长江口北支大型底栖动物群落周年变化特征
2014-03-25赵开彬吴惠仙边佳胤王宝强薛俊增
赵开彬, 刘 婧, 吴惠仙, 边佳胤, 王宝强, 薛俊增
(上海海洋大学 水产种质资源发掘与利用教育部重点实验室, 上海 201306)
底栖动物群落是河口生态系统中重要的组成部分,影响着河口底栖生态系统的生态过程[1],河口底栖动物群落主要有多毛类、软体动物、甲壳动物和脊索动物,其中多毛类、软体动物和甲壳动物最为常见[2-4],季节、水文、环境以及人类活动等影响着河口底栖动物的生态特征[ 5-8]。
河口作为河流与海洋的交汇区域,其生态环境独特,生物群落有其特有的生态特点[9],大型底栖动物由于运动能力差和群落相对稳定的特点,成为河口生态研究中的重要研究对象[ 10],在河口底栖动物的时空分布[ 11-14]、群落特征[ 15-17]、丰度和生物量[18]等方面都有相关报道。长江口是中国最大的河口,中国在20世纪50年代末就开始长江口底栖动物研究,在长江口底栖生物的种类组成[19, 20]、群落结构[20, 22]、次级生产力[21]等方面开展了许多工作,有关长江口底栖动物的报道大都集中在长江口南支和口外海域[6, 7, 9-14, 21-23],北支底栖动物的研究至今报道不多[11],对北支周年变化特征分析以及与环境因子的关系少有报道,长江口北支受海水影响较大[11],盐度在季节水平上有较大差异,且含盐度有所降低[24],有大片淤积滩涂,底栖动物丰富。本文在长江口北支进行了周年大型底栖动物群落生态学研究,一方面阐明长江口北支大型底栖动物群落结构和多样性的生态现状,另一方面探讨流域内人类活动对该区域底栖动物的影响,以期为长江口北支的生态环境保护与修复、长江口湿地以及中华鲟等水生生物资源的保护提供理论基础。
1 材料与方法
2010年10月至2011年9月于崇明北水闸至黄瓜沙下口的崇明北滩沿岸,设5个采样点,调查水域位于31°37.008′~31°769′N与121°46.633′~121°50.733′E(图1),依据《海洋调查规范海洋生物调查》[25],每月中旬选取大潮高潮的平潮期进行大型底栖动物样本拖网采集和野外水化学指标现场测定工作。

图1大型底栖动物采样点示意图
利用经过1 m口宽网兜孔径0.5 cm的改良阿氏拖网,在每个样点以进行约15 min的低速(2 kn)拖网采样。起网后现场抽取海水洗涤样本,过筛(孔径0.5 mm)清洗装罐。现场按照软体动物、环节动物、脊索动物、节肢动物和其它门类进行初步分拣,根据大型底栖动物不同类型各自特点分别添加5%的福尔马林或75%的酒精进行样本固定。采样结束后将样本带回实验室统计各种类数量和称重后依据样本个体大小等分别装瓶密封、标记并保存[14]。
水质样本采样依据《海洋监测规范》进行。悬浮物含量和浊度的测定根据《水环境监测标准分析方法》[26]相关内容进行,温度、盐度和溶解氧含量使用YSI 85型号的手提式野外水质检测仪在采样现场进行测定。
数据分析均在 R 2.15.1[27]中进行,使用 VEGAN 程序包计算长江口大型底栖动物群聚的 Margalef 种丰富度指数 (D) 、Shannon-Wiener 信 息 指 数 ( H′) 、Pielou 均匀度指数( J),利用长江口北支5个样点的密度和环境因子进行RDA分析。其它数据参考公式如下:
式中N表示各采样点所有物种个体总数,ni代表第i种的个体总数,fi表示该物种在各个采样点出现的频率,当Y>0.02时,该物种为群落中的优势种。
2 结果
2.1 环境因子
本次研究分析了盐度、水温、溶解氧含量、悬浮物含量和浊度共5个环境因子周年变化具体结果见图2。

