夏季南大洋南极大磷虾种群分布、结构及生长研究
2014-03-08时永强孙松李超伦陶振铖
时永强 孙松 李超伦 陶振铖
(1中国科学院海洋研究所海洋生态与环境科学重点实验室,山东青岛266071;2中国科学院大学,北京100049;3中国科学院海洋研究所胶州湾海洋生态系统研究站,山东青岛266071)
0 引言
南极磷虾在南大洋食物网中处于中枢位置,既是自养及异养浮游生物的重要捕食者,又是许多高营养级动物的主要饵料,在南极海洋生态系统中起着承上启下的重要作用[1]。生活在南极南大洋海域的磷虾通称为南极磷虾,但人们所讲的南极磷虾通常指的是南极大磷虾。南极大磷虾在南大洋环南极海域均有分布,但分布不均匀,超过50%磷虾分布于西南大西洋[2-4]。南极大磷虾丰度降低会对它的捕食者造成很大影响[5]。近些年来,大量研究结果表明,伴随气候变暖海冰范围减小,磷虾丰度呈现下降趋势,纽鳃樽数量上升,进而会影响到海洋食物网乃至整个南大洋生态系统[3,5]。
南极大磷虾作为地球上最大的单种生物资源,其现存量为6.5—10亿吨,被认为是地球上最大、也是最后一个蛋白库[6],无论从哪个角度来衡量,都是我们这颗星球上数量最大、繁衍最成功的物种之一,吸引着人类进行渔业捕捞及开发利用[6-7]。对于中国这样一个海洋生物资源相对贫乏的人口大国来讲,南极资源的调查和开发利用具有重要战略意义。中国从20世纪80年代开始南大洋的科学研究工作,在南极半岛及普里兹湾海域开展了南极大磷虾部分生态学研究,包括生殖特点、种群结构、年龄组成、丰度变化及生长状况等研究[8-13],但对于环南极大磷虾的生长状况及种群结构方面研究较少。
本研究利用中国第24次(2007/2008年)和第25次(2008/2009年)南极科学考察的网采磷虾样品,结合同期物理环境资料,分析了南大洋南极大磷虾分布、丰度、生长状况及种群结构,以期为南极海洋生态系统研究及中国南极磷虾资源开发利用提供基础理论和科学依据。
1 材料和方法
1.1 站位及采样
2008年1月2日—3月15日在南大洋50°S以南海域进行了环南极走航采样,调查经度范围为64.95°W—80.61°E。2009年 1月 11日—3月 13日在南大洋普里兹湾及其邻近海域进行了走航采样,调查经度范围为70.47°E—141.84°E。
两次考察均利用高速采集器(High Speed Collector)和 IKMT网(Isaac-Kidd Midwater Trawl)两种网具采集南极大磷虾。高速采集器(网口直径70 mm,网目350μm)在船舶正常航渡期间,每隔6 h采样一次,使用绞车放缆400 m,然后稳定拖曳20 min,最后低速(速度约 0.3 m·s-1)收网。IKMT网(网口面积2 m2,网目6 mm)在船速维持3—4节时拖网取样,放缆300 m,拖曳20 min,收网速度小于0.3 m·s-1,主要根据渔探仪映像进行定点斜拖采样。以上两种网具均主要进行水深<50 m的斜拖采样,采集50 m水深以内的整个表层和次表层磷虾样品。采样站位见图1。
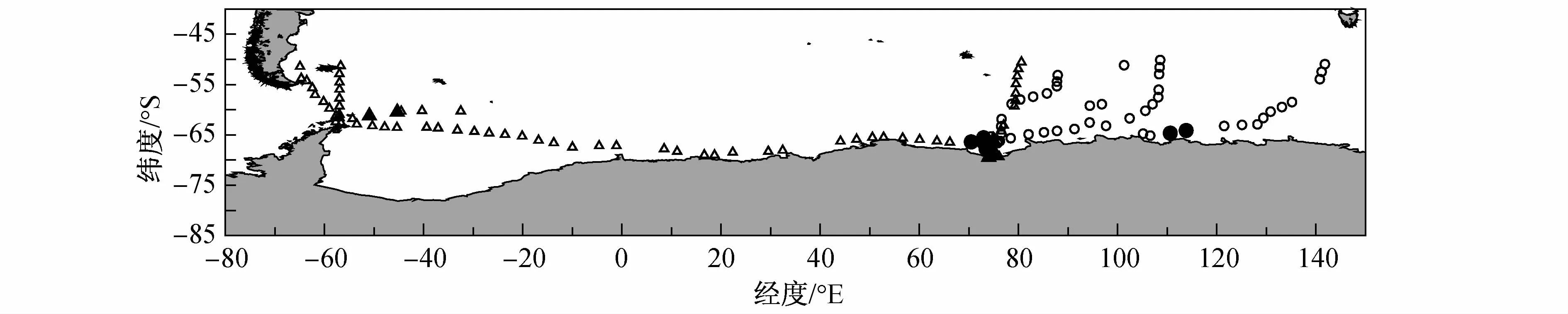
图1 采样站位图.△—2008年高速采集器采样站位;▲—2008年IKMT网采样站位;○—2009年高速采集器采样站位;●—2009年IKMT网采样站位Fig.1.Sampling stations.△—stationswith High Speed Collector in 2008;▲—stations with IKMT in 2008;○—stations with High Speed Collector in 2009;●—stations with IKMT in 2009
