Purification, characterization and production optimization of a vibriocin produced by mangrove associated Vibrio parahaemolyticus
2014-03-23BaskarBalakrishnanJayappriyanKothilmozhianRanishreeSathishThadikamalaPrabakaranPanchatcharam
Baskar Balakrishnan, Jayappriyan Kothilmozhian Ranishree, Sathish Thadikamala, Prabakaran Panchatcharam
1Center for Bioenergy, Cooperative research, Lincoln University, Jefferson City, Missouri 65101, USA
2Marine Biotechnology, Andaman and Nicobar Centre for Ocean Science and Technology, National Institute of Ocean Technology, PortBlair, Andaman and Nicobar Islands 744103, India
3Department of Microbiology, PRIST University, Thanjavur, Tamil Nadu 614904, India
Purification, characterization and production optimization of a vibriocin produced by mangrove associated Vibrio parahaemolyticus
Baskar Balakrishnan1,2,3*, Jayappriyan Kothilmozhian Ranishree2, Sathish Thadikamala2, Prabakaran Panchatcharam3
1Center for Bioenergy, Cooperative research, Lincoln University, Jefferson City, Missouri 65101, USA
2Marine Biotechnology, Andaman and Nicobar Centre for Ocean Science and Technology, National Institute of Ocean Technology, PortBlair, Andaman and Nicobar Islands 744103, India
3Department of Microbiology, PRIST University, Thanjavur, Tamil Nadu 614904, India
PEER REVIEW
Peer reviewer
Dr. Nguyen Hoang Loc, Department of Biotechnology, Centre for Technology Incubation and Transfer, Hue University, Hue, Vietnam.
Tel: +84-54-6505051
Fax: +84-54-3830208
E-mail: nhloc@hueuni.edu.vn
Comments
This is a valuable research in which authors have improved the vibriocin production with the molecular weight of approximate 18 kDa from V. parahaemolyticus and demostrated its antibacterial activity.
Details on Page 260
Objective:To identify a potential bacterium which produces antimicrobial peptide (vibriocin), and its purification, characterization and production optimization. The bacteria subjected in the study were isolated from a highly competitive ecological niche of mangrove ecosystem.
Vibriocin, Vibrio parahaemolyticus, Vibrio harveyi, Mangrove rhizosphere, Antimicrobial peptide
1. Introduction
Antimicrobial peptides have been isolated from many living forms that include microbes, plants, invertebrates and vertebrates. Bacteria can produce different kinds of antimicrobial peptides, especially ribosomally synthesized antimicrobial peptides like bacteriocins, and non-ribosomally synthesized peptides like gramicidins, polymixins and bacitracins[1,2]. Bacteriocin produced by lactic acid bacteria is the extensively studied ones with an established classification system[3].
Bacteriocins produced by the genusVibrioare called vibriocins.They were reported from species such asVibrio cholerae(V. cholerae), Vibrio harveyi(V. harveyi), Vibrio mediterranei(V. mediterranei), Vibrio vulnificus(V. vulnificus) and someVibriosp., possess good antibacterial activity against their own closely relatedVibriomembers such asV. cholerae,V. harveyi,V. mediterranei,Vibrio parahaemolyticus(V. parahaemolyticus) ,V. vulnificusandVibrio fischeri[3-9]. Though there are reports available on the bacteriocins produced by the genusVibrio, such reports are only at latent stage of basic research. Being capable of producing bacteriocins againsttheir own closely related species and having diverse ecological adaptations, the genusVibriocould be an interesting candidate to search for bacteriocins, for the effective disease management in mariculture industry.
V. parahaemolyticusis a widely reported bacterium (both pathogenic and non-pathogenic) associated with the flora and fauna of the marine environments[10]. It is found even in rhizosphere of the mangrove plants as a heterotrophic diazotroph[11]. This organism is a major cause of acute gastroenteritis in humans through the contaminated sea foods, particularly in the areas of the world where sea food consumption is high[12]. Still the organism is widely studied for its harmfulness, and there are not too many reports available on its beneficial aspects.
Mangrove has a unique microbial diversity[13]. It is a highly challenging environment as it is nutritionally rich and dynamic due to its frequent change of physical and chemical conditions. Adaptable studies of these microbial communities would be helpful in understanding the nature of the habitat[14]. Only few works have been carried out on the rhizosphere microbiota of the mangrove plants[15]. Hence, the investigation of this microbiota will lead to the identification of novel, biotechnologically potent microorganisms having unambiguous variety of applications. Pichavaram mangrove is one of the important mangrove forests in India withAvicennia marina(A. marina) as the dominant plant flora[16].
In the present study, an effort has been made to purify, characterize and production optimization of vibriocin produced by the bacterium isolated from the rhizosphere ofA. marinapresent in the Pichavaram mangrove ecosystem. This compound showed good antibacterial activity against the mariculture pathogenic members ofVibrio.
2. Materials and methods
2.1. Selection of potential isolate
A total of 66 isolates used in this study were isolated from the rhizosphere soil of Pichavaram (latitude 11°23’-11°30’N and longitude 79°45’-79°50’E) mangrove plantAvicennia marinaand all the isolates were characterized using phenotype and 16S rRNA gene sequence analysis. All the isolates were cultivated in Zobell’s Marine broth for 24 h at 37 °C in an orbital shaker at 150 r/min. Culture broths were centrifuged to get cell free supernatant (CFS), which were sterilized through 0.22 µm filter paper (Millipore, Bedford, MA, USA) and subjected to agar well diffusion assay to check the cultures for the production of antimicrobial metabolites. Eighteen hour cultures of pathogenic type strains obtained from Microbial Culture Collection Centre (MTCC), India and environmental isolates (Table 1) were diluted with pre-sterilized normal saline and the turbidity of the culture was adjusted to 0.5 McFarland turbidity. Lawns of cultures were prepared by sterilized cotton swabs over the surface of the Brain Heart Infusion (BHI) agar. Wells were made in the inoculated plates using sterile metal borer. A total of 20 µL CFS were added in the wells, and the plates were incubated at 37 °C for 24 h. Then, the zone of inhibition was observed. The vibriocin titre and activity unit (AU) was determined with indicator type strain [V. harveyi(MTCC 3438)] in each experiment using the method described previously by Motta and Brandelli[17].
