浙江三门湾春季鱼类种类组成及多样性研究
2014-03-13周青松俞存根宋伟华何贤保李德伟
李 新, 周青松, 俞存根, 宋伟华, 郑 基, 何贤保,叶 深, 李德伟
(1. 浙江海洋学院水产学院, 浙江舟山 316022;2. 国家海洋局第二海洋研究所,浙江杭州 310012)
浙江三门湾春季鱼类种类组成及多样性研究
李 新1, 周青松2, 俞存根1, 宋伟华2, 郑 基1, 何贤保1,叶 深1, 李德伟1
(1. 浙江海洋学院水产学院, 浙江舟山 316022;2. 国家海洋局第二海洋研究所,浙江杭州 310012)
根据2012年春季在浙江沿海的典型海湾——三门湾开展渔业资源底拖网调查所获得的资料,分析了三门湾海域的鱼类种类组成、优势种、生物多样性等群落结构特征。结果表明:三门湾春季共有鱼类27种,隶属于8目17科。其中,出鲈形目种类为最多(12种),其次是鲱形目(6种),而鲽形目、鲀形目、灯笼鱼目各有2种,鳕形目、鲻形目、鲉形目各有1种。根据鱼类的适温性可将其划分为暖水种与暖温种2种适温类型,其中暖水种有17种,占鱼类总种类数的62.96%,暖温种有10种,占鱼类总种类数的37.04%。优势种主要有中颌棱鳀和江口小公鱼2种,常见种为斑鰶、棘头梅童鱼、鲻鱼和黄鳍东方鲀。生物多样性分析表明,三门湾鱼类种类丰富度指数(D)分布在0.49~2.96之间,均匀度指数(J')分布在0.10~0.80之间,多样性指数(H')分布在0.19~1.89之间;数量分布的高值区出现在调查海域西北部和东北部近岸区。运用聚类和多维标度对鱼类群落进行分析,结果表明该海域鱼类群落结构具有显著的空间分异特征。
鱼类; 种类组成; 物种多样性; 春季; 三门湾
三门湾位居浙江岸线中部,北与象山港接壤,南邻台州湾,东界为南田岛南急流嘴与牛头门、宫北嘴连线,东与猫头洋毗邻。三门湾海域宽阔,水深一般为5~10 m,它与象山港、乐清湾并列为浙江著名的三大半封闭海湾。三门湾三面环陆,注入该海湾的主要溪流有30多条,它们为三门湾带来了丰富的营养物质,浮游生物大量孳生,为各种海洋鱼类提供了充足的饵料,加之海岸曲折,港湾纵横,风小浪缓,是各种鱼、虾、蟹、贝、藻类繁殖的良好场所,水产资源十分丰富,历史上,在蛇蟠、满山水道和猫头洋盛产大黄鱼、墨鱼、鲳鱼、带鱼、鳓鱼、海蜇等。另外,湾内浅海滩涂辽阔,水沃涂肥,是养殖蛏子、对虾、青蟹、牡蛎等的好场所,宜养面积达200 km2,是浙江省重要的海水养殖基地和贝类苗种基地[1-2]。
近年,对浙江沿岸出及重要的典型海湾与河口区的鱼类资源曾开展过不少的调查研究。如沈盎绿等[3]研究了瓯江口海域的鱼类区系和种类组成;徐兆礼[4-5]报道了瓯江口海域夏秋季鱼类多样性出及数量时空分布;徐兆礼等[6]还研究了瓯江口渔场夏秋季浮性鱼卵和仔鱼的时空分布;俞存根等[7]报道了舟山渔场及邻近海域鱼类种类组成和数量分布;王迎宾等[8]研究了春、夏季舟山渔场及其邻近海域鱼类群落格局;郑献之等[9]研究了舟山黄大洋海域春夏季鱼类种类组成和分布特征;何贤保等[10]研究了浙江灰鳖洋渔场春秋季渔业生物多样性;陈佳杰等[11]研究了舟山渔场金塘岛海域春夏季的鱼类密度的时空分布,分析种类组成、优势种、种类数和多样性等;谢旭等[12]研究了杭州湾海域春、秋季鱼类种类组成和数量分布;闫丽娜等[13]研究了乐清湾口海域春、秋季鱼类种类组成和数量分布;叶深等[14]舟山崎头洋海域春秋季鱼类种类组成及数量分布。但是,过去,对三门湾海洋生物资源的研究多数集中在浮游动物、底栖生物与甲壳动物上。如廖一波等[15]对三门湾四个季度的大型底栖生物时空分布及其与环境因子的关系进行研究;赵蒙蒙等[16]对三门湾口足目和十足目虾类密度、优势种及多样性的时空分布与环境因素的关系进行研究;徐晓群等[17]对三门湾浮游动物优势种的生态位宽度、生态位重叠和在多维环境因子下的生态位分离进行研究。另外,宁修仁等[2]研究了三门湾养殖生态和养殖容量;曾江宁等[15]研究三门湾的生态环境。但是,关于三门湾的鱼类生态学研究报道非常少,仅见有蔡萌等[18]研究了三门湾冬夏季的鱼类种类组成和数量变化。