1.2 样品处理
南极磷虾样品均用甲醛固定,最终浓度为5%。航次结束后,在实验室中对磷虾样品进行鉴定、测量和分析。对每个站位南极大磷虾样品(包括伤残和完整磷虾样品)进行计数,并对其总重(湿重)进行测量(精确到0.001 g)。高速采集器及IKMT网的滤水体积由各自网口面积乘以相应船速和拖网时间获得。每个站位南极大磷虾的丰度和生物量由该站样品中大磷虾总个数和总湿重除以滤水体积获得,单位分别为 ind·1 000 m-3和 g·1 000 m-3。
对于磷虾数量较多的站位样品,从样品中随机挑出30尾个体完整的大磷虾成体进行体长测定、眼径测定和性期分析。对于磷虾数量相对较少的站位,测量和分析全部个体完整的大磷虾样品。南极大磷虾的体长根据Mauchline[14]的标准1:由大磷虾额角尖端到尾肢的最末端用游标卡尺进行测量,测量精度为0.02 mm。取下大磷虾左眼,在解剖镜下利用目镜测微尺测量大磷虾复眼眼径,测量精度为0.05 mm。大磷虾发育期根据 Makarov等[15]的标准在解剖镜下鉴定分析,没有明显雌雄体分类特征的个体划分为未成体(Juvenile)。雌性划分依据如下:次成体(sub-adult)不分期,以2F表示;成体(adult)分五期,3AF为未交配雌性成体,3BF为交配的雌性成体,3CF为卵巢充满胸部的雌性成体,3DF为临产雌性成体,3EF为产后雌性成体。雄性划分依据为:次成体分三期,2AM—2CM表示交接器在发育中的雄性次成体;雄性成体分两期,3AM为贮精囊中尚无精荚的成熟雄性成体,3BM为贮精囊中有精荚存在,随时可以交配的雄性成体。
1.3 环境因子测量
在每个站位走航采样过程中同步进行温度和表层叶绿素a浓度测定。采样深度和温度利用小型自容式温度深度剖面测定仪(COMPACT-TD,型号:ATD-HR,JFE ALEC公司)同步测得。用于测定叶绿素a(chlorophyll a)浓度的海水直接采自表层,水样采集后,将500 ml水样过滤到Whatman GF/F玻璃纤维滤膜(0.70μm)上,然后立即将滤膜保存在-20℃冰箱内。叶绿素的测定先经90%丙酮溶液在≤4℃条件下萃取24 h,然后用Turner Designs荧光计测定叶绿素a浓度的含量。每10 d一次的海冰照片由美国国家海洋大气局(NOAA)提供。
2 结果
2.1 环境因子
采集到大磷虾站位的采样时间、表层海水温度及表层叶绿素a浓度,见表1(G代表高速采集器获取样品,K代表IKMT网获取样品)。2008年除1月3日的K24-01站(0.56℃)、1月4日的 K24-02站(2.28℃)和2月9日的 G24-01站(1.86℃)外,2月10—20日环南极走航采样站位中,表层水温均低于0℃。在2009年普里兹湾及其邻近海域调查中,大多数站位表层水温则高于0℃。2008年夏季采集到大磷虾站位的表层叶绿素a浓度范围是0.09—0.65μg·L-1,其中叶绿素 a浓度最大值出现在威德尔海区的K24-01和G24-02站。2009年夏季采集到大磷虾站位的表层叶绿素a浓度范围是0.13—3.09μg·L-1,其中 G25-03和 G25-04站叶绿素a浓度均超过1.50μg·L-1,位于普里兹湾东侧邻近海域。在普里兹湾中,采集到大磷虾的站位表层叶绿素 a浓度范围是0.13—0.80μg·L-1。
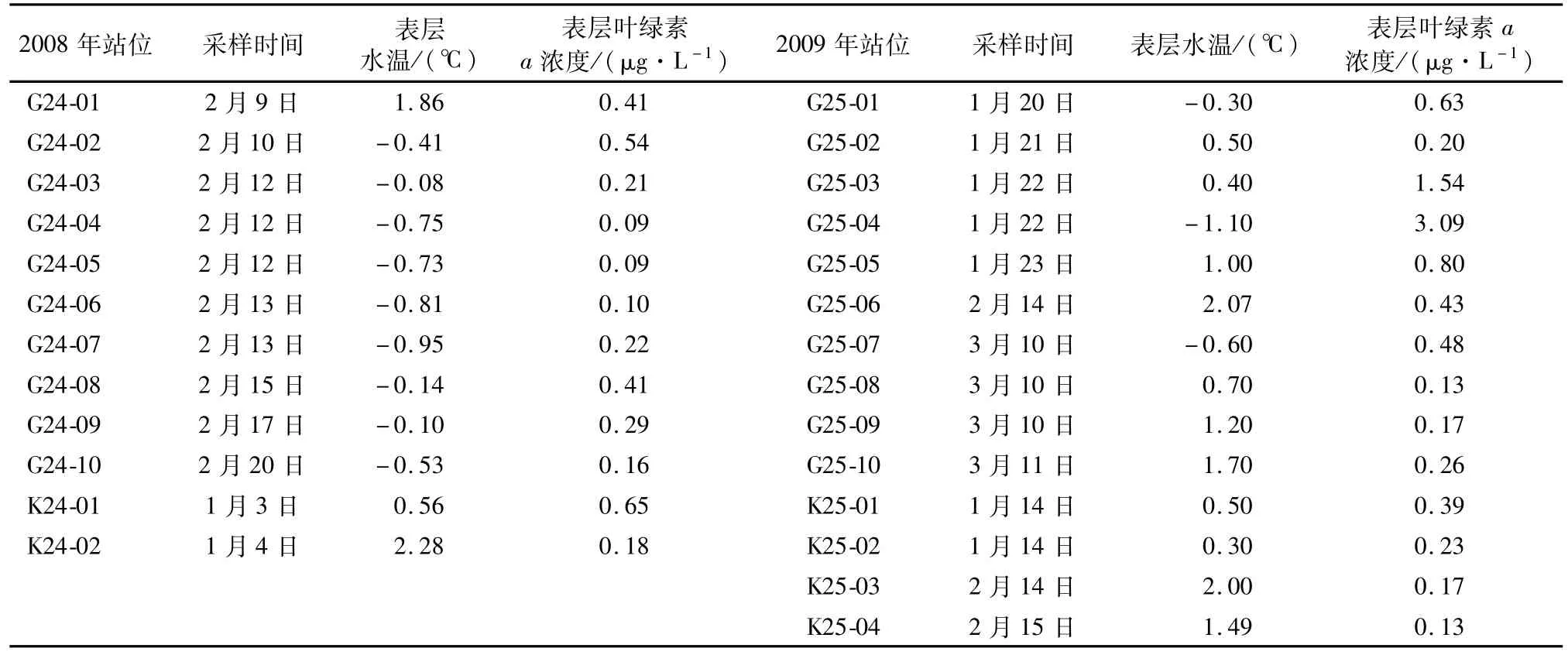
