青海高寒区不同人工林配置下灌木种群的生态位研究
2014-03-25杨海龙贺康宁王伟伟高二鹏刘慧博王冰洁
曹 恒,杨海龙,贺康宁,王伟伟,高二鹏,刘慧博,王冰洁
(北京林业大学 水土保持学院,北京 100083)
生态位是指群落内一个种群在时间、空间上的位置以及种群在群落中的机能关系[1-2]。最初,生态位主要应用于动物生态学方面,随着对生态位研究的不断深入,该理论在种间关系、动态演替、物种进化及城市规划、植物保护、经济评价等方面都得到了广泛应用[3-6]。目前,对生态位宽度(Niche breadth)和生态位重叠(Niche overlap)的定量计测是生态位理论研究中一个非常重要的内容[7]。
迄今为止,对不同人工林配置下灌木种群之间的相互关系、各种群的功能和地位及其是否会对人工林群落的结构产生影响等问题,并没有得到应有的关注,而这些问题可能会对人工林生物多样性的正确理解产生影响[8]。本研究将运用生态位理论,对青海大通高寒区不同人工林下灌木物种主要种群的生态位宽度和生态位重叠进行定量研究[9],旨在了解灌木层种群对环境资源的利用程度和相互关系,以及种群在群落中的功能和地位,为评价人工林群落结构的稳定性和物种多样性及解释该区域不同人工林下灌木物种共存和竞争的机制等提供依据,并为当地人工林群落结构的不断优化和可持续经营管理提供理论依据。
1 研究区概况
研究区位于青海省大通县宝库林场(北纬36°55′~42°36′,东经100°52′~101°39′),气候属半干旱、半湿润温良性气候。年平均气温2.4 ℃,极端最高气温30.9 ℃,极端最低气温-33.1 ℃。年平均降雨量549.9 mm,年内无霜期70~120 d,年气温日较差平均为14.7 ℃,年湿润指数为0.56~1.32,平均风速为2 m/s。土壤为山地棕褐土,平均森林覆盖率为79.0%。主要造林乔木树种有青海云杉(Piceacrassifolia)、白桦(Betulaplatyphylla)、华北落叶松(Larixprincipis-rupprechtii)等,主要灌木种有杜鹃(Rhododendronsimsii)、沙棘(Hippophaerhamnoides)、金露梅(Potentillafruticosa)、山生柳(Salixoritrepha)、小蘖(Berberiscircumsterrata)等,主要草本物种有细叶绣线菊(Spiraeasalicifolia)、窄叶鲜卑花(Sibiraeaangustata)、高山柳(SalixcupularisRehd)、峨眉蔷薇(RosaomeiensisRolfe)、刚毛忍冬(LonicerahispidaPall)等。
2 研究方法
2.1 野外调查
在对青海省大通县宝库林场不同人工林进行实地踏勘的基础上,选取具有代表性的青海云杉林、白桦林、华北落叶松林和云杉-白桦混交林4种人工林进行植被样方调查,并测取相应的海拔、坡度、坡位、坡向等生态因子(表1),每种林分设置4个5 m×5 m的灌木样方,并调查各灌木的株数、株高、地径和冠幅等指标。
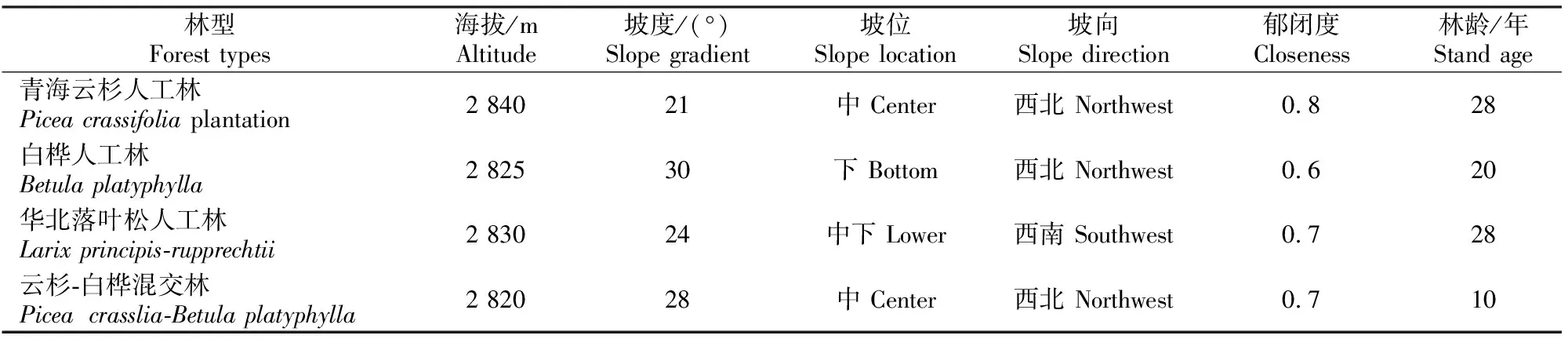
表1 青海大通高寒区4种不同人工林群落的基本情况