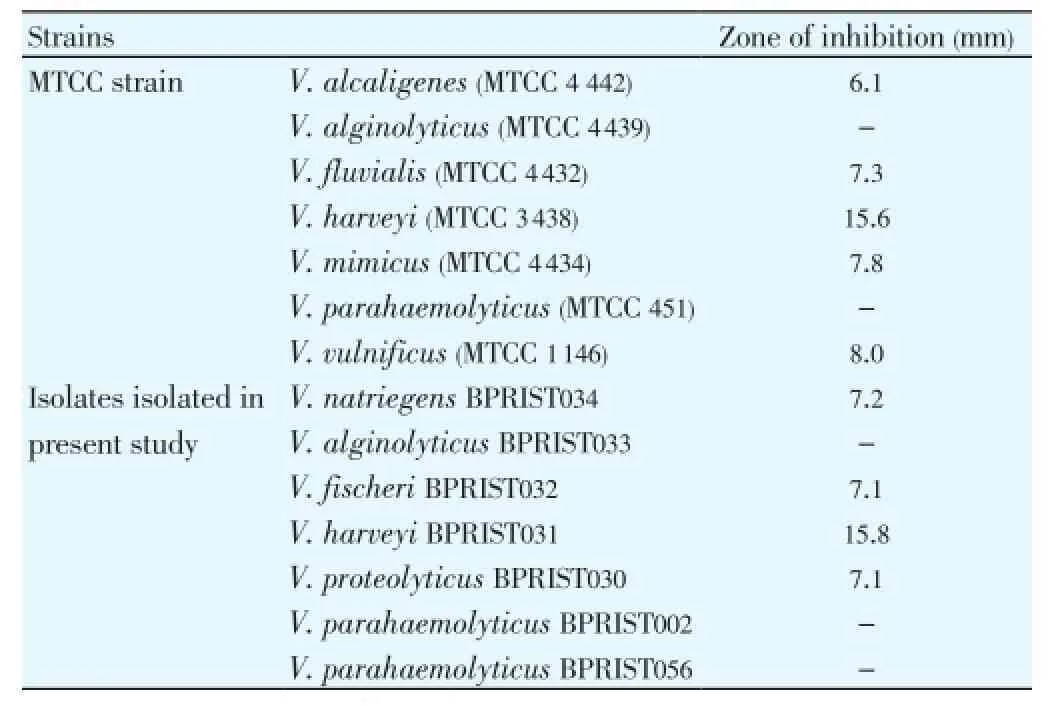
Table 1 Antibacterial activity of the culture free supernatant against type strains and environmental isolates of the genus Vibrio.
2.2. Characterization of potential isolate
Phenotypic characterizations were performed as reported by Alsina and Blanch[18] and Farmer and Janda[19]. Further, phenotypic characters of the isolate BPRIST035 were compared with the type strain ofV. parahaemolyticus(MTCC 451). For 16S rRNA gene sequencing and phylogenetic analysis, the genomic DNA of the potential isolate was extracted using the method described by Chen and Kuo[20]. PCR amplification and sequencing were performed using universal primers pA (5’-AGA
GTT TGA TCC TGG CTC AG-3’) and pH (5’-AAG GAG GTG ATC CAG CCG CA-3’). The partial 16S rRNA gene sequence of the isolate was compared with 16S rRNA gene sequences available with the BLASTN search of the NCBI, GenBank database (http://www. ncbi.nlm.nih.gov). Phylogenetic dendrogram was constructed by the neighbour-joining method using MEGA 4.1 (Molecular Evolutionary Genetic Analysis). The partial 16S rRNA gene sequence of the isolate BPRIST035 was submitted in GenBank data base and obtained the accession number (JF431424).
2.3. Purification of the vibriocin
The chosen potential strain was subjected to mass cultivation in Zobell’s Marine broth for 24 h at 37 °C in an orbital shaker at 150 r/min. The supernatant was separated by centrifugation and sterilized through 0.22 µm filter paper. The culture filtrate was precipitated with ammonium sulphate at 60% (w/v) saturation. The precipitate was recovered by centrifugation at 10 000gfor 30 min at 4 °C and dissolved in phosphate buffer saline (PBS) and then extracted thrice with 1-butanol and evaporated with reduced pressure[21]. The resultant was suspended in 10 mL of 10 mmol/L phosphate buffer (pH 7.5) and designated as crude vibriocin.
The crude vibriocin was subjected to gel permeation chromatography using Sephadex G-100 column. Before elution, the column was pre-equilibrated with 20 mmol/L sodium phosphate pH 7.5, and then the sample was eluted with PBS and fractionated by 2 mL. Eluted fractions were tested forthe antibacterial activity against the indicator strain. The active fractions were pooled and subjected to ion-exchange chromatography using diethyl-aminoethanol cellulose anion exchange column, pre-equilibrated with 20 mmol/L sodium phosphate pH 7.5, then the sample was eluted with the gradient of 0 to 1.5 mol/L NaCl prepared in PBS and fractionated by 6 mL. Each fraction was subjected to antibacterial activity against the indicator strain. The fraction showing highest antibacterial activity was dialysed against double distilled water, subsequently freeze-dried and used for further analysis. Final purification of the sample was achieved with HPLC (Shimadzu, Japan) using a C18 column, UV detector absorbance at 280 nm and eluted with acetonitrile and PBS mixture (1:10), collected fractions tested for antibacterial activity against indicator strain by using above mentioned method. At each purification step, CFS, precipitation, extraction, gel filtration and ion exchange chromatography, protein concentration was evaluated (A280 nm).
2.4. Molecular weight determination of the vibriocin
To determine the molecular weight of purified vibriocin, sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDSPAGE), (14% Polyacrylamide gel) was performed as described previously[22]. Medium range molecular weight markers (Sigma, USA) were used. Purity of the vibriocin was compared with crude and partially purified vibriocin. Band appeared on the gel was separated and washed with sterile double distilled water in order to remove the SDS, then placed in sterile Petri plate and flooded the BHI agar with the indicator strainV. harveyiover the gel, then the plate was incubated for 24 h at 37 °C and the zone of inhibition was observed. Remaining gel portion was subjected to Coomassie brilliant blue staining to visualise the markers and the target protein, the observed results were used for molecular weight identification of the vibriocin.