表1 2008年和2009年夏季采集到南极大磷虾站位的采样时间、表层水温度及叶绿素a浓度Table 1.Sampling time,surface seawater temperature and chlorophyll a concentration atstationswhere Euphausia superba was found in summer 2008 and 2009
根据美国国家海洋大气局提供的每10 d一次的海冰照片,对2008年和2009年的1—3月普里兹湾海域的海冰发展情况进行比对,结果显示,2009年海冰回退时间较早,且冰缘线更靠近近岸。以航次中采集到大磷虾量较多的站位中(依据结果2.2和2.3)最早的采样时间为依准,将采样位置与采样时间提前至少一个月的海冰消长情况的相对位置进行比对(表 2、3)。2007/2008年夏季,K24-01、K24-02和G24-09站在采样时间前有近一个月时间位于海冰边缘或冰缘外,而G24-03、G24-07和G24-10站在采样前站位未被海冰覆盖的时间较短。2008/2009年夏季,除K25-01和K25-02站采样前未被海冰覆盖的时间较短外,其余站位均有超过一个月时间未被海冰覆盖。
2.2 南极大磷虾的分布和丰度
在2007/2008年夏季的南极考察走航调查中,在纬度高于50°S的南大洋海域共获得66个高速采集器样品,其中仅有10个站位采集到发育期不低于未成体的南极大磷虾,在获得的5个IKMT网样品中,仅在威德尔海域的2个站位采集到大磷虾(图2)。由于高速采集器与IKMT网这两种网具存在差异,因此在分析种群丰度和分布时需要区别对待。在调查海域,南极大磷虾斑块状分布明显。利用高速采集器获取的磷虾样品中,南极大磷虾的丰度为25.7—2 428.4 ind·1 000 m-3,生物量为 4.3—921.3 g·1 000 m-3,样品量最大的两个站位是G24-09和G24-02,大磷虾密度均超过2 200 ind·1 000 m-3和700 g·1 000 m-3,另外有5个站位的大磷虾丰度小于500 ind·1 000m-3。IKMT网采集到南极大磷虾样品的两个站位是K24-01和K24-02,大磷虾的丰度分别为 28.9 ind·1 000 m-3和7.8 ind·1 000 m-3。从海区来看,普里兹湾海区未捕获到大磷虾,威德尔海的大磷虾丰度较高,并且在30°E—50°E陆架边缘,大磷虾捕获量也较多。

表2 2007/2008年夏季调查采集到南极大磷虾站位与海冰边缘的相对位置关系Table 2.Relative positional relationship between sea ice and stations recording Euphausia superba in summer 2007/2008
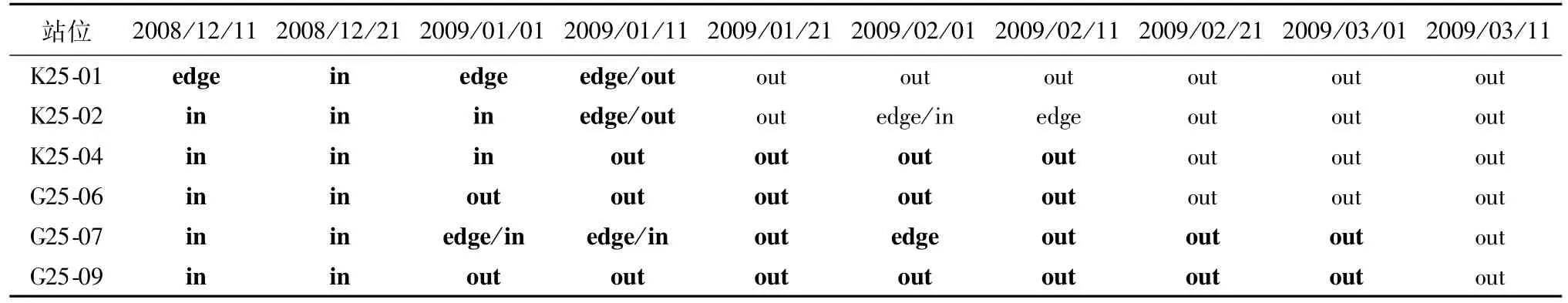
表3 2008/2009年夏季调查采集到南极大磷虾站位与海冰边缘的相对位置关系Table 3.Relative positional relationship between sea ice and stations recording Euphausia superba in summer 2008/2009
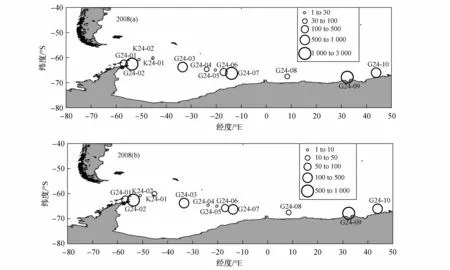
图2 2008年夏季南极大磷虾的种群丰度、分布和生物量.(a)丰度(ind·1 000 m-3);(b)生物量(g·1 000 m-3)Fig.2.Spatial distribution,abundance and biomass of Euphausia superba in summer 2008.(a)Abundance(ind·1 000 m-3);(b)Biomass(g·1 000 m-3)