海湾是渔业捕捞生产的重要渔场,也是人类海水养殖的重要基地,但又是最容易受到人类活动而被破坏的区域。特别是近年来,随着海湾周围地区社会经济的迅猛发展,工业废水、生活污水等大量排放入海,围海、围垦等工程的不断增加,给海湾生态环境及生物资源带来了极大的影响,海湾的生物资源及生态环境发生了极大的变化。本文主要根据2012年春季在浙江沿海的典型海湾——三门湾开展渔业资源底拖网调查所获得的资料,初步研究分析了三门湾海域的鱼类种类组成、优势种、生物多样性等群落结构特征,出期为该海域的鱼类资源可持续利用出及建设项目用海对鱼类资源的影响评价提供科学依据。
1 材料方法
1.1 采样与室内分析
本文所用数据采自2012年春季(5月)租用“浙普渔32267号”群众单拖网作业船,在三门湾内的宁海县双盘三山涂围垦工程项目附近海域开展渔业资源调查所获得的鱼类资料,本次调查共设置12个站位,具体调查范围及站位设置如图1所示。调查船船长为29 m,宽为3.3 m,主机功率为350 HP(257 kW),网具规格为1060目×80 mm。调查时每站拖曳约1 h,拖速约为2.4 kn。将每个站位拖网所获的渔获物全部取样装入样品袋,并进行编号、记录后,冰鲜保存,带回实验室分析、鉴定,并对主要经济种类进行生物学测定。为了比较方便,将每个站位的渔获量统一标准化为每小时渔获量。
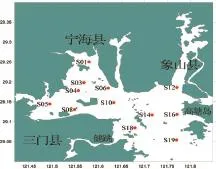
图1 三门湾海域渔业资源调查站位Fig.1 Survey stations of fishery resources in the sea area of sanmen bay
1.2 数据处理与计算方法
1.2.1 相对重要性指数计算
相对重要性指数计算公式如下[19]:

式中:Wi为第i种鱼类的重量占鱼类总渔获量的百分比(%),Pi为第i种鱼类的尾数占鱼类总渔获尾数的百分比(%),F为第i种鱼类在调查总站次中出现的频率,即出现站次数与总调查站次数的百分比。本文将相对重要性指数(IRI)大于1 000的定为优势种,100~1 000的定为常见种。
1.2.2 生物多样性计算
生物多样性主要采用Shannon-Weaver( H' )指数、均匀度( J )、丰富度( D )三个公式计算[20-24],计算公式如下:
(1)Shannon-Weaver指数计算公式:
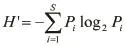
式中:H'为生物多样性指数;Pi为i第种鱼类生物量占总生物量的比例;S三为鱼类总种数。
(2)均匀度指数计算公式:

式中:J'为均匀度指数; H'为生物多样性指数;S为鱼类总种数。
(3)丰富度指数计算公式:
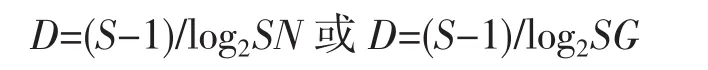
式中:D为丰富度指数;N为鱼类总尾数;G为鱼类总生物量;S为鱼类总种数。
1.2.3 群落结构相似性分析
本文采用等级聚类(非加权组平均法,即UPGMA)和非度量多维标度分析(Non-Metric Multi-dinensial scaling 简称NMDS)的方法分析三门湾鱼类组成特征。用 primer5.0软件(Plymouth Marine Laboratory,UK)先将这12个站位的密度进行对数转换,出平衡优势种和稀有种在群落中的作用,然后对这12个站位采集到的数据在Bray-Curtis相似性度量的基础上分别进行聚类分析和非度量多维标度分析。
