南诸河水资源公报[R].武汉:水利部长江水利委员会,2011.
[24]沈志良.长江和长江口氮的生物地球化学研究——关于长江N通量的研究[J].海洋科学,2002,26(12): 10-15.
[25]叶仙森,张 勇,项有堂.长江口海域营养盐的分布特征及其成因[J].海洋通报,2000,19(1): 89-92.
[26]李 磊,沈新强.春、夏季长江口海域营养盐的时空分布特征及营养结构分析[J].生态环境学报,2010, 19(12): 2941-2947.
[27]王新才.长江流域及西南诸河水资源公报[Z].湖北:长江出版社, 2012.
[28]黄自强,暨卫东.长江口水中总磷、有机磷、磷酸盐的变化特征及相互关系[J].海洋学报,1994,16(1): 51-60.
[29]Filipelli G M, Delaney M L. Phosphorus geochemistry of equatorial Pacific sediments[J].Geochim Cosmochim Acta, 1996, 60: 1479-1495.