图2 长江口北支水域环境因子周年变化
盐度周年均值维持在18.48,冬季盐度尤高,均值为24.26;夏季最低,均值为12.49;春、秋季节盐度均值分别为21.22和15.96。
周年水温均值为16.29℃,全年季节变化明显,夏季均温最高,为25.98℃;冬季最低为5.39℃;春季14.69℃明显高于冬季均温,秋季水温均值19.12℃略高于春季。
水体溶解氧含量周年平均水平在7.00 mg/L上下变动,春冬季节的溶解氧含量和夏秋季节溶解氧含量分别较为接近,其均值分别为7.50 mg/L、7.45 mg/L、6.34 mg/L和6.72 mg/L。
悬浮物年平均值为841.60 mg/L,春季悬浮物浓度最高,为1289.31 mg/L;其次为夏季的804.30 mg/L;冬季略低于夏季为768.88 mg/L;秋季最低,其均值为503.93 mg/L。
浊度周年变化波动较大,年平均值为871.62 NTU;春季水体浊度最高,为1362.18 NTU,夏季次之为931.66 NTU,秋季浊度最低,为498.10 NTU;冬季比秋季略高,为694.56 NTU。
2.2 种类组成
共采集到大型底栖动物78种,其中软体动物23种、环节动物7种、节肢动物28种、脊索动物18种和其它种类2种(紐虫、拟细浅室水母各1种),具体见表1。全年除冬季节肢动物占比较大外,其它季节软体动物占比均较大(图3)。
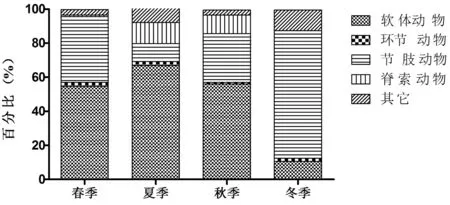
图3 周年各季节种类占比
春季(3月—5月)底栖动物种类组成从高到低依次为软体动物(占比54.67%)、节肢动物(占比38.50%)、其它种类(占比3.29%)、环节动物(占比2.38%)、脊索动物(占比0.98%)。
夏季(6月—8月)底栖动物种类组成由高到低依次为软体动物(占比66.89%),脊索动物(占比12.45%),节肢动物(占比10.41%),其它种类(占比7.88%),环节动物(占比2.38%)。
秋季(9月—11月)底栖动物种类组成由高到低依次为软体动物(占比56.13%),节肢动物(占比28.63%),脊索动物(占比10.80%),其它种类(占比3.01%) ,环节动物(占比0.88%)。
冬季(12月—翌年2月)底栖动物种类组成由高到低依次为节肢动物(占比75%),其它种类(占比12.17%) ,软体动物(占比10.60%),环节动物(占比1.77%)。
2.3 大型底栖动物周年优势种
长江口北支水域大型底栖动物群落全年优势种(Y>0.02)共有8种(表2),优势度最高的类群为软体动物,节肢动物次之,脊索动物最少。

表1长江口北支大型底栖动物的种类组成及分布

续表1(Table 1 Continued)

表2 长江口北支水域大型底栖动物优势种
2.4 大型底栖动物周年密度变化
大型底栖动物栖息密度季节变化明显(图4),年均值为0.45 ind/m2,5月至8月密度相对较大,5月密度最大,为1.24 ind/m2,7月次之,为1.10 ind/m2,1月密度最少,为0.08 ind/m2,各样点密度相差不大,S3密度相对较大,为1.38 ind/m2,其次S1密度较大,为1.28 ind/m2, S2密度最少,为0.80 ind/m2。

图4 大型底栖动物密度的周年变化
2.5 大型底栖动物周年生物量变化
大型底栖动物栖息生物量季节变化明显(图5),变化趋势和密度基本一致,5月至8月生物量相对较大,5月密度最大,为1.16 g/m2,7月次之,为0.93 g/m2,2月生物量最少,为0.01 g/m2,各样点生物量稍有变化,S1和S3生物量最大,为0.64 g/m2, S5生物量最少,为0.17 g/m2。