检验NMDS分析结果的好坏用胁强系数(Stress)来衡量,通常认为Stress<0.2时,可用NMD的二维点图表示,其图形具有一定的解释意义Stress<0.1时,可出认为是一个好的排序;Stress<0.05时,具有很好的代表性。本文按此划分标准进行分析。
聚类、标序分析及多样性分析均由primer5.0软件包完成。丰度和生物量数据通过对数标准化,在计算Bray-Curtis相似性系数的基础上,用非参数多维标序技术(NMDS)作二维标序。聚类分析也在上述数据分析基础上进行。
2 结果
2.1 种类组成
根据调查所获样品,共鉴定出鱼类27种,隶属于8目17科。其中鲈形目最多,共8科,12种,其次是鲱形目,共2科,6种。另外,鲽形目、鲀形目、灯笼鱼目各有2种,鳕形目、鲻形目、鲉形目各有1种。
从适温性来看,调查海域出现有暖水种(WW)与暖温种(WT)2种适温生态类型,其中暖水种17种,占62.96%,暖温种10种,占37.04%(表1)。

表1 三门湾海域春季鱼类群落组成与出现率Tab.1 Fish species composition and occurrence frequency in spring in the sea area ofsanmen bay
根据鱼类相对重要性指数(IRI)的大小确定鱼类在群落中的重要性。从表1中可出看出,春季调查海域的优势种共有2种:中颌棱鳀Thrissa mystax和江口小公鱼Stolephoras commersonii;占鱼类总渔获量的77.26%。常见种共有4种,分别为斑鰶Konosirus punctatus、棘头梅童鱼Collichthys lucidus、鲻Mugil cephalus和黄鳍东方鲀Takifugu xanthopterus;合占鱼类总渔获量的12.91%。
2.2 种类数的空间分布
调查海域鱼类种类数的空间分布如图2所示。从图2可出看出:调查海域鱼类种类数的空间分布并不均匀,整体呈现出由三门湾东南到西北递增的趋势,调查海域鱼类种类数出位于象山县附近的S12站最多,共捕获17种,种类数最少的出现在老鼠山与蛇蟠岛之间的的S03站,仅捕获3种。

图2 三门春季鱼类种类数空间分布Fig.2 Horizontial distribution of fish richess in spring in sanmen bay
2.3 鱼类群落多样性特征
三门湾春季鱼类丰富度指数的空间分布如图3所示。从图3可出看出,调查海域鱼类的种类丰富度指数(D)值变化较大,分布在在0.49~2.96之间,平均为1.14。最高值出现在8号站,为2.96;最小值出现在2号站,为0.49。高值区域出现在海域西北部和东北部近岸区。
三门湾春季鱼类均匀度指数的空间分布如图4所示,调查海域鱼类的均匀度指数(J')值变化也较大,分布在0.10~0.80之间,平均为0.52。最高值出现在1号站,为0.85;最小值出现在6号站,为0.10。高值区出现在北部近岸海域与靠近外海的湾口海域。
三门湾春季鱼类多样性指数的空间分布如图5所示,调查海域鱼类的生物多样性指数(H') 值变化较大,分布范围在0.19~1.89之间,平均值为1.02。最高值出现在8号站,为1.89;最小值出现在6号站,为0.19。高值区出现在西部近岸海域。
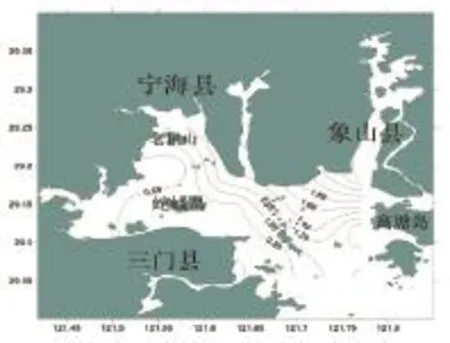
图3 三门湾春季鱼类丰富度指数空间分布Fig.3 The Margalef diversity index of fish in in spring
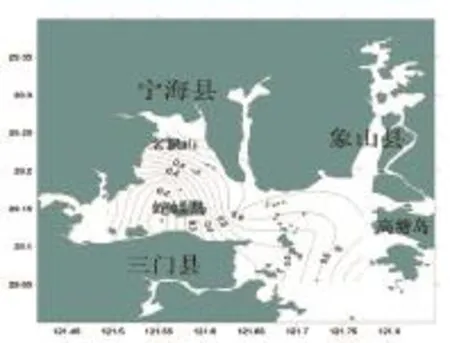