2008/2009年夏季调查海域主要集中在普里兹湾及其以东海域,在纬度高于50°S的南大洋海域共获得44个高速采集器样品,其中10个站位采集到南极大磷虾,获取的6个IKMT网样品中有4个捕获到大磷虾(图3)。高速采集器获取的大磷虾样品中,丰度范围为17.4—949.4 ind·1 000 m-3,生物量范围为15.3—693.0 g·1 000 m-3。南极大磷虾在普里兹湾站位捕获量相对较高,丰度最大的两个站位G25-06和G25-09均位于普里兹湾海域,丰度超过750 ind·1 000m-3。IKMT网获取的大磷虾丰度范围是0.1—8.0 ind·1 000 m-3,且丰度最大的站位K25-04同样位于普里兹湾海域。
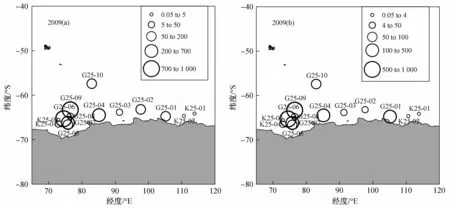
图3 2009年夏季南极大磷虾的丰度、分布和生物量.(a)丰度(ind·1 000 m-3);(b)生物量(g·1 000 m-3)Fig.3.Spatial distribution,abundance and biomass of Euphausia superba in summer 2009.(a)Abundance(ind·1 000 m-3);(b)Biomass(g·1 000 m-3)
2007/2008年和2008/2009年夏季的调查结果比较来看,无论高速采集器还是IKMT网,威德尔海中南极大磷虾最大丰度都要高于普里兹湾海域。
2.3 南极大磷虾生长状况及种群结构
高速采集器由于网口面积较小,每网捕获的大磷虾个数较IKMT网少很多,并且两种网对磷虾损伤均较大,将每个站位采集的磷虾样品中个体相对完整的大磷虾挑出,测定其性期、眼径及体长,仅对测量磷虾个数大于8尾的网样进行分析,并做大磷虾生长状况图(图4)及发育期组成百分比分布图(图5)。磷虾的“负生长(negative growth)”现象是指磷虾成体在饥饿状态下仍能正常蜕皮,但是每蜕一次皮体长就要缩短一些(负生长),形成一种独特的生长方式,以适应比较恶劣或者食物匮乏的外界环境[16]。Sun等[17-18]首次提出了利用南极大磷虾复眼晶椎数目和复眼直径测定磷虾生长状况的方法,进而确定环境变化对磷虾种群的影响,并可利用大磷虾生长状况指示环境的年际变化[19-20]。Sun等[18]指出盛夏南极大磷虾在正常的生长状况下眼径与体长存在明显的指数关系:
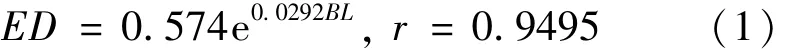
其中ED为眼径,BL为体长,单位均为mm,测量的大磷虾样品包括未成体、雌性成体及雄性成体。以该曲线为标准曲线,若磷虾发生负生长,则代表ED值的点将位于图中标准曲线的上方,而如果磷虾生长状况较好,则会接近标准曲线,或者位于标准曲线之下。图4给出各个站位南极大磷虾眼径体长状况与标准曲线的关系。从图中可以看出,在2007/2008年夏季调查中,大磷虾的生长状况总体上保持正常,但K24-02及G24-02站大磷虾的生长状况较差,可能出现了负生长,其他各站大磷虾生长状况较好。在2008/2009年夏季调查中,总体上磷虾生长状况类似于2007/2008年夏季,而K25-02和G25-09站大磷虾生长状况较差,其他各站大磷虾生长状况较好。
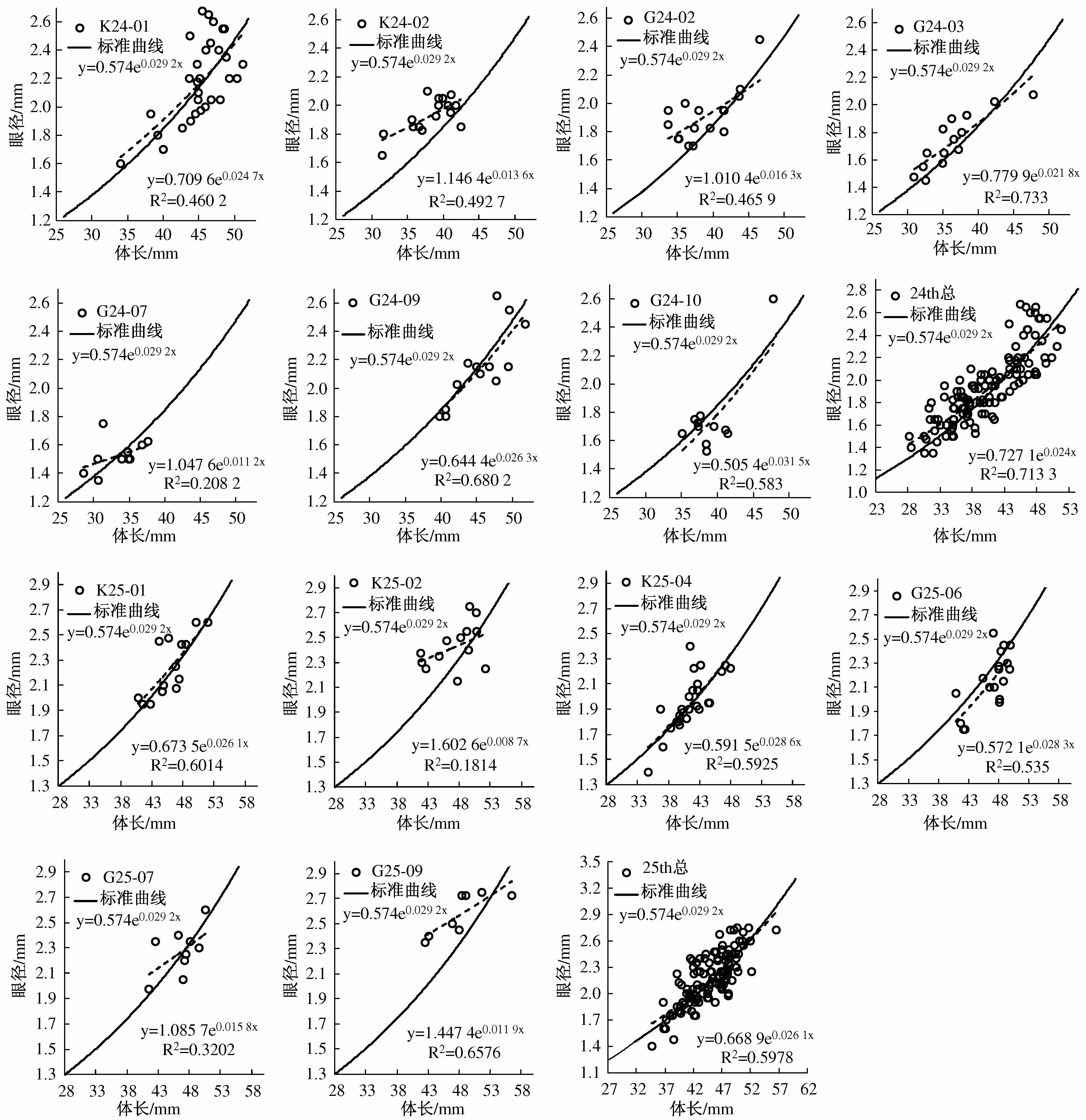