2.2 指标计算方法
2.2.1 重要值 重要值作为度量植物种群的重要指标,综合反映了植物物种在群落中的重要程度。其计算公式为:
重要值=(相对密度+相对频度+相对显著度)/3。
2.2.2 生态位宽度 生态位宽度有多种计算方法,本研究采用较为通用的 Levins生态位宽度[10]计算公式,即:
式中:BLi代表物种的生态位宽度;Pij代表物种i对第j资源的利用占其对全部资源利用的百分率,Pij=nij/Ni,其中nij为种i在样方j中的重要值,Ni=∑nij;r代表样方数。
2.2.3 生态位重叠度 生态位重叠度采用 Pianka生态位重叠指数[11]公式计算:
式中:Oik为物种i与物种k的重叠度指数,nij和nkj为种i和种k在样方j中的重要值,r为样方数。
3 结果与分析
3.1 不同人工林群落下灌木层种群的生态位宽度
生态位宽度可以用来衡量一个物种对资源的利用状况和对所处环境的适应情况,一个物种的生态位宽度越大,说明该物种在该区域分布越广,对所处环境的适应能力越强,同时具有更强的资源竞争能力[12]。4种人工林群落下各灌木层种群的生态位宽度如表2所示。

表2 青海大通高寒区不同人工林下灌木层种群的生态位宽度
由表2可见,在白桦人工林群落中,高山柳和八宝卫矛的生态位宽度最大,均为0.999,说明其在该群落中可利用的资源最为丰富且利用资源的能力很强,在该生境范围内对诸如水分、热量、土壤等生态因子无特殊要求,从而成为生态幅度最广的种群。在云杉-白桦混交林群落中,生态位宽度排在前边的依次是金露梅>高山柳>窄叶鲜卑花>细叶绣线菊,其生态位宽度值为0.973~0.994,表明金露梅、高山柳、窄叶鲜卑花和细叶绣线菊对所处环境的适应能力较强,具有较强的资源竞争能力,能利用群落环境中的绝大多数资源;而蓝果忍冬(Loniceracaerulea)、峨眉蔷薇、水荀子(CotoneasterhorizontalisDecne)等物种的生态位宽度均在0.4以下,表明其在群落中处于劣势地位,该群落环境不太适宜这些种群生长,或者意味着其在该群落中的地位是暂时的和次要的,随着群落的不断演替,可能将被其他种群逐渐取代。在青海云杉人工林下,八宝卫矛是该群落灌木层的优势种,在创造群落内部独特环境中发挥着重要作用,且分布广、数量多,利用资源较为充分,具有较大的生态适应范围,作为耐阴树种,在群落郁闭度较高的林冠下仍然能够进行更新和演替,故其生态位宽度最大[13]。在华北落叶松人工林中,生态位宽度表现为峨眉蔷薇>锦鸡儿>八宝卫矛,其生态位宽度值依次为0.986,0.933和0.295,可见峨眉蔷薇和锦鸡儿是该人工林下灌木层的优势种。
在这些人工林群落中,峨眉蔷薇和八宝卫矛均有出现,为不同人工林下的优势灌木种,说明其在不同人工林下对环境的利用较为宽泛,但峨眉蔷薇在云杉-白桦混交林中的生态位宽度较低,仅有0.296,而在纯林中的生态位宽度均在0.890以上,说明峨眉蔷薇更适合在纯林中生长,在该群落中有更加充足的资源供其利用。