2.5. Effect of physiochemical factors on the vibriocin activity
Vibriocin was treated with various enzymes (proteinase, pepsin, trypsin, pronase E, lysosyme, lipase and α-amylase) as described by Prasadet al[7]. To analyze thermal stability, Purified vibriocin was treated to 50, 75, 100 and 125 °C for 15 min respectively. Sterile Petri plates containing 5 mL aliquots of vibriocin were exposed to UV irradiation for 15 to 90 min by placing at 30 cm from the UV bulb[23]. The activity of vibriocin at pH range (2-12) was tested at ambient temperature. The detergents, Tween 80 and Triton-×-100 were treated by preparing the final concentration at 2% (v/v) and for SDS at 2% (w/v), for metalloprotease inhibitor EDTA, 1% (w/v) was prepared with vibriocin. All these preparations were incubated for 15 min at ambient temperature. The effect of solvents (acetic acid, acetone, benzene, chloroform, dichloromethane, dichloromethane, dmso, ethanol, ethyl acetate, hexane, isopropanol and methanol) were tested by equal proportion of vibriocin and the solvent was mixed well and incubated for 1 h at ambient temperature. After the physiochemical treatments, the treated vibriocin was subjected to antibacterial activity assay against indicator strain.
2.6. Optimization of the vibriocin production
In order to improve vibriocin production by potential bacterium, seven critical parameters,viz., NaCl concentration, temperature, agitation speed, pH, inoculum concentration, carbon and nitrogen source were optimized. A 32 runs orthogonal array (L32-OA) design was used to investigate the effect of selected parameters on vibriocin production. In Taguchi’s method, quality is measured by the deviation of a characteristic from its target value and a loss function [L(y)]. The loss function was calculated by the Eq(1).
Eq (1): L(y)=k×(y-m)2
Where, k=The proportionality constant, m=Target value and y =Experimental value obtained for each trial.
In the present study, the goal is to achieve the higher vibriocin production. Such case “bigger-is-better” was chosen. In case of the “bigger-is-better” quality characteristics, the loss function can be written as
Eq (2): L(y)=k×(1/y2)
In such case the expected loss function can be represented by
Eq (3): E[(Ly)]=k×E (1/y2)
Where E(1/y2) can be estimated from a sample of n as
To explore the obtained results, analysis of variance (ANOVA) test was performed. Further conformational experiments were performed in order to validate the optimum conditions. Qulitek-4®software, Nutek Inc. was used for designing and analysis of the experiments.
3. Results
3.1. Antibacterial activity
Among the 66 isolates screened, the isolate PBRIST035 showed good antibacterial activity against variousVibriotype strains, as well as environmental isolates exceptV. parahaemolyticusandV. alginolyticus. The maximum inhibitory activity was observed againstV. harveyiand minimum inhibitory activity against type strain ofV. alcaligenes(MTCC 4442). Table 1 shows the antibacterial activity of isolate PBRIST035 against the variousVibriotype and wild strains.
3.2. Characterization of potential isolate
16S rRNA gene sequence and phylogenetic analysis of the potential isolate showed 99.6% similarity withV. parahaemolyticusATCC 17802T and 99.5% similarity withV. alginolyticusATCC 17749T and the cluster formed by PRIST035 was evidenced by high bootstrap values (Figure 1). However, based on the results of phenotypic characterization the potential organism was identified asV. parahaemolyticusand the comparison study with the type strain,V. parahaemolyticus(MTCC 451) also confirmed the same (data not given), sinceV. parahaemolyticuscannot ferment sucrose and has a negative Voges Proskauer reaction, this was distinguished fromV. alginolyticus[19,24]. As like otherVibriomembers, it can withstand a wide range of pH and NaCl concentration. Tolerance up to 12 % NaCl concentration had confirmed its halotolerance.
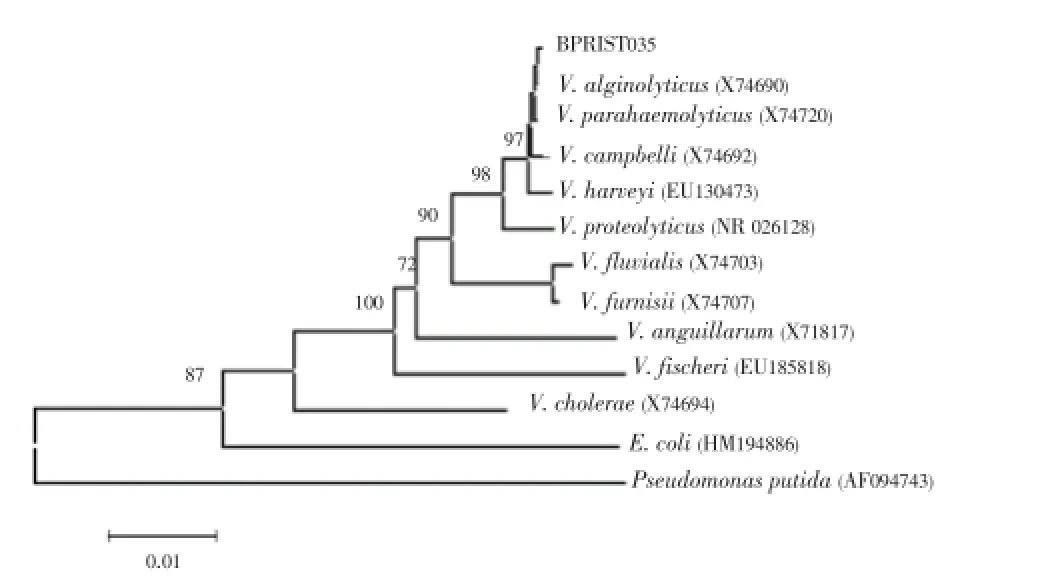
Figure 1. Neighbour-joining phylogenetic tree is based on 16S rRNA gene sequence of the potential isolate and their closest phylogenetic neighbours. Bootstrap values are indicated at nodes. Scale bar represents observed number of changes per nucleotide position.
3.3. Purification and molecular weight identification
Purification steps and recovery values of the peptide are given in Table 2. Where the total activity represents the activity given by the purified product and the specific activity represents activity with the amount of protein observed in each purification step. Extraction with 1-butanol showed the huge increase in the purification with 120.5 fold of increment. Gel permeation chromatography also showed 173.8 fold increases in the purification from the CFS. From the ion-exchange chromatography, the target protein was eluted at 1.1% of mol/L NaCl concentration and the final purification fold increased up to 260.4. The partially purified vibriocin showed 74.66% of purity, and pure vibriocin was eluted at the retention time of 2.92 min (Figure 2) in HPLC. The purified vibriocin showed 18 kDa of molecular weight in the SDS-PAGE (Figure 3).

Table 2 Purification and activity of the vibriocin.