图4 2007/2008年和2008/2009年夏季南极大磷虾的生长状况Fig.4.Growth condition of Euphausia superba in summer 2007/2008 and 2008/2009
南极大磷虾发育期组成百分比分布情况见图5,从图中可见在2007/2008年夏季考察中,G24-03、G24-07和G24-10站未成体大磷虾所占比例均超过85%,其中G24-07站获取磷虾全部为未成体。G24-02站大磷虾发育期仅达到次成体,临产雌体(3DF)大磷虾仅出现在G24-09和K24-02站,其中3DF在G24-09站比例超过45%。雄性成体大磷虾在K24-01站占比例达50%,在G24-09站也有捕获。在2008/2009年夏季考察中,雄性成体(3AM)大磷虾在所有站位中均存在,且在G25-09、K25-01和K25-02站有较高比例分布;3BM大磷虾在K25-01站中占较高比例,另外在除G25-07站以外站位中均有捕获;临产雌体(3DF)大磷虾除在G25-09站未捕获到以外,其余各站均有捕获,并且在G25-06、K25-01和K25-02站均占较高比例。总体来看,2008/2009年夏季南极大磷虾的成熟度较高。

图5 2007/2008年和2008/2009年夏季南极大磷虾幼体后期发育期组成百分比分布.n—测量的磷虾尾数Fig.5.Percentage distribution of Euphausia superba in post-larval sexualmaturity stage in summer2007/2008 and 2008/2009.n—the number of krillmeasured
3 讨论
研究认为南大西洋南极磷虾丰度要远远高于南印度洋[2-4,21]。本研究中,2007/2008年利用高速采集器及IKMT网在普里兹湾的多个站位均未采集到南极大磷虾,而在威德尔海海域两种网具均采集到丰度较高的大磷虾。2008/2009年夏季考察主要集中在普里兹湾及其以东海域,普里兹湾海域使用IKMT网站位获取的南极大磷虾平均丰度为2.02 ind·1 000 m-3,而2007/2008年夏季在威德尔海域同样网具获取的大磷虾平均丰度为18.33 ind·1 000 m-3,要远高于普里兹湾的大磷虾丰度。因此,本研究也证实了南极大磷虾在威德尔海域丰度及捕获率都高于普里兹湾海域,然而2007—2009年在普里兹湾的两次调查,也同样显示了南极大磷虾的丰度具有一定的年际变化[5,22-23]。Atkinson等[3]指出,1976年以后西南太平洋海区南极磷虾丰度有明显的下降趋势。通过对比往年环南极走航利用高速采集器捕获的大磷虾丰度历史数据[24],可以发现1992/1993年和1997/1998年夏季大磷虾的捕获率及丰度要高于1999/2000年和本研究中的2007/2008年。2000年和2002年1月在普里兹湾利用IKMT网获得的调查海区的南极大磷虾平均丰度分别为 16.17 ind·1 000 m-3[8]和 68.85 ind·1 000 m-3[10],均高于本研究中 2008/2009年获取的平均丰度2.02 ind·1 000 m-3。大磷虾丰度可能受到现场调查的采样时间及磷虾斑块状分布特征影响,但整体上看,2007/2008年和 2008/2009年夏季大磷虾的丰度要低于历史同期。
