Role of miRNA in Mammary Gland Development and Lactation
2014-03-07LiQingzhangWangChunmeiandGaoXuejun
Li Qing-zhang, Wang Chun-mei, and Gao Xue-jun
Key Laboratory of Dairy Science of Education Ministry, Northeast Agricultural University, Harbin 150030, China
Role of miRNA in Mammary Gland Development and Lactation
Li Qing-zhang, Wang Chun-mei, and Gao Xue-jun
Key Laboratory of Dairy Science of Education Ministry, Northeast Agricultural University, Harbin 150030, China
miRNA can regulate development and milk yield of the mammary gland through epigenetic mechanism. miRNA can directly and indirectly modulate the activity of the epigenetic machinery, target genes through post-inhibition of translation initiation, mediate miRNA decay, target genes and inhibit the positive regulation, regulate tone modification, and regulate DNA methylation of target genes. Here we reviewed the role of miRNAs in mammary gland development and lactation. Researching miRNA in mammary gland development and lactation process, and understanding the response of the epigenetic mechanisms to external stimuli will be an important necessity to devise new technologies for maximizing their activity and milk production in the dairy cow.
miRNA, epigenetic, mammary gland, milk production
Introduction
Cells of a multicellular organism are genetically homogeneous but structurally and functionally heterogeneous owing to the differential expression of genes. These alterations have been shown among others involved in the development of the central nervous system, the skin, and the liver (Jaenisch and Bird, 2003). This is epigenetic regulation. Epigenetic is defined as the stable alterations in gene expression potential to arise during development and proliferation (Campos and Reinberg, 2009; Chakrabarty et al., 2007; Bernstein and Hake, 2006; Bestor, 2000). Increasing evidence suggests that, among their roles as posttranscriptional repressors of gene expression, microRNAs (miRNAs) help to confer robustness to biological processes by reinforcing transcriptional programs and attenuating aberrant transcripts, and they may in some network contexts help suppress random fluctuations in transcript copy number (Vasudevan et al., 2007). Although many miRNAs and their target binding sites are deeply conserved, which suggests an important function that a typical miRNA target interaction produces only subtle reduction (<2-fold) in protein level, and many miRNAs can be deleted without creating any obvious phenotype. Early observations of miRNA expression profiles revealed that miRNAs tend to be anti-correlated with target gene expression in contiguous developmental stages or tissues (Fazi et al., 2005).
Correspondingly, a view emerged that miRNA evolved primarily to play the role of a reinforce, in which its activities cohere with transcriptional patterns to sharpen developmental transitions and entrench cellular identities. It is also possible that miRNAs buffer fluctuations in gene expression and more faithfully signal outcomes in the context of certain regulatory networks.
Epigenetic Regulation of Target Gene Transcriptional Level
The non-coding RNAs refer to functional RNA molecules that are not translated into protein, they are divided into non-housekeeping coding RNA (for) and regulating RNA which has the regulation effect of coding RNA. According to their sizes, they were divided into two classes: short chain non-coding RNA (including siRNA, miRNA, and piRNA) and long chain non-coding RNA (including lncRNA RNA). Although in recent, a lot of researches show that the non-coding RNA plays an important role in epigenetic regulation, can regulate gene expression in the genome level and chromosome level, and decides the fate of cell differentiation, compared to other creatures (yeast, fruit flies, worms, plants, and so on), mammalian cell epigenetic research relatively lags behind.
MicroRNA (miRNA) is a class of evolutionarily conservation, approximately 21 to 25 single nucleotide endogenous non-coding single-stranded RNA, encoded by the code in protein genes interval area or the intron miRNA genes encoded of protein for transcription regulation (Avril et al., 2009). miRNA formation process and function mechanism is: first, use miRNA gene transcription to create initial miRNA (pri-miRNA), and then the pri-miRNA is processed into precursor miRNA (pre-alpha-miRNA), then after Drosha and Dicer enzyme identification, it is shore into mature miRNA; mature miRNA shears into RNA induction silence complex (RISC) to form the asymmetric RISC components, and the last step, miRNA degrades mRNA or inhibits the transcription by identifying its corresponding target genes, and translating silence target gene expression and silencing mediated gene (Li et al., 2007; Weber 2005; Rodriguez et al., 2004). Lewis et al. (2005) studied miRNA potential effects and speculated histone methylation enzyme, methylation CpG binding protein, chromatin domain protein and histone acetylation enzyme. Previous studies have shown that miRNA could be caused by regulating histone modification chromatin remodeling, and miRNA could alsoinfluence DNA methylation through regulating DNA methylation enzyme expression (Kedde and Agami, 2008).
Through regulating target gene expression, miRNA stresses the individual development and cell differentiation, proliferation and apoptosis, and other biological process in every level of adjustment. Such as miR-196 in mammalian cells of the target genes for HOXB8 regulates vertebrate development (Mansfield et al., 2004); miR-223 can promote differentiation of neutrophils in hematopoietic stem cell (Fazi et al., 2005). miRNA and its target genes form a complex regulation network, controls the cells and individual life activities (Lewis et al., 2005)
miRNA control of mammary gland development and lactation
Epigenetic modifications can be expected to play a role during mammary gland development. Furthermore, epigenetic also may be defined as "the manifestation of a phenotype, which can be transmitted to the next generation of cells or individual, without alterations to DNA sequence (genotype)" (Watson and Khaled, 2008; Rijnkels et al., 2010). The potential inheritable chemical change of epigenetic regulation (nonsequence) is in chromatin, such as DNA methylation and histone modification, which affect gene expression to manipulate mammary gland on emerging. It is proposed that a substantial proportion of unexplained phenotypic variation in the dairy cow is due to epigenetic regulation. Permanent environmental effects can be further subdivided into within and across lactation (multiple lactation) effects. It is found that roughly 21%-25% of the permanent environmental effect was across an animal's lactation, but how these factors regulate milk production mechanism is not clear (Singh et al., 2010). It must be noted that milk mechanism of epigenetic regulation has been some promising results.
At present, miRNA has been proven in mice, cows, human, including mammals' mammary glandin different development stages, suggesting that it plays an important role in differentially expressing mammary development and lactation. Weber (2005), based on chip technology analysis of mice and human breast tissue, found 23 miRNAs dysregulated in breast; Lee and Dutta (2006) analyzed human breast normal tissue and breast cancer tissue miRNA expression and found that let-7a, let-7b, let-7c, mir-26a, and mir-26 microRNA expressed abnormalities in breast cancer, revealing miRNA participation mammary gland disease. Wang and Li (2007) applied chip technology and found that miRNA was differentially expressed in mice mammary gland at different developmental stages. Applications such as library establishment method in breast fat cells and breast tissue showed that miR-23a, miR-24, and miR-133 might play a role in cattle mammary gland development and lactation (Gu et al., 2007; Avril et al., 2009; Silveri et al., 2006).
miRNA in mammary gland
Through adjusting its target gene expression, miRNA plays a role. A war application, such as gene knock out technology confirmed miR-17-5 p influences breast cancer cell proliferation and transfer process through regulating AIB1 mRNA 3 'UTR sequence (Li et al., 2010; Hossain et al., 2006). Kong reported MiR-155 affected cell proliferation and apoptosis including main control target genes for RhoA in mice mammary gland epithelial cell target TGF and mediated through regulating TGF-beta signaling pathways (Kong et al., 2008; Kong et al., 2010). Tanaka's applications such as chip technology, gene knock out, and carrier construction technology revealed miR-101a regulated beta