由表2可见,各纯林之间平均生态位宽度最大的为白桦人工林,平均值达0.798,这与白桦林林分稀疏、郁闭度小、光照强、通风条件好、土壤腐殖质含量高密切相关;同时也表明在各纯林中,白桦人工林下灌木层种群对资源的利用率较大,生长较好;与纯林相比,云杉-白桦混交林下灌木层的平均生态位宽度最小,仅为0.639,表明在青海高寒区,混交人工林群落并不利于林下灌木种群的生长和发育。
3.2 不同人工林群落下灌木层种群的生态位重叠度
生态位重叠体现了种群之间对于相同等级资源利用的相似性程度和在空间上的配置关系[14],不同植物群落在系统结构、物种功能及组成等方面都是不一样的,这也造成其林下灌木层在生态位重叠度方面相差较大。因此,本研究对比分析了白桦和青海云杉人工林与二者混交林(云杉-白桦混交林)群落下灌木层种群的生态位重叠度,结果见表3。由表3可见,在3种人工林群落下,灌木种群生态位重叠度大于0.600的物种百分率依次为33.3%,42.2% 和16.7%,表现为针阔叶混交林下灌木种群的生态位重叠度最高。
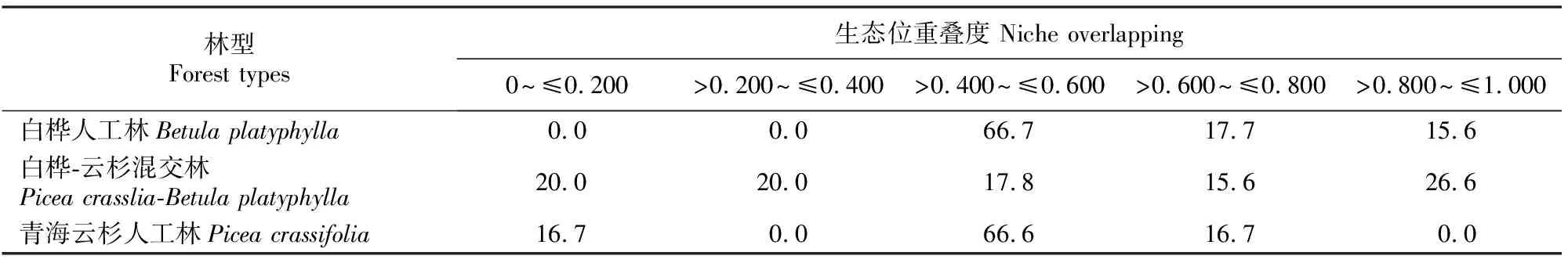
表3 青海大通高寒区不同人工林下灌木层种群在各生态位重叠度的分配比例
由表4可见,生态位重叠度平均值和生态位宽度平均值均以白桦人工林最大,其次为青海云杉人工林,最小的为云杉-白桦混交林。该结果表明,纯林下物种更加丰富,灌木层物种对环境的适应范围广,对资源的利用更为充分,相应纯林下灌木层物种间的竞争也更为激烈,可最大限度地利用了周边资源,在物种对环境的不断适应和物种间相互激烈竞争的双重作用下,群落中的物种相互适应和协同进化,在形态、结构和生理特性方面出现了许多互补的特征,使群落自身结构不断趋于稳定,物种多样性不断丰富,从而出现了物种与环境和谐统一的局面[15]。
由表5可见,在白桦人工林群落中,各种群之间的生态位重叠度相对较高,绝大多数在0.500~1.000 波动,根据生态位重叠的定义(具有相同生物生态学特性的物种之间在与其相联系的生态因子上具有相似性),表明该群落中的绝大多数种群对生态因子有相似的要求[16],特别是高山柳,其与八宝卫矛、细叶绣线菊、金露梅的生态位重叠度均在0.800以上,这是因为高山柳分布广,对资源的利用能力强,与其他种的分布地段重叠大,有较大的生态位宽度,这与前面的计算结果一致;水荀子与密枝柳、红花忍冬的生态位重叠度也较大,均在0.900以上,但其生态位宽度却均在0.52以下,这可能是由于其生物生态学习性基本一致,对资源环境的要求相同,为了生长发育,在局部生境条件下,有限种群对仅有环境资源(光照、水分等)激烈竞争的结果。
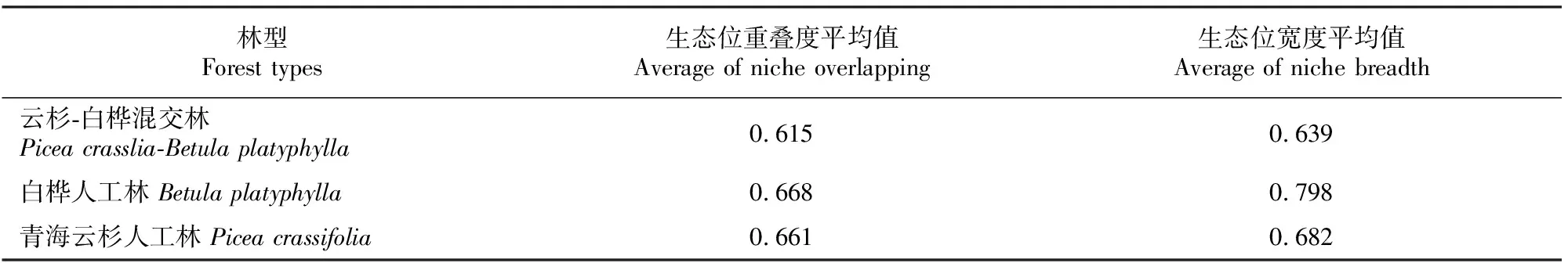
表4 青海大通高寒区不同人工林下灌木层生态位重叠度及生态位宽度的平均值

表5 青海大通高寒区白桦人工林下灌木层种群的生态位重叠度
由表6可见,在云杉-白桦混交林下,白桦幼苗和锦鸡儿与八宝卫矛、水荀子、峨眉蔷薇、蓝果忍冬的生态位重叠度均为0,表明在该群落中,这些种群的生物生态学习性及其对生境因子的要求均不相同,不具有共同的资源位;金露梅、高山柳、窄叶鲜卑花、细叶绣线菊之间的生态位重叠度均较低,为 0.400~0.600,但其生态位宽度均较高;峨眉蔷薇、蓝果忍冬、水荀子等的生态位宽度较小,但生态位重叠度较大,均在0.890以上。这表明物种间的生态位宽度与其生态位重叠度大小之间并无明显的正相关关系,主要是因为生态位宽度反映的是物种对资源的利用状况和对所处环境的适应情况,生态位宽的物种其生物生态学特性并不一定相同,在对资源的要求上就不一定完全一致,从而导致其生态位重叠度不一定均较高[17]。