图5 长江口北支各站位大型底栖动物生物量的周年变化
2.6 大型底栖动物周年生物多样性变化
长江口北支大型底栖动物群落物种多样性在各个采样站点间的分布特征各有不同(表3)。Margalef丰富度指数(D)的高峰值在样点S1,S2最低。Pielou均匀度指数(J)的变化范围在0.68至0.75之间,S2和S5的均匀度指数同为最高值,S3的均匀度指数最低。 采样站点S5的Shannon-Wiener多样性指数(H′)最高,数值是2.86,S4样点的Shannon-Wiener多样性指数数值最低,为2.43。
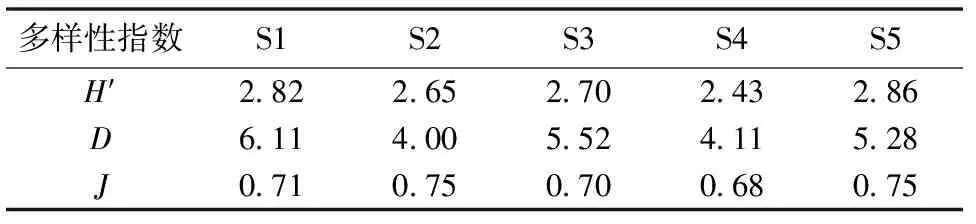
表3 长江口北支大型底栖动物群落多样性站位分布
全年12个月中(表4),Shannon-Wiener多样性指数(H′)的变化规律为秋季10月份最高,8月份仅次于10月份;次高峰出现在冬季的12月份;最低值为冬季的2月份,次低值在9月份出现。Pielou均匀度指数(J)最高值和Shannon-Wiener多样性指数分布不一致,出现在冬季的1月份,最低值在7月份,次高峰在秋季的10月份出现。Margalef丰富度指数(D)的分布规律和前两个多样性指数有所不同,峰值在夏季的7月份,次高值在夏季的6月份和秋季的10月份,这两个月份丰富度指数一样,最低值在冬季的1月份出现。

表4 长江口北支大型底栖动物群落多样性月度分布
2.7 环境因子与各月大型底栖动物密度相关性分析和蒙特卡罗置换检验
长江口北支水域各月出现的大型底栖动物栖息密度与溶氧量、浊度、悬浮物、盐度和温度进行RDA相关性分析。如图6所示,溶氧量、浊度、悬浮物、盐度和温度对长江口北支水域各月大型底栖动物栖息密度差异性显著,1月、2月、3月、4月、8月、9月、11月和12月与盐度、溶氧量、悬浮物、浊度成正相关,与温度成负相关;5月、6月、7月与温度成正相关;6月与温度相关性最强。对各月大型底栖动物栖息密度和环境因子进行蒙特卡罗置换检验,结果如表5所示,从全年来看,盐度(r2=0.7190,P<0.01)和温度(r2=0.8291,P<0.01)是影响长江口北支大型底栖动物群落分布的主要环境因子。