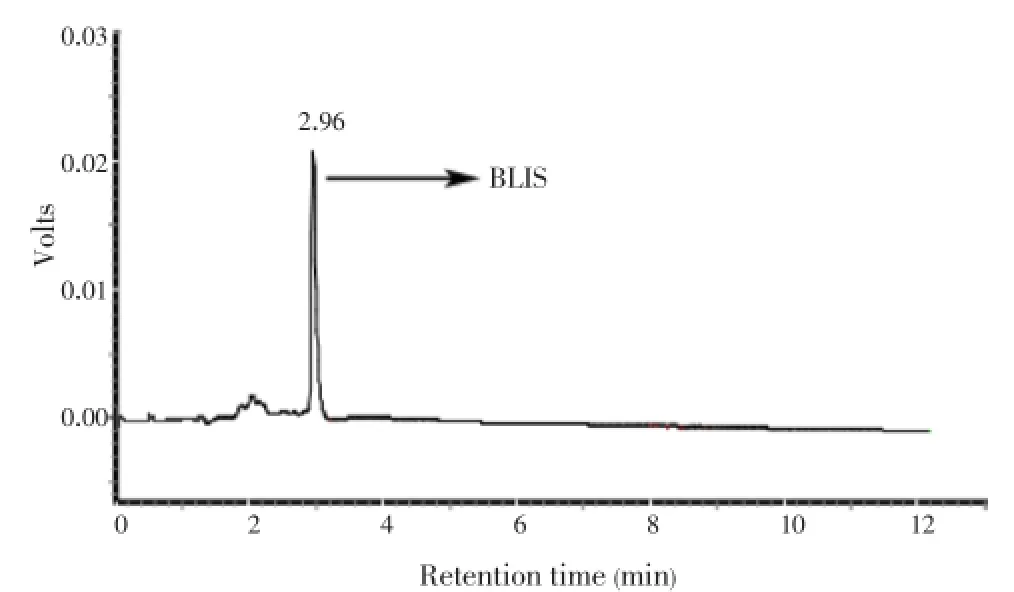
Figure 2. HPLC chromatogram showing a major peak corresponding to vibriocin.

Figure 3. SDS-PAGE of the BLIS.A: Molecular mass markers; B: Crude vibriocin; C: Partially purified vibriocin; D: Purified vibriocin; E: Vibriocin activity on the overlaid gel with a lawn of V. harveyi cells.
3.4. Effect of physiochemical parameters on the vibriocin activity
The vibriocin produced byV. parahaemolyticusPBRIST035 was stable in non proteolytic enzyme and unstable with proteolytic enzymes used. The vibriocin was stable in all the pH tested
(pH 2-12) and resists the heat up to 75 °C for 15 min and resist the UV radiation up to 75 min exposure. Presence of residual activity were observed in detergents and polar solvents were used. The effect of physiochemical parameters on the vibriocin is summarised in Table 3.

Table 3 Effect of physiochemical parameters on the vibriocin activity.
3.5. Production optimization
According to the experimental plan (L-32) the experiments were conducted, and the data revealed significant variation in the vibriocin production (Table 4). Minimum and maximum vibriocin production values were observed to be 490 and 2 351 U/mL under the selected fermentation conditions. The average effect of the factors at the assigned levels on the vibriocin production byV. parahaemolyticusPBRIST035 is shown in Figure 4. At the individual level stage, the highest effect on vibriocin production was noticed with NaCl concentration at 4th level (1 605.00) followed by inoculum concentration at 2ndlevel (1 603.25), pH at 2nd level (1 472.13), agitation speed at 1st level(1 469.38), peptone at 3rd level (1 453.38) temperature at 3rd level (1 443.25) and glucose at 1st level(1 400.75). The selected variables preferences have followed the order:
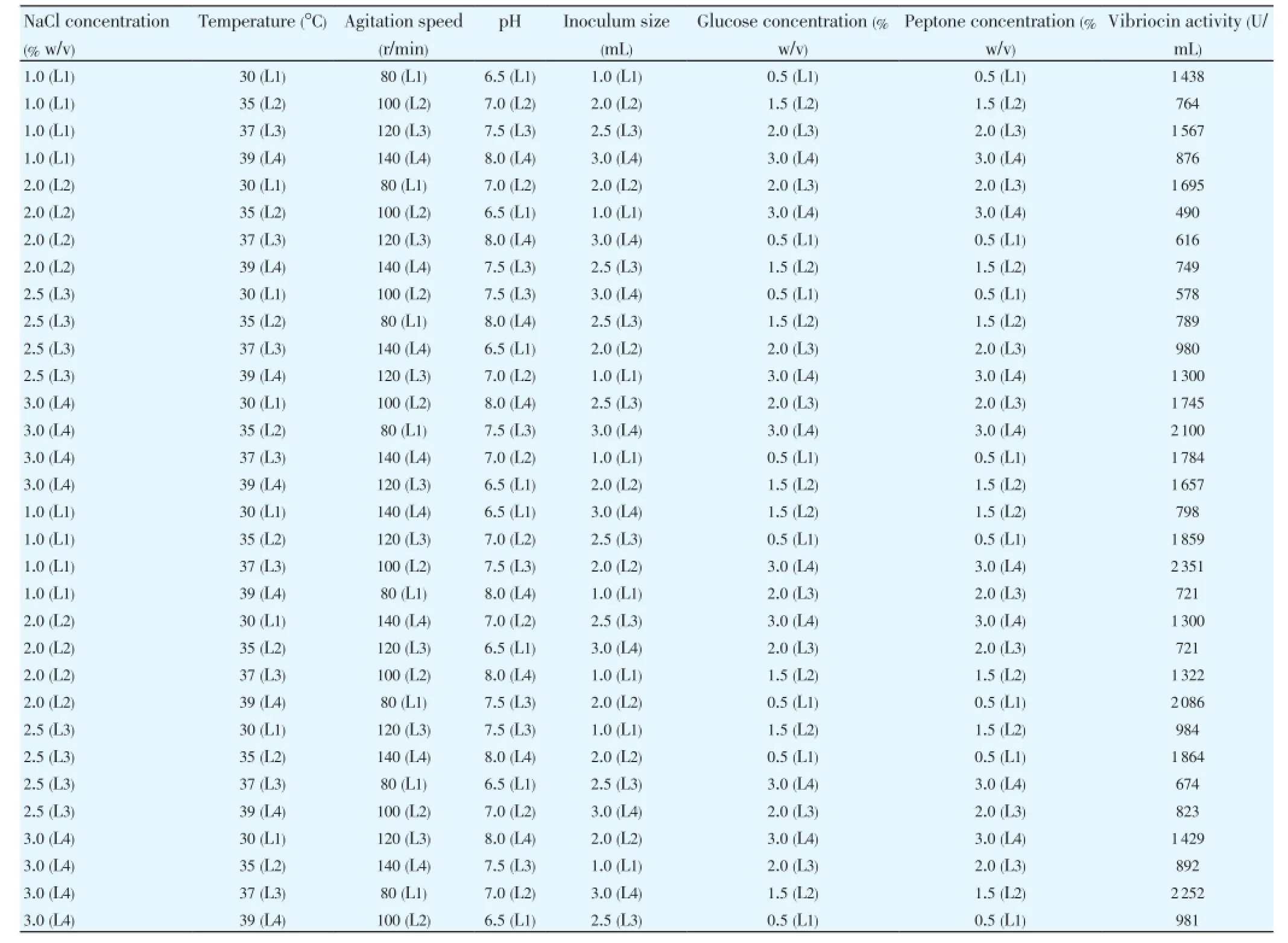
Table 4 L-32 orthogonal design along with the obtained vibriocin activity produced by V. parahaemolyticus BPRIST035.