表6 青海大通高寒区云杉-白桦混交林下灌木层物种的生态位重叠度
4 结论及建议
生态位宽度的大小反映了种群对资源利用程度的高低及其对环境适应能力的强弱。本研究中,不同人工林下相同灌木物种的生态位宽度也不同,表明不同人工林下灌木种群的广生态幅物种存在较大的差异,同一人工林下不同灌木物种的地位也存在着较大的差异,如在云杉-白桦混交林群落中,金露梅、高山柳、窄叶鲜卑花和细叶绣线菊处于优势地位,而蓝果忍冬、峨眉蔷薇、水荀子等处于劣势地位,不同人工林的立地条件和不同物种的生物生态学特性共同决定了这一现象。
从不同人工林下灌木物种的平均生态位宽度上看,纯林之间平均生态位宽度以白桦人工林最大,其次为华北落叶松人工林,最小的为青海云杉人工林,表明在各纯林中,白桦人工林下灌木层种群对资源的利用率较好,更适应所处环境,这与白桦林林分稀疏、郁闭度小、光照强、通风条件好、土壤腐殖质含量高密切相关。该研究结果表明,群落生境范围内的热量、水分、土壤等生态因子是多物种并存的人工林配置模式筛选的原因和驱动机制之一;与纯林相比,云杉-白桦混交林下灌木层的平均生态位宽度最小,表明在青海高寒区,混交人工林群落并不利于林下灌木种群的生长和发育,相反纯林更有利于促进林下灌木层对周边资源的利用。因此,建议当地在造林过程中以纯林为主,适当推广人工白桦林的栽植。
从不同灌木种群生态位重叠度的分配比例上看,其趋势表现为针阔叶混交林下的灌木种群生态位具有相当程度的重叠,对资源的共享趋势不太明显,显示现阶段该群落结构较不稳定,而纯林恰恰相反,表明人工林下灌木物种对提高单一群落结构的稳定性有一定的作用。
由生态位重叠度平均值和生态位宽度平均值研究结果可以看出,人工林下灌木物种对人工林单一群落结构的改善和物种多样性的提高有一定作用,这也是多物种并存的人工林配置模式筛选的原因和驱动机制之一。
一般而言,生态位宽度与生态位重叠度呈正相关关系,即生态位宽度较大的物种,其与其他种群间的生态位重叠度也较大,如白桦林群落中的高山柳与八宝卫矛、细叶绣线菊、金露梅的生态位重叠度均在0.800以上,其生态位宽度也均在0.960以上,这与前人的研究结果相似[18-19]。然而本研究结果表明,在云杉-白桦混交林和白桦人工林中,一些灌木物种生态位宽度较小,而生态位重叠度却较大,造成这种情况的原因可能与物种呈斑块性分布及环境资源的高度空间异质性有关,使适宜物种生活的环境呈斑块形式,由此导致物种呈少数不连续的斑块分布,造成物种常常较多地聚集在这些适宜斑块中,而在适宜斑块以外的空间分布较少,使物种在总体环境空间中生态位宽度较小,从而出现了生态位宽度较小而生态位重叠度较高的现象。
[参考文献]
[1] Grinell J.The niche relationship of theCaliforniathrasher[J].Auk,1917,21:364-382.
[2] 张继义,赵哈林,张铜会,等.科尔沁沙地植物群落恢复演替系列种群生态位动态特征 [J].生态学报,2003,23(12):2741-2746.