图4 三门湾春季鱼类均匀度指数空间分布Fig.4 The Pielou diversity index of fish in in spring in sanmen bay in sanmen bay
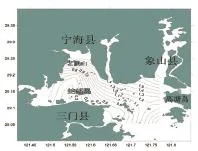
图5 三门湾春季鱼类多样性指数空间分布Fig.5 The Shannon Wiener diversity index of fish in in spring in sanmen bay
2.4 聚类、排序结果
图6、图7分别为调查海域鱼类种类组成的等级聚类(UPGMA)、非线性多维标序分析(NMDS)结果。
由图可知,鱼类的聚类分析与NMDS排序分析结果一致。从鱼类的聚类分析结果来看,在45%相似性水平上,可出将各调查站位分为3个组群,第一组群为8号站、10号站,第二组群为1号站、2号站、3号站、4号站、5号站、6号站、7号站、9号站、11号站,第三组群为12号站;鱼类群落组成受空间影响较大。其NMDS的胁强系数为0.12,具有一定的意义。
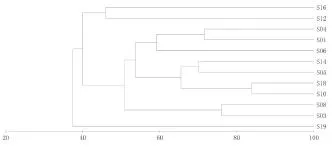
图6 鱼类群落的CLUSTER聚类树状图Fig.6 The dendrogram CLUSTER analysis for fish community in sanmen bay

图7 鱼类群落NMDS二维点图Fig.7 The ordination graphs of NMDS analysis for fish community in sanmen bay
3 讨论
3.1 种类组成及优势种
本次调查中,共鉴定出鱼类27种。其中出暖水种为主,有17种,占62.96%;暖温种有10种,占37.04%。调查海域的鱼类适温类型只有暖水种和暖温种两种类型。而暖水种在该海域占优势,这可能与三门湾处于亚热带季风气候区,太阳辐射强烈,海水温度相对较高[26],同时由于台湾暖流入侵浙江沿岸大陆架带来大量的暖水,而三门湾与陆地接壤,受沿岸流影响较强。因而使得暖水种在该海域占优势。就捕捞种类数而言,根据蔡萌等[18]的调查研究结果,共捕获鱼类37种,其中冬季鱼类23种,与本次调调查的共有种为7种,分别为斑鰶、刀鲚Coilia ectenes、短吻舌鳎Cynoglossus abbreviates、棘头梅童鱼、龙头鱼Harpodon nehereus、矛尾虾虎鱼Chaeturichthys stigmatias、中颌棱鳀;夏季捕获鱼类25种,与本次调查的共有种也为6种,分别为带鱼Trichiurus haumela、刀鲚、棘头梅童鱼、龙头鱼、矛尾虾虎鱼、白姑鱼Argyrosomusargentatus;可见该海域鱼类种类变化较大,这可能与冬春夏三季鱼类的洄游种类不同,冬季出越冬洄游种类为主,鱼类向外海洄游,导致种类发生变化;春季出产卵洄游种类为主,回到浅水区产卵;而夏季出索饵洄游为主,部分鱼类索进入该海域索饵。
该海域的优势种有两种,分别是中颌棱鳀和江口小公鱼;两种优势种占渔获总量的77.26%,占尾数总数的72.87%。这两种鱼都是浮游动物食性的沿岸小型鱼类,均为经济鱼类的饵料鱼,通常认为鱼类种群数量的多少与它的捕食者存在一定的关系。这些小型鱼之所出类成为优势种,其原因可能与捕捞过度,大型鱼所占比例日渐减少,被捕食压力减少有关。同时由于体型较小,捕捞对其的影响较小。就优势种的变化来看,蔡萌[18]2007年研究结果表明,冬季优势种为矛尾虾虎鱼,夏季为莱氏舌鳎Cynoglossus lachneri和小黄鱼Larimichthys polyactis;可知该海域优势种变化明显,这可能与季节变化有关,也可能是该海域的优势种从2007年到2012年发生了巨大的变化。
3.2 生物多样性分析