Table 5 Interaction influence, ANOVA and optimized conditions for selected factors for vibriocin production by V. parahaemolyticus BPRIST035.
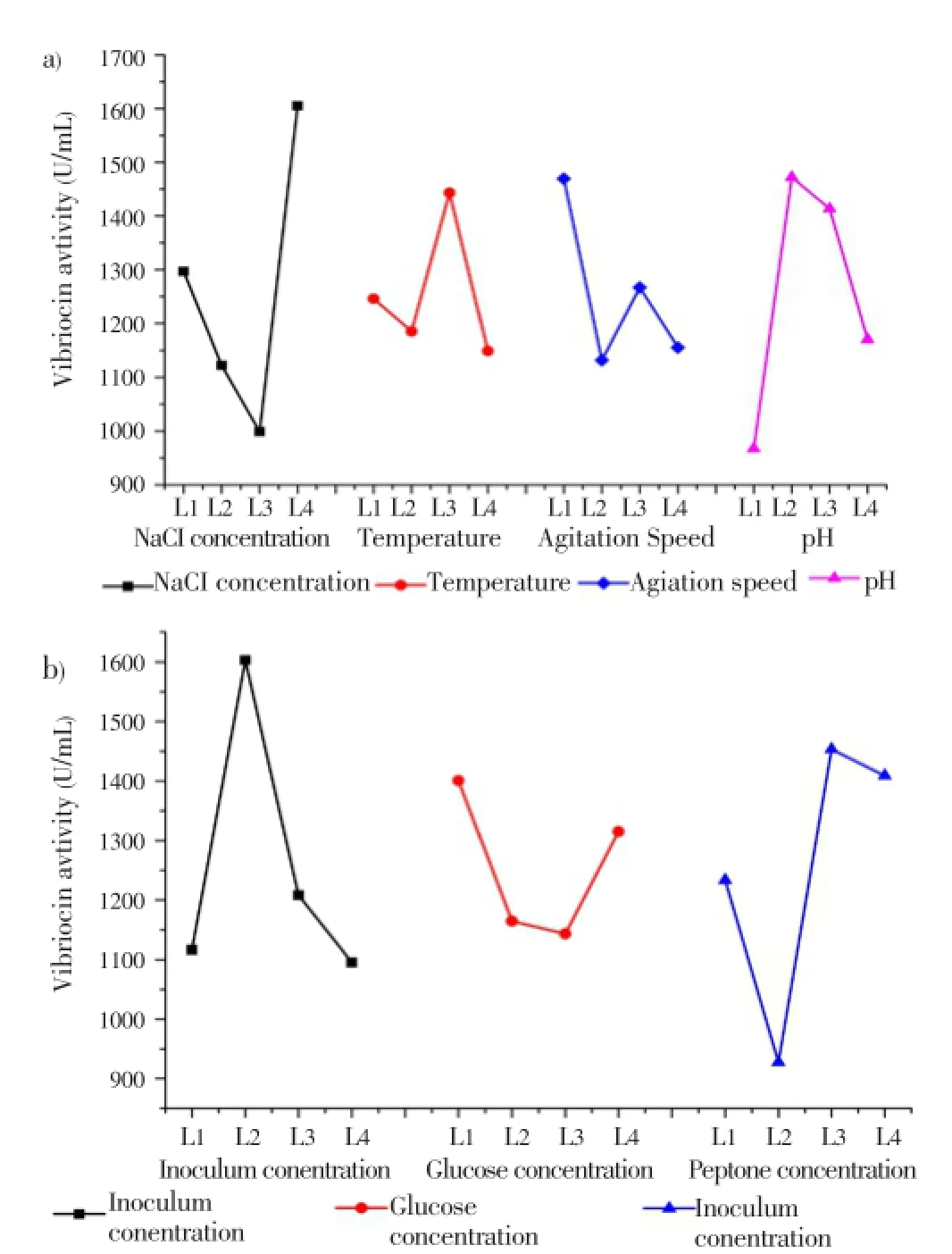
Figure 4. Main effects of the selected parameters at different levels on vibriocin production by V. parahaemolyticus BPRIST035.a: NaCl, temperature, agitation speed and pH. b: Inoculum, glucose and peptone.
NaCl>Inoculum>pH>Agitation>Peptone>Temperature>Glucose
The levels of factors to produce the best results showed ANOVA with the percentage of contribution of each factor with interactions shown in Table 5. It was observed that incubation temperature has the major contribution (23.64%) on overall fermentation process for effective vibriocin production. After temperature, peptone concentration (19.477%) and inoculum concentration has the highest effect followed by agitation speed and NaCl concentration on vibriocin production byV. parahaemolyticusPBRIST035. Glucose concentration and pH has the least significance (5.199 & 5.899%) on antibiotic production. The observed very low error contribution (0.004%) indicates the accuracy of the experimental data.
The Taguchi DOE design, used in the present study for the maximizing the production of vibriocin fromV. parahaemolyticusPBRIST035, revealed the effect of interaction of varying factors. Higher levels of vibriocin production were achieved with the software generated culture conditions: 3% NaCl, at the temperature of 37 °C, at the agitation speed of 80 r/min, pH 7, 2% inoculum concentration, 0.5% glucose concentration and 2% peptone concentration. The expected vibriocin yield at these optimum conditions is 2 912 U/mL with a total contribution from all the factors being 1 656 U/mL with grand average performance of 1 255 U/mL. Performing the validation experiments at the predicted conditions 2 187 U/mL activity was achieved. The obtained antibacterial activity is nearer to the predicted value. The validation experimental data at predicted optimum conditions revealed that 466 U/mL activity was improved.