Zhang J Y,Zhao H L,Zhang T H,et al.Niche dynamics of main populations of plants communities in the restoring succession process in Horqin Sandy Land [J].Acta Ecologica Sinica,2003,23(12):2741-2746.(in Chinese)
[3] 缪 勇,邹运鼎,孙善教,等.棉蚜及其捕食性天敌时空生态位研究 [J].应用生态学报,2003,14(4):549-552.
Miao Y,Zou Y D,Sun S J,et al.Spatial and temporal niches ofAphisgossypiiand its predatory enemies [J].Chinese Journal of Applied Ecology,2003,14(4):549-552.(in Chinese)
[4] 刘玉清.生态位与商家经营定位 [J].商业研究,2003(6):11-13.
Liu Y Q.Ecological position and business positioning [J].Commercial Research,2003(6):11-13.(in Chinese)
[5] 韩 路,王海珍.生态位理论的发展及其在农业生产中的应用 [J].新疆环境保护,1999,21(4):10-15.
Han L,Wang H Z.Development and appliance in agriculture of niche theory [J].Environmental Protection of Xinjiang,1999,21(4):10-15.(in Chinese)
[6] 魏文超,何友均,邹大林,等.澜沧江上游森林珍稀草本植物生态位研究 [J].北京工业大学学报,2004,26(3):7-12.
Wei W C, He Y J, Zou D L,et al.Niche of rare herbage plants in the upstream of Lancang River [J].Journal of Beijing Forestry University,2004,26(3):7-12.(in Chinese)
[7] 方江平,卢 杰,罗 建,等.高寒植物长鞭红景天种群生态位研究 [J].安徽农业科学,2009,37(5):2027-2031.
Fang J P,Lu J,Luo J,et al.Study on the ecological niche of PsychrophyteRhodiolafasitiatapopulation [J].Journal of Anhui Agri Sci,2009,37(5):2027-2031.(in Chinese)
[8] 王伟伟,杨海龙,贺康宁,等.青海高寒区不同人工林配置下草本群落生态位研究 [J].水土保持研究,2012,19(3):156-165.