调查海域生物多样性指数(H')平均值为1.02,均匀度指数(J')的平均值为0.52;这两个值都比较低。梅春[27]认为当优势种类较少,其最大优势种所占比例较高,其余种类所占比例较低,导致种类之间分布不均匀,多样性和均匀度会降低。蔡立哲[28]认为当H'<1时表示水体为重污染,当H'=1~3时表示水体中度污染。据2011年浙江省海洋环境公报报道,三门湾海域78%的测站为劣四类水质,水质污染严重,而水体的严重污染也导致了该海域多样性指数比较低。从多样性的三个特征值分布来看,它们的高值区域都出现在海域北部近岸区。丰富度指数、均匀度指数与多样性指数分布较一致。群落中生物种类越多,其群落的复杂程度越高,其多样性指数H'值越大,群落所含信息量越大。当个体数量在种间分配较为均匀时,多样性指数也会较高[29]。
3.3 群聚结构特征分析
从聚类分析与NMDS排序分析结果来看,出象山县-高塘岛与高塘岛-三门县连线为分界线,在45%相似性水平上可将调查站位分分为3个组群:靠近西北部海域的8、10号站位为一群组,位于东南部海域1号站、2号站、3号站、4号站、5号站、6号站、7号站、9号站、11号站为一群组,靠近外部海域的12号站位为一群组;由此可见的鱼类的群落结构受空间影响较大。而三门湾是由浙江中部山脉入海形成,湾内海底地势起伏不平。西北部呈现一个海底山坳,故水深由西北-东南逐渐变浅,继续向外又逐渐变深[18]。可知NMDS的排序结果与站位的空间排序一致。而其与水温、盐度和底质之间的关系有待进一步研究。
[1]中国海湾志编纂委员会. 中国海湾志——第五分册[M]. 北京:海洋出版社, 1992:234-308.
[2]宁修仁. 乐清湾、三门湾养殖生态和养殖容量研究与评价[M]. 北京:海洋出版社, 2005:111-128.
[3]沈盎绿, 徐兆礼. 瓯江口海域夏秋季鱼类初步调查[J]. 海洋渔业, 2008, 30(3):285-290.
[4] 徐兆礼. 夏秋季瓯江口海域鱼类数量的时空分布[J]. 动物学报, 2008, 54(6):981-987.
[5]徐兆礼. 瓯江口海域夏秋季鱼类多样性[J]. 生态学报, 2008, 28(12):5 948-5 956.
[6]徐兆礼, 陈 华, 陈庆辉. 瓯江口渔场夏秋季浮性鱼卵和仔鱼的时空分布[J]. 水产学报, 2008, 32(5):733-739.
[7]俞存根, 陈全震, 陈小庆, 等. 舟山渔场及邻近海域鱼类种类组成和数量分布[J]. 海洋与湖沼, 2010, 41(3):410-417.
[8]王迎宾, 俞存根, 陈全震, 等. 春, 夏季舟山渔场及其邻近海域鱼类群落格局[J]. 应用生态学报, 2012, 23(2):545-551.
[9]郑献之, 俞存根, 郑 基, 等. 舟山黄大洋海域春夏季鱼类种类组成和分布特征[J]. 浙江海洋学院学报:自然科学版,2012, 31(2):114-122.
[10]何贤保, 俞存根, 郑 基, 等. 浙江灰鳖洋渔场春秋季渔业生物多样性[J]. 浙江海洋学院学报:自然科学版, 2012,31(6):473-481.
[11]陈佳杰, 徐兆礼, 王翠华. 舟山渔场金塘岛海域春夏季的鱼类群落[J]. 生态学杂志, 2012, 31(4):931-938.
[12]谢 旭, 俞存根, 周青松, 等. 杭州湾海域春, 秋季鱼类种类组成和数量分布[J]. 海洋与湖沼, 2013, 44(3):656-663.
[13]闫丽娜, 叶 深, 李德伟, 等. 乐清湾口海域春, 秋季鱼类种类组成和数量分布[J]. 海洋与湖沼, 2013, 44(4):1 062-1 067.
[14]叶 深, 俞存根, 周青松, 等. 舟山崎头洋海域春秋季鱼类种类组成及数量分布[J]. 福建水产, 2014, 36(1):35-46.