The expected improvement on overall vibriocin production by this bacterial strain under submerged fermentation could be viewed from Figure 5. This figure explains the variation reduction achieved with optimized conditions. The improved S/N ratio at optimum condition depicts the reduction in the standard deviation before optimization.
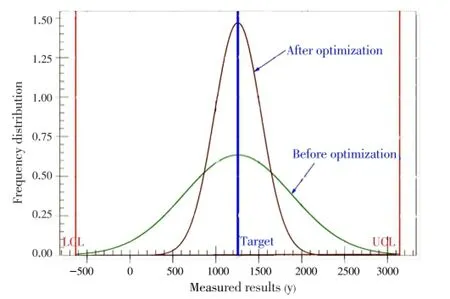
Figure 5. Variation reduction plot on vibriocin production by V. parahaemolyticus BPRIST035.
4. Discussion
This study revealed the identification of novel vibriocin producingV. parahaemolyticusBPRIST035, which is highly active againstV. harveyi, a major mariculture pathogen, and the vibriocin was purified, characterized and increased production through optimizing the growth factors. ThoughV. parahaemolyticusis reported to be a diazotrophic rhizobacteria of mangrove plant[11], the ability of producing vibriocin in the competitive ecological niche is the first report ever made. CFS of the potential isolate did not exhibit intra species activity, an attribute unique to this strain. Prasadet al.[7] reported that the bacteriocin produced byV. harveyiVIB 571 shown activity against type strains, as well as environmental isolates ofV. harveyi. Carraturoet al.[4] had reported that bacteriocin produced byV. mediterranei1 showed good activity against environmental isolates of the same species. Same kind of results was obtained by Shehane and Sizemore[8] forV. vulnificusIW1, BC1 and BC2. Furthermore, the CFS did not exhibit any activity againstV. alginolyticus, which may be due to its taxonomically close relatedness withV. parahaemolyticusspecies. Moreover,V. alginolyticuswas previously known as biotype 2 ofV. parahaemolyticus[24]. Indeed, the 16S rRNA gene sequence analysis of the potential strain also revealed the same. CFS showed excellent activity againstV. harveyiMTCC 3438, which was twofold higher than activity observed with other type strains used. Same kind of activity was observed in the isolates ofVibriofrom the same ecological niche. With these, it is clearly evident that the vibriocin produced by theV. parahaemolyticusBPRIST035 has good specificity against theV. harveyi. Hence, the strain,V. harveyi(MTCC 3438) was used as an indicator strain throughout the study. This feature contributes significant scope for the use of BLIS for the disease management against pathogenicV. harveyiin mariculture industries.
Vibriocin was released in the culture media in a relatively low concentration. In order to maintain the conditions of isolation procedure and strain’s halotolerant nature, Zobell marine broth was used as the production medium. Though the quantity of vibriocin produced is low, it is good enough for the preliminary characterization. Accordingly, the vibriocin was concentrated by ammonium sulphate precipitation. Extractions with butanol showed a huge increase in the purification which is comparable to the Bizaniet al.[21] results on the purification of cerein 8A and the same type of results were observed for gel permeation chromatography and ion-exchange chromatography. The purified vibriocin showed unique molecular mass (18 kDa) compared to the notable bacteriocins reported from the members ofVibrio, like 63-65 kDa Bacteriocin Like inhibitory substance (BLIS) produced byV. mediterranei[4], 24 kDa harveyicin produced byV. harveyi[5,6], 32 kDa BLIS fromV. harveyi[7] and 1.35 kDa, 7.5 kDa and 9 kDa multiple peptides produced byV. vulnificus[8] and also well classified bacteriocins reported in other organisms till date[25].
As like other widely reported bacteriocin, the vibriocin produced byV. parahaemolyticusPBRIST035 was sensitive to all proteolytic enzymes used, but on the converse, it is resistant to other used enzymes, which indicated that the vibriocin is only proteinaceous in nature and not having any carbohydrate or lipid moiety in its structure. This is in contrast to the BLIS produced byV. harveyi, which exhibits lipid moiety in its structure[7] and BLIS was produced byV. vulnificusIW1 and BC1 which has carbohydrate moiety in it structure[8]. In the same way, BLIS produced byV. mediterranei1 is resistant to trypsin[4]. Stability of the vibriocin in pH, heat and UV radiation, are useful for the application of the vibriocin as medicated feeds in mariculture industries. Because generally these characters will be considered for the storage and application of a feed, this is one of the easiest and economically important ways of drug delivery in the aquaculture[26]. The ability of the vibriocin existence in the marine medium and not losing its activity indicates that it can survives very well in the marine water conditions thus the application of the compound in mariculture industries holds lot of scopes. As like other bacteriocins, it is resistant to detergents like Tween 80, Triton ×-100 and SDS as also to EDTA. Vibriocin resistance to the SDS once again confirmed its mono peptide nature. In the same way, it is resistant to all polar solvents and sensitive to all non polar solvents used. On the whole, the vibriocin produced byV. parahaemolyticusis showed uniqueness and compatibility with other reported bacteriocins or BLIS in its character as well as in the structure[25].
Production of vibriocin byV. parahaemolyticusmight be the sole defensive mechanism to survive in the particular ecological niche, when comparing with otherVibriomembers isolated from the same niche, especially withV. harveyi, because population wiseV. harveyiwas the dominant member of the genus in the ecological niche by showing highest bacterial count.V. parahaemolyticusBPRIST035 did not possess any other rhizobacteria characters like N2 fixation, phosphate solubilization and/ or IAA production to survive in the rhizosphere. Moreover, it is the only strain showed the production of vibriocin among the three strains ofV. parahaemolyticus(BPRIST002, BPRIST034 and BPRIST056) isolated from different sites of the mangrove forest (results not given).
Characterization and optimization of each factor which influence the growth and yield of desired product are essential requirements before the selected strain is used for further investigation. In preliminary studies (data not given), it revealed that the secretion of vibriocin depends on the physiological and nutritional parameters as well as biochemical nature of the bacterium. Hence, further investigations were made to understand various requirements for vibriocin production byV. parahaemolyticusBPRIST035. A Taguchi experimental design was employed to investigate the interaction between the selected parameters and optimize the conditions for enhanced antibiotic production. The observed variation in vibriocin yield at various experiment certainly indicates the imperative role of optimization of all fermentation related factors in achieving the best possible yields.