不同的地理海域,由于生存环境和饵料等一系列因素的影响,导致大磷虾的生长状况有所不同。南极大磷虾“负生长”的生存策略可以适应条件恶劣或者食物匮乏的外界环境[16]。从本研究两年夏季调查的结果来看,大部分站位的大磷虾保持基本正常的生长状况,但在 K24-02、G24-02、K25-02和G25-09站大磷虾生长状况较差。K24-01和K24-02站采样位置与海冰边缘发展的相对位置关系具有相同情况,但是相较于K24-01站,K24-02站的叶绿素a浓度较低,因而推测食物短缺可能是造成该站大磷虾生长状况相对较差的主要原因。G25-09站位于深海区,这与刘会莲等[8]在普里兹湾调查结果一致,即陆坡区大磷虾生长状况要好于深海区。另一方面,海冰消退情况也会影响大磷虾的生长状况。海冰对于大磷虾种群补充具有十分重要的作用[3,25],冬季海冰为大磷虾提供了庇护场,并且以冰藻为主的海冰生物群落可以为大磷虾提供必要的食物,春夏季伴随海冰消退的浮游植物暴发又为大磷虾提供大量的饵料[26-28]。而在采样时,G24-02和K25-02站无海冰覆盖的持续时间较短,推测在海冰覆盖阶段大磷虾因饵料不足发生负生长,由于无冰期持续时间较短,大磷虾还没有从负生长状况中恢复,导致其生长状况较差。
南极夏季海冰的消退状况对大磷虾性期组成和分布也有一定影响。Siegel[29]认为大磷虾幼体、未成体和成体的分布区域在各个季节中都相互分开。冬季,在海冰覆盖区域成体和未成体大磷虾主要位于深水处;春季,在海冰融化区域发生浮游植物暴发,未成体大磷虾主要分布在海冰边缘,而成体大磷虾分布区域更靠近外海[30]。本研究中,G24-03、G24-07及G24-10站磷虾发育期组成中,大磷虾未成体所占比例均超过85%。这三个站位海冰回退时间均较晚,且除了G24-03站外,G24-07和G24-10站采样时均位于冰缘处,表现出未成体大磷虾分布于海冰边缘的特性。在G24-09、G25-06、G25-07和G25-09四个站位,成体大磷虾所占比例均较高,在这些站位采样时,海冰消退已经过去至少1个月时间,站位离冰缘线较远,表现出成体大磷虾远离海冰边缘的分布特征。另外,成体大磷虾中可能也存在一部分个体是经由未成体大磷虾在充分饵料及充分时间内发育而来。
在自然环境下,1岁龄磷虾中未成体大磷虾占绝大部分,而次成体在2岁龄大磷虾中占比例最高,3岁及以上年龄组中,成体大磷虾占比例最高[31]。从9月至次年4月,3岁及以上年龄组雌性大磷虾未成体或未交配雌性成体(3AF)的生殖器官逐渐发育成熟,并经过循环发育卵巢产卵几次后,最后还原回初期卵巢发育水平[32]。普里兹湾海区调查中,南极大磷虾雄性先于雌性成熟[8],主要产卵群体产卵开始于1月,高峰期开始于2月中下旬[12],而较早的产卵对磷虾幼体存活率有十分重要的作用[33-34]。本研究利用IKMT网在K24-01站采集到大磷虾雄性成体,但未采集到雌性成体,推测此区域大磷虾可能正处于生殖初期,而在K24-02站采集到待产雌体3DF,两站均在2008年1月上旬采样,由此推测威德尔海域大磷虾的生殖时间较早。在普里兹湾以东海域(110°E—114°E)K25-01和 K25-02站均采集到较高比例的雄性成体大磷虾(3AM+3BM),并且在K25-02站采集到产后雌体3EF,表明该海域在2009年1月中旬已有磷虾排卵。在普里兹湾K25-04站同样采集到雄性成体及雌性临产及产后雌体大磷虾。在2007/2008年夏季,利用高速采集器在G24-09站采集到临产雌体大磷虾3DF,同时在该站位采集到雄性成体大磷虾3AM和3BM。然而,在2008/2009年夏季,在 G25-06、G25-07和 G25-09站利用高速采集器采集到的雄性磷虾成熟度均较高,同时在G25-06及G25-07站采集到临产雌体大磷虾3DF,表明普里兹湾海域的大磷虾在2009年2月中旬—3月中上旬均有产卵行为。南极大磷虾在不同海域的种群结构存在一定的差异,这对种群补充会产生一定的影响。