图6 长江口北支各月大型底栖动物栖息密度与环境因子RDA排序图

表 5长江口北支各月大型底栖动物栖息密度与环境因子蒙特卡罗置换检验
Signif. codes:‘***’, 0.001, ‘**’, 0.01, ‘*’, 0.05;Pvalues based on 999 permutations.
3 讨论
国内外河口盐度对大型底栖动物的影响[2, 3, 28-36]主要为群落结构组成和优势种变化,长江口北支盐度对底栖动物的研究较少,调查区域主要在口外海域,且为群落结构变化,本文通过一周年的调查研究发现,长江口北支大型底栖动物的生态特点为群落结构受盐度(r2=0.7190,P<0.01)、酸碱度(r2=0.6977,P<0.01)和温度(r2=0.8291,P<0.01)影响显著,盐度季节变化明显,冬季较高,春季有所下降,夏季较低,秋季开始回升,这和章飞军等人[23]对盐度变化影响相同,研究区域也相对一致,而田伟等人对椒江口的研究[35]以及寿鹿等人对瓯江口的研究[32]认为盐度和底栖动物密度并无显著相关关系,这与长江口北支位于口内,受上游冲淡水影响,盐度变化剧烈,而椒江口和瓯江口海域受上游来水影响较小、盐度分布比较均匀有关;相比罗民波等人[11]的研究种类数增加近5倍,密度和生物量相对较少,这与北支生态修复[13]有一定影响,改善了北支水环境,也可能与来自上游来水的减少导致盐度增加、高盐水种增多有关,从优势种的变化[4-6,9-14]来看,北支底栖动物生活环境处于动态变化过程,持久的监测是对北支水环境保护和利用开发的必要前提。
温度对于河口底栖动物的研究一般表现在季节的差异上,四季温差较大,不同底栖动物受温度变化,表现出不同生长繁殖水平,底层水温低将导致底栖动物种类数降低,栖息密度较低[10],长江口水体较浅,底温受季节变化影响明显,冬季大型底栖动物密度远低于其它3季,且只有夏季的1/4,多样性指数(Shannon-Wiener)低于其它3季,丰富度指数(Margalef)只有夏季的1/2,低于其它3季,均匀度(Pielou)基本相差不大,种类数、密度和生物量在水温较高的春季、夏季和秋季明显高于水温较低的冬季,而水温是影响软体动物分布的重要因子[34],本次调查软体动物种类数、密度和分布随季节变化非常明显,夏季水温较高,软体动物占总体比例66.89%,远大于冬季10.60%,密度和生物量也显著升高,广泛分布于各站位,这和彭松耀等人对珠江口的研究[31]、寿鹿等人对瓯江口的研究[32]底栖动物受温度变化规律基本一致,而王晓晨等人对黄河口的研究[37]认为温度对大型底栖动物种类组成的相关性不是很明显,这可能与研究区域有关。本文研究区域主要位于长江口北支口内海域,而王晓晨等人对黄河口的研究主要分布在口外海域,口外海域温度随季节变化不是很明显,这在张敬怀等人对珠江口的研究[33]中也得到支持。
参考文献:
[1]叶属峰,纪焕红,曹 恋,等. 河口大型工程对长江河口底栖动物种类组成及生物量的影响研究[J]. 海洋通报,2004,23(4):32-37.
[2]Thompson B, Ranasinghe J A, Lowe S, et al. Benthic macrofaunal assemblages of the San Francisco Estuary and Delta,USA[J]. Environ Monit Assess,2013, 185: 2281-2295.
[3]Reinicke G B. Small-scale patchiness of benthos and sediment parameters in a freshwater tidal mud-flat of the river Elbe Estuary (Germany)[J]. Limnologica, 2000,30: 183-192.
[4]陈 亚, 瞿 安, 赵 林. 长江口九段沙潮间带大型底栖动物季节分布特征的初步研究[J]. 水产学报,2007,31(1): 52-58.
[5]单秀娟, 线薇薇, 武云飞. 长江河口生态系统鱼类浮游生物生态学研究进展[J]. 海洋湖沼通报,2004,4: 87-93.
[6]方 涛, 李道季, 李茂田,等. 长江口崇明东滩底栖动物在不同类型沉积物的分布及季节性变化[J]. 海洋环境科学,2006,25(1): 24-27.
[7]钱小娟, 陈 艳, 顾爱辉. 长江口北支段纳污能力分析[J]. 水资源保护,2012,28(3): 64-67.
[8]郭丰义. 美国海洋生物调查研究的现状和趋向[J]. 海洋科技资料. 1978,3: 89-96.
[9]刘录三,孟 伟,田自强,等. 长江口及毗邻海域大型底栖动物的空间分布及历史演变[J].生态学报,2008,28(7): 3027-3034.
[10]刘录三,郑丙辉,李宝泉,等. 长江口大型底栖动物群落的演变过程及原因探讨[J]. 海洋学报,2012,34(3): 135-145.
[11]罗民波,沈新强,徐兆礼,等. 长江口北支水域潮间带大型底栖动物研究[J]. 海洋环境科学,2006,25(4): 43-47.
[12]罗祖德. 长江口是个稀世之宝[J]. 自然与科技,2011,183: 40-43.
[13]沈新强,陈亚瞿,罗民波. 长江口底栖生物修复的初步研究[J]. 农业环境科学学报, 2006,25(2):373-376.