It was observed that increased NaCl concentration played a vital role on vibriocin production. It is obvious that it could improve the bacterial growth as the bacteriumfrom marine origin and slightly halophilic in nature. There are several reports that showed increased production of bacteriocin produced by members ofLactobacillusby increased NaCl concentration[27,28]. It can also be discussed as the condition may mimic the environmental condition where the bacterium tend to produce vibriocin as a defensive product and the adopted expression mechanism of the bacterium to the particular salt concentration.
Next to NaCl inoculum size played an important role in the increased production of vibriocin. High inoculums will attain the late log phase quickly with increased biomass at this phase and may lead higher production of vibriocin. In the present study, it was observed that the vibriocin activity was first detected in late log phase and it confirms that the vibriocin is a secondary metabolite. Other parameters were not showed much effect in the increased vibriocin production.
In Taguchi method, the interaction between the two factors could be representing as interaction severity index (SI) which indicates the percent interaction between two factors. From Table 5, it was observed that interaction between NaCl concentration and agitation speed is the highest (48.5%) when compared with other factor interactions. Least SI was observed with the interaction between the inoculum concentration and agitation speed (5.29%). At the individual level, inoculum concentration has shown higher effect and the agitation speed showed moderate effect on vibriocin production whereas interaction of these two has least SI.
Though temperature and glucose have the least effect on the vibriocin production at individual levels their interaction has second highest SI (44.42%), which indicates that the production of antibiotic is depending on the individual as well as interaction effect of selected parameters. On the whole, using Taguchi experimentation enhanced the vibriocin yield from 2 351 to 2 187 U/mL (~20 % improvement) with the modified culture conditions.
To conclude, less effectiveness of chemically synthesized drugs in marine water leads to uncontrolled use in mariculture, and results in serious toxicological consequences on the living organisms[26]. The vibriocin obtained from the present study provided ample scope for the successful disease management of mariculture industry devastated byV. harveyi. In fact, this may be the first report on the production of vibriocin with novel characters by a halotolerantV. parahaemolyticus, which has substantial activity against both type strain and environmental isolate ofV. harveyi. Extracellular production of vibriocin in the marine medium makes it easy to purify and scale up at industrial level. The obtained antibiotic has resistance to a wide range of pH, temperature, UV radiation and chemicals which designates it is best suitable for mariculture industry. This study also signifies the orthogonal method of optimization to improve the production of antibiotics with a minimum number of experiments.
Conflict of interest statement
We declare that we have no conflict of interest.
Acknowledgements
The authors are grateful to Dr. R. Kirubagaran and Dr. N.V. Vinithkumar, Marine Biotechnology, National Institute of Ocean Technology, India for sparing laboratory facilities from the Project Drug from Sea, funded by Ministry of Earth Sciences, Govt. of India (Grant # 34030020005).
Comments
Background
Vibriocins are a group of bacteriocins produced by Gram-negative bacteria in the genusVibrio. Like other bacteriocins, vibriocins are protein toxins. They can kill bacteria beyond the genusVibrio, including other proteobacteria. Mariculture industry is drastically affected by the diseases caused by theVibrios which usually result in economic losses. The present study identified the potential candidateV. parahaemolyticusisolated from a highly competitive ecological niche of mangrove ecosystem, which produces vibriocin active against pathogenicVibrios.
Research frontiers
The present study performed a purification, characterization and production optimization of vibriocin produced byV. parahaemolyticusisolated from the rhizosphere ofAvicennia marinapresent in the Pichavaram mangrove ecosystem. The vibriocin showed good antibacterial activity against the mariculture pathogenic members ofVibrio.
Related reports
There are many reports on bacteriocins production by theVibrios such asV. cholerae, V. harveyi, V. mediterranei, V. vulnificusand someVibriosp. The bacteriocins exhibited good antibacterial activity against their own closely relatedVibriomembers. However, according to our knowledge no work has been carried out on bacteriocin production fromV. parahaemolyticus.
Innovations and breakthroughs
The authors used statistical tools and culture parameters to optimize the vibriocin production. An overallapproximately 20% of antibiotic production was improved, it was noticed that sodium chloride and agitation speed played a vital role in secretion of vibriocin.
Applications
The vibriocin produced fromV. parahaemolyticusmay be an effective alternative for chemically synthesized drugs.
Peer review
This is a valuable research in which authors have improved the vibriocin production with the molecular weight of approximate 18 kDa fromV. parahaemolyticusand demostrated its antibacterial activity.
[1] Hancock RE, Chapple DS. Peptide antibiotics. Antimicrob Agents Chemother 1999; 43(6): 1317-1323.
[2] Joerger RD. Alternatives to antibiotics: bacteriocins, antimicrobial peptides and bacteriophages. Poult Sci 2003; 82(4): 640-647.
[3] Klaenhammer TR. Genetics of bacteriocin produced by lactic acid bacteria. FEMS Microbiol Rev 1993; 12: 39-85.
[4] Carraturo A, Rajeta K, Ottaviani D, Russo GL. Inhibition of Vibrio parahaemolyticus by a bacteriocin-like inhibitory substances (BLIS) produced by Vibrio mediterranei 1. J Appl Microbiol 2006; 101: 234-241.
[5] Hoyt PR, Sizemore RK. Competitive dominance by a bacteriocin producing Vibrio harveyi strain. Appl Environ Microbiol 1982; 44(3): 653-658.
[6] McCall JO, Sizemore RK. Description of a bacteriocinogenic plasmid in Beneckea harveyi. Appl Environ Microbiol 1979; 38(5): 974-979.
[7] Prasad S, Morris PC, Hansen R, Meaden PG, Austin B. A novel bacteriocin-like substance (BLIS) from a pathogenic strain of Vibrio harveyi. Microbiol 2005; 151: 3051-3058.
[8] Shehane SD, Sizemore RK. Isolation and preliminary characterization of bacteriocins produced by Vibrio vulnificus. J Appl Micrbiol 2002; 92(2): 322-328.
[9] Sugita H, Matsuo N, Hirose Y, Iwato M, Deguchi Y. Vibrio sp. strain NM 10, isolated from the intestine of a Japanese coastal fish, has an inhibitory effect against Pasteurella piscicida. Appl Environ Microbiol 1997; 63(12): 4986-4989.
[10] Kaneko T, Cowell RR. Ecology of Vibrio parahaemolyticus in Chesapeake Bay. J Bacteriol 1973; 113(1): 24-32.