Wang W W,Yang H L,He K N,et al.The study herb community niche of different plantation configuration in alpine region of Qinghai Province [J].Research of Soil and Water Conservation,2012,19(3):156-165.(in Chinese)
[9] 王伟伟,杨海龙,贺康宁,等.祁连山青海云杉人工林草本层生态位及生态响应研究 [J].草地学报,2012,20(4):626-630.
Wang W W,Yang H L,He K N,et al.Niche and ecological response of herb layer in spruce plantation of Qilian Mountains [J].Acta Agrestia Sinica,2012,20(4):626-630.(in Chinese)
[10] Levens R. Evolution in changing environments:Some theoretical explorations [M].Princeton:Princeton University Press,1968.
[11] Pianka E R.The structure of lizard communities [J].Animal Revies of Ecology & Systematics,1973,4:53-74.
[12] 张 峰,上官铁梁.翅果油树群落优势种群生态位分析 [J].西北植物学报,2004,24(1):70-74.
Zhang F,Shangguan T L.Niche characteristics of dominant populations inElaeagnusmolliscommunities [J].Acta Botanica Boreal-Occidentalia Sinica,2004,24(1):70-74.(in Chinese)
[13] 胡正华,钱海源,于明坚.古田山国家级自然保护区甜槠林优势种群生态位 [J].生态学报,2009,29(7):3670-3677.
Hu Z H,Qian H Y,Yu M J.The niche of dominant species populations inCastanopsiseyreiforest in Gutian Mountain National Nature Reserve [J].Acta Ecologica Sinica,2009,29(7):3670-3677.(in Chinese)
[14] 王 刚.关于生态位定义的探讨及生态位重叠计测公式改进的研究 [J].生态学报, 1984,4(2):119-126.
Wang G.On the definition of niche and the improved formula for measuring niche overlap [J].Acta Ecologica Sinica,1984,4(2):119-126.(in Chinese)
[15] 李登武,张文辉,任争争.黄土沟壑区狼牙刺群落优势种群生态位研究 [J].应用生态学报,2005,16(12): 2231-2235.
Li D W,Zhang W H,Ren Z Z.Niche characteristics of dominant populations ofSophoradavidiicommunity in loess gully region [J].Chinese Journal of Applied Ecology,2005,16(12):2231-2235.(in Chinese)
[16] 李军玲,张金屯,郭逍宇.关帝山亚高山灌丛草甸群落优势种群的生态位研究 [J].西北植物学报,2003,23(12):2081-2088.
Li J L,Zhang J T,Guo X Y.Study on niche of dominant species of subalpine scrubland and meadow community in Guandi Mountains [J].Acta Botanica Boreali-Occidentalia Sinica,2003,23(12):2081-2088.(in Chinese)
[17] 马 晓,杨宇明,杜 凡,等.高黎贡山生物多样性研究:Ⅱ.印度木荷、硬斗石栎林主要树种生态位研究 [J].西南林学院学报,2007,27(1):15-19.
Ma X,Yang Y M,Du F,et al.Study on biodiversity of the Gao ligong Mountains:Ⅱ.Study on niche of the main tree species in Schima [J].Journal of Southwest Forestry College,2007,27(1):15-19.(in Chinese)
[18] 张忠华,梁士楚,胡 刚,等.桂林岩溶石山阴香群落主要种群生态位研究 [J].林业科学研究,2009,22(1):63-68.
Zhang Z H,Liang S C,Hu G,et al.Niche characteristics of dominant populations inCinnamomumburmanniicommunity on Karst hills of Guilin [J].Forest Research,2009,22(1):63-68.(in Chinese)
[19] 郑翠玲,曹子龙,赵廷宁,等.浑善达克沙地南缘农牧交错带弃耕地植被的演替规律 [J].中国水土保持科学,2005,3(1):72-76.
Zheng C L, Cao Z L, Zhao T N,et al.Succession disciplinarian of vegetation in abandoned land in agricultural cross bedding on south edge of Otindag sandy land [J].Science of Soil and Water Conservation,2005,3(1):72-76.(in Chinese)