[14]王金辉,黄秀清,刘阿成,等. 长江口及邻近水域生物多样性变化趋势分析[J]. 海洋通报,2004,23(1): 32-39.
[15]徐长乐. 建设海上人工岛及长江口亚三角洲体系的战略意义[J]. 江南论坛,2011:20-22.
[16]叶属峰,纪焕红,曹 恋,等. 河口大型工程对长江河口底栖动物种类组成及生物量的影响研究[J]. 海洋通报,2004,23(4): 32-37.
[17]陈 炜,李九发,李占海,等. 长江口北支强潮河道悬沙运动及输移机制[J]. 海洋学报(中文版),2012,34(2): 84-91.
[18]朱晓君,陆健健. 长江口九段沙潮间带底栖动物的功能群[J]. 动物学研究,2003,24(5): 355-361.
[19]王小谷,王春生,张东声,等. 长江口及其陆架春季小型底栖生物丰度和生物量[J]. 生态学报,2010,30(17): 4717-4727.
[20]吕巍巍,马长安,余 骥,等. 围垦对长江口横沙东滩大型底栖动物群落的影响[J]. 海洋与湖沼,2012,43(2):340-347.
[21]严 娟,庄 平,侯俊利,等. 长江口潮间带大型底栖动物次级生产力及其影响因子[J]. 应用与环境生物学报,2012,18(6): 935-942.
[22]李宝泉,李新正,王洪法,等. 长江口附近海域大型底栖动物群落特征[J]. 动物学报,2007,53 (1) :76-82 .
[23]章飞军,童春富,张 衡,等. 长江口潮下带春季大型底栖动物的群落结构[J]. 动物学研究,2007,28 ( 1) : 47-52.
[24]李伯昌,余文畴,陈 鹏,等. 长江口北支近期水流泥沙输移及含盐度的变化特性[J]. 水资源保护,2011,27(4):31-34.
[25]GB/T12763.2-1991 Specifications for oceanographic survey [Z]. 1991. [GB/T 12763. 2-1991海洋调查规范[Z]. 1991.]
[26]薛巧英. 水环境质量评价方法的比较分析[J]. 环境科学保护,2004,30(124):64-67.
[27]R Development Core Team R D S. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, 2010. Vienna, Austria. Available at: http://www.R-project.org.
[28]Teske P R, Wooldridge T H. What limits the distribution of subtidal macrobenthos in permanently open and temporarily open/closed South African estuaries Salinity vs. sediment particle size [J]. Estuarine, Coastal and Shelf Science, 2003, 57 :225 238.
[29]Beauchard O, Jacobs S, Ysebaert T, et al. Sediment macroinvertebrate community functioning in impacted and newly-created tidal freshwater habitats[J]. Estuarine, Coastal and Shelf Science , 2013,120 : 21-32.
[30]Martins I, Oliveira J M, Flindt M R, et al. The effect of salinity on the growth rate of the macroalgaeEnteromorphaintestinalis(Chlorophyta) in the Mondego estuary (west Portugal)[J]. Acta Oecologica , 1999,20 (4) :259-265.
[31]彭松耀,赖子尼,蒋万祥,等. 珠江口大型底栖动物的群落结构及影响因子研究[J]. 水生生物学报, 2010,34(6):1179-1189.
[32]寿 鹿,曾江宁,廖一波,等. 瓯江口海域大型底栖动物分布及其与环境的关系[J]. 应用生态学报,2009,20(8):1958-1964.
[33]张敬怀,高 阳,方宏达, 等. 珠江口大型底栖生物群落生态特征[J]. 生态学报,2009,29(6):2989-2999.
[34]王国强. 软体动物胚胎发育的温度效应研究进展[J]. 安徽农业科学,2012,40( 28):13830-13831.
[35]田 伟,徐兆礼,周 进,等. 椒江口海域大型底栖动物群落结构的研究[J]. 复旦学报:自然科学版,2013,52(2):271-277.
[36]Xu H Z, Lin J , Wang D X. Numerical study on salinity stratification in the Pamlico River Estuary[J]. Estuarine, Coastal and Shelf Science ,2008,80:74-84.
[37]王晓晨,李新正,王洪法. 黄河口岔尖岛、大口河岛和望子岛潮间带秋季大型底栖动物生态学调查[J]. 动物学杂志,2008 ,43 (6) :77-82.