[11] Criminger JD, Hazen TH, Sobecky PA, Lovell CR. Nitrogen fixation by Vibrio parahaemolyticus and its implications for a new ecological niche. Appl Environ Microbiol 2007; 73(18): 5959-5961.
[12] Daniels NA, MacKinnon L, Bishop R, Altekruse S, Ray B, Hammond RM, et al. Vibrio parahaemolyticus infections in the United States, 1973-1998. J Infect Dis 2000; 181(5): 1661-1666.
[13] Smith TJ, Boto KG, Frusher SD, Giddins RL. Keystone species and mangrove forest dynamics: the influence of burrowing by crabs on soil nutrient status and forest productivity. Estuar Coast Shelf Sci 1991; 33: 419-432.
[14] Semenov AM, van Bruggen AHC, Zelenev VV. Moving waves of bacterial population and total organic carbon along roots of wheat. Microbial Ecol 1999; 37(2): 116-128.
[15] Kathiresan K, Selvam MM. Evolution of beneficial bacteria from mangrove soil. Bot Mar 2006; 49: 86-88.
[16] Kathiresan K. A Review of studies on Pichavaram mangrove, Southern India. Hydrobiologia 2000; 430: 185-205.
[17] Motta AS, Brandelli A. Characterization of an antimicrobial peptide produced by Brevibacterium linens. J Appl Microbiol 2002; 92(1): 63-70.
[18] Alsina M, Blanch AR. Improvement and update of a set of keys for biochemical identification of Vibrio species. J Appl Bacteriol 1994; 77(6): 719-721.
[19] Farmer JJ, Janda JM, Family I. Vibrionaceae. In: Brenner DJ, Krieg NR, Staley JT, Garrity GM, editors. Bergey’s manual of systematic bacteriology. 3rd ed, vol 2. New York: Springer; 2005, p. 491-555.
[20] Chen WP, Kuo TT. A simple and rapid method for the preparation of gram negative bacterial genomic DNA. Nucleic Acids Res 1993; 21(9): 2260.
[21] Bizani D, Dominguez AP, Brandelli A. Purification and partial chemical characterization of the antimicrobial peptide cerein 8A. Lett Appl Microbiol 2005; 41(3): 269-273.
[22] Hames BD. An introduction to polyacrylamide gel electrophoresis. In: Hames BD, Rickwood D, editors. Gel electrophoresis of proteins. UK: IRL Press; 1981, p. 1-92.
[23] Ogunbanwo ST, Sanni AI, Onilude AA. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr J Biotechnol 2003; 2(8): 219-227.
[24] Pien F, Lee K, Higa H. Vibrio alginolyticus infections in Hawaii. J Clin Microbiol 1977; 5(6): 670-672.
[25] Desriac F, Defer D, Bourgougnon N, Brillet B, Chevalier PL, Fleury Y. Bacteriocin as weapons in the marine animalassociated bacteria warfare: inventory and potential applications as an aquaculture probiotic. Mar Drugs 2010; 8(4): 1153-1177.
[26] Rodgers CJ, Furones MD. Antimicrobial agents in aquaculture: Practice, needs and issues. In: Rodgers C, Basurco B, editors. The use of veterinary drugs and vaccines in Mediterranean aquaculture. Series A: Mediterranean Seminars, No. 86. Spain: CIHEAM; 2009, p. 41-59.
[27] Verluyten J, Messens W, De Vuyst L. Sodium chloride reduces production of curvacin A, a bacteriocin produced by lactobacillus curvatus strain LTH 1174, originating from fermented sausage. Appl Environ Microbiol 2004; 70: 2271-2278.
[28] Rajaram G, Manivasagan P, Thilagavathi B, Saravanakumar A. Purification and characterization of a bacteriocin produced by Lactobacillus lactis Isolated from marine environment. Adv J Food Sci Technol 2010; 2(2): 138-144.
10.12980/APJTB.4.2014C947
*Corresponding author: Baskar Balakrishnan Ph.D., Post-Doctoral Research Fellow, 108 Founders Hall, 820 Chestnut Street, Lincoln University of Missouri, Jefferson City, Missouri 65101, USA.
Tel: +1 573 821 3581
E-mail: baskarmicrobiologist@gmail.com, BalakrishnanB@lincolnu.edu
Fundation Project: Supported by the Project Drug from Sea, funded by Ministry of Earth Sciences, Govt. of India (Grant # 34030020005).
Article history:
Received 28 Dec 2013
Received in revised form 11 Jan, 2nd revised form 16 Jan, 3ed revised form 18 Jan 2014
Accepted 20 Feb 2014
Available online 28 Apr 2014
Methods:The bacterium was characterized by phenotype besides 16S rRNA gene sequence analysis. The antibacterial activity was recognised by using agar well diffusion method. The vibriocin was purified using ammonium sulphate precipitation, butanol extraction, gel filtration chromatography, ion-exchange chromatography and subsequently, by HPLC. Molecular weight of the substance identified in SDS-PAGE. Production optimization performed according to Taguchi’s mathematical model using 6 different nutritional parameters as variables.
Results:The objective bacterium was identified as Vibrio parahaemolyticus. The vibriocin showed 18 KDa of molecular mass with mono peptide in nature and highest activity against pathogenic Vibrio harveyi. The peptide act stable in a wide range of pH, temperature, UV radiation, solvents and chemicals utilized. An overall ~20% of vibriocin production was improved, and was noticed that NaCl and agitation speed played a vital role in secretion of vibriocin.
Conclusion:The vibriocin identified here would be an effective alternative for chemically synthesized drugs for the management of Vibrio infections in mariculture industry.
杂志排行

Asian Pacific Journal of Tropical Biomedicine的其它文章
- The effects of exposure to pesticides on the fecundity status of farm workers resident in a rural region of Fars province, southern Iran
- A cross sectional study on antibiotic resistance pattern of Salmonella typhi clinical isolates from Bangladesh
- Comparative susceptibility to permethrin of two Anopheles gambiae s.l. populations from Southern Benin, regarding mosquito sex, physiological status, and mosquito age
- Effects of melatonin on changes in cognitive performances and brain malondialdehyde concentration induced by sub-chronic coadministration of chlorpyrifos and cypermethrin in male Wister rats
- Phytochemical and in vitro biological investigations of methanolic extracts of Enhydra fluctuans Lour.
- Theoretical and experimental study on lipophilicity and wound healing activity of ginger compounds