本研究对2008年1—3月及2009年1—3月特定海域的南极大磷虾进行了分析,今后还需要对这些海域继续进行长期观测,以获取南极大磷虾生长状况和种群结构的年际变化,并探究其对南极环境变化的指示作用。
4 结语
(1)南极大磷虾在威德尔海域丰度及捕获率都高于普里兹湾海域。2007/2008年南极夏季威德尔海域获取的大磷虾丰度要低于历史同期调查获取的磷虾丰度,而2008/2009年南极夏季普里兹湾海域大磷虾丰度同样低于磷虾历史捕获的丰度。
(2)从两次调查总体上看,南极大磷虾生长状况基本正常,个别生长状况较差站位可能与海冰回退时间较晚或叶绿素a浓度较低有关。
(3)利用高速采集器采集的磷虾样品,在海冰边缘区域站位,大磷虾未成体占有很大比例,而在海冰消退已较长时间的海域站位中,成体大磷虾占比例较大,这反映了未成体与成体不同的分布特征,也可能海冰回退时间较早,给了一部分未成体大磷虾在充分饵料及充分时间条件下发育为成体的机会。
(4)南极大磷虾在不同海域的种群结构存在一定的差异,对种群补充会产生一定的影响。
致谢 感谢雪龙号极地考察船全体船员的帮助,感谢中国科学院海洋研究所张永山和王敏晓同志的采样工作。
1 Marr JW S.The natural history and geography of the Antarctic krill(Euphausia superba Dana).Cambridge:The University Press,1962.
2 Nicol S,Constable A J,Pauly T.Estimates of circumpolar abundance of Antarctic krill based on recent acoustic densitymeasurements.CCAMLR Science,2000,7:87—99.
3 Atkinson A,Siegel V,Pakhomov E,et al.Long-term decline in krill stock and increase in salps within the Southern Ocean.Nature,2004,432(7013):100—103.
4 Siegel V.Distribution and population dynamics of Euphausia superba:summary of recent findings.Polar Biology,2005,29(1):1—22.
5 Loeb V,Siegel V,Holm-Hansen O,et al.Effects of sea-ice extent and krill or salp dominance on the Antarctic food web.Nature,1997,387(6636):897—900.
6 Everson I,Miller GM.Krillmesoscale distribution and abundance:results and implicationsof research during the BIOMASSProgramme//EI-Sayed S Z.Southern Ocean Ecology:The BIOMASSPerspective.UK:Cambridge University Press,1994.