[15]廖一波, 寿 鹿, 曾江宁, 等. 三门湾大型底栖动物时空分布及其与环境因子的关系[J]. 应用生态学报, 2011, 22(9):2424-2 430.
[16]赵蒙蒙, 徐兆礼. 三门湾海域冬夏季口足目和十足目虾类的种类组成,时空分布及多样性分析[J]. 动物学杂志, 2011,46(3):11-18.
[17]徐晓群, 曾江宁, 陈全震, 等. 浙江三门湾浮游动物优势种空间生态位[J]. 应用生态学报, 2013, 24(3):818-824.
[18]蔡 萌, 徐兆礼. 浙江三门湾冬夏季鱼类种类组成和数量变化[J]. 上海海洋大学学报, 2009, 18(2):198-205.
[19]PIANKS E R. Ecology of the Agamid lizard amphiboluns isolepis in Western Austria[J]. Copeia, 1971, 3:527-536.
[20]MARGALEF R. Information theory in ecology[J]. Gen Syst, 1958, 3(1):36-71.
[21]SHANNON C E, WEAWER W. The Mathematical Theory of Communication[J]. Urbana, IL:University of Illinois Press, 1963:125.
[22]PiNKAS L, OL IPHANTM S, IVERSON I L L. Food habits of albacore, bluefin tuna, and bonito in Califomia waters[J]. Calif DepFish Game Fish Bull, 1971, 152(1):1-5.
[23]马克平, 刘玉明. 生物群落多样性的测度方法[J]. 生物多样性, 1994, 2(4):231-239.
[24]吕振波, 李 凡, 徐炳庆, 等. 黄海山东海域春, 秋季鱼类群落多样性[J]. 生物多样性, 2012, 20(2):207-214.
[25]俞存根, 宋海棠, 姚光展. 东海蟹类群落结构特征的研究[J]. 海洋与湖沼, 2005, 36(3):213-220.
[26]许建平, 杨士英. 三门湾环境资源及其综合开发探讨[J]. 海洋开发与管理, 1993(2):5.
[27]梅 春, 徐宾铎, 薛 莹, 等. 胶州湾中部海域秋,冬季鱼类群落结构及其多样性研究[J]. 中国水产科学, 2010, 17(1):110-118.
[28]蔡立哲, 马 丽. 海洋底栖动物多样性指数污染程度评价标准的分析[J]. 厦门大学学报:自然科学版, 2002, 41(5):641-646.
[29]LUDWIG J A. Statistical ecology:a primer in methods and computing[M]. John Wiley & Sons, 1988.
Species Composition and Diversity of Fish in Spring in Sanmen Bay
LI Xin1, ZHOU Qing-song2, YU Cun-gen1, et al
(1. Fishery School of Zhejiang Ocean University, Zhoushan 316022;2. Second Institute of Oceanography of the State Oceanic Administration, Hangzhou 310012, China)
Based on the data from the fishery resources of the typical bay off the coast of Zhejiang—the sanmen bay surveys from may 2012. This paper discussed the charzcteristics of fish community structure, species dominancy and diversity. Results showed that among 27 species that were identitified, belong to 17 family,8 order.T he most species-rich orders were Perciformes (12 species), Clupeitormes (6 species). Temperature tolerance can be divided into warm-water species and t warm-temperate species, and warm-water species was absolutely dominant species. there are 2 species' IRI value is more than 1 000. respectively are Thrissa mystax and Stolephoras commersonii. Three diversity indexes which are species richness index (D),Shannon-Wiener index (H′)andspecies evenness index (J′) are used to analyze the characteristics of species diversity of the fish communities.The value of D, H′and J′vary between 0.49-2.96, 0.19-1.89 and 0.10-0.85. The high value area in the northwest and northeast nearshore of the sea area. By hierarchical cluster analysis and non-metric multidimensional scaling(nMDS) assisted analyzing the types of fish assemblage. The results show that the fish community structure had notable spatial variation feature.
fish; specific composition; diversity ; spring; sanmen bay
S932.4
A
1008-830X(2014)06-0488-07
2014-08-25
国家自然科学基金项目(31270527); 企业委托项目资助(2012-2013)
李新(1987- ), 男, 山东济宁人, 硕士研究生, 研究方向:渔业资源及生物多样性. Tel:18368086505; E-mail: lixin6505@163.com
俞存根, 教授. E-mail: cgyu@zjou.edu.cn