7 Nicol S,Endo Y.Krill fisheries:Development,management and ecosystem implications.Aquatic Living Resources,1999,12(2):105—120.
8 刘会莲,孙松,吉鹏.1999/2000年夏季南极普里兹湾地区南极磷虾的分布、丰度和生长状态研究.极地研究,2001,13(4):231—244.
9 王荣,鲁北伟,李超伦,等.南极磷虾年龄组成的体长频数分布混合分析.海洋与湖沼,1995,26(6):598—605.
10 刘永芹,孙松,张永山,等.2002年1月份南极普里兹湾海域南极磷虾(Euphausia superba Dana)的丰度和种群结构研究.极地研究,2011,23(4):275—282.
11 王荣,陈时华.大磷虾性比、怀卵量与某些生殖特点的分析.南极研究,1989,1(3):61—68.
12 王荣,张云波,仲学锋,等.普里兹湾邻近海域大磷虾的生殖特点研究.南极研究,1993,5(4):12—21.
13 王荣,仲学锋,孙松,等.普里兹湾邻近海域大磷虾的种群结构研究.南极研究,1993,5(4):1—11.
14 Mauchline J.Measurements of body length of Euphausia superba Dana.BIOMASSHandbook Series,1980,4:1—9.
15 Makarov R R,Denys C J.Stages of sexualmaturity of Euphausia superba Dana.BIOMASSHandbook Series,1981,11:1—13.
16 Ikeda T,Dixon P.Body shrinkage as a possible over-winteringmechanism of the Antarctic krill,Euphausia superba Dana.Journal of Experimental Marine Biology and Ecology,1982,62(2):143—151.
17 Sun S,Mare D LW,Nicol S.The compound eye as an indicator of age and shrinkage in Antarctic krill.Antarctic Science,1995,7(4):387—392.
18 Sun S,Wang R.Study on the relationship between the diameter of the compound eye and the growth of the Antarctic krill.Antarctic Research,1996,7(2):87—93.
19 Sun S.Using Antarctic krill as an indicator of environmental interannual change.Korean Journal of Polar Research,1997,8(1-2):97—103.
20 Shin H C,Nicol S.Using the relationship between eye diameter and body length to detect the effects of long-term starvation on Antactic krill Euphausia superba.Marine Ecology Progress Series,2002,239:157—167.
21 Pauly T,Nicol S,Higginbottom I,et al.Distribution and abundance of Antarctic krill(Euphausia superba)off East Antarctica(80—150°E)during the Austral summer of 1995/1996.Deep Sea Research PartⅡ:Topical Studies in Oceanography,2000,47(12-13):2465—2488.
22 Pakhomov E A.Demography and life cycle of Antarctic krill,Euphausia superba,in the Indian sector of the Southern Ocean:long-term comparison between coastal and open-ocean regions.Canadian Journal of Fisheries and Aquatic Sciences,2000,57(S3):68—90.
23 Siegel V,Loeb V,Gröger J.Krill(Euphausia superba)density,proportional and absolute recruitment and biomass in the Elephant Island region(Antarctic Peninsula)during the period 1977 to 1997.Polar Biology,1998,19(6):393—398.
24 刘会莲.中国近海及南大洋磷虾关键种种群生态学研究.青岛:中国科学院研究生院,2002:21—31.
25 Siegel V,Loeb V.Recruitment of Antarctic krill Euphausia superba and possible causes for its variability.Marine Ecology Progress Series,1995,123:45—56.
26 Meyer B.The overwintering of Antarctic krill,Euphausia superba,from an ecophysiological perspective.Polar Biology,2012,35(1):15—37.
27 Daly K L,Macaulay M C.Influence of physical and biologicalmesoscale dynamics on the seasonal distribution and behavior of Euphausia superba in the Antarctic marginal ice zone.Marine Ecology Progress Series,1991,79:37—66.
28 Brierley A S,Thomas D N.Ecology of Southern Ocean pack ice.Advances in Marine Biology,2002,43:171—276.
29 Siegel V.Krill(Euphausiacea)demography and variability in abundance and distribution.Canadian Journal of Fisheries and Aquatic Sciences,2000,57(S3):151—167.
30 Nicol S.Krill,currents,and sea ice:Euphausia superba and its changing environment.BioScience,2006,56(2):111—120.
31 Siegel V.Age and growth of Antarctic Euphausiacea(Crustacea)under natural conditions.Marine Biology,1987,96(4):483—495.
32 Quetin L B,Ross R M.Environmental variability and its impact on the reproductive cycle of Antarctic krill.American Zoologist,2001,41(1):74—89.
33 Ross R M,Quetin L B.Energetic cost to develop to the first feeding stage of Euphausia superba Dana and the effect of delays in food availability.Journal of Experimental Marine Biology and Ecology,1989,133(1-2):103—127.
34 Ross R M,Quetin L B,Newberger T,et al.Growth and behavior of larval krill(Euphausia superba)under the ice in late winter2001 west of the Antarctic Peninsula.Deep Sea Research PartⅡ:Topical Studies in Oceanography,2004,51(17-19):2169—2184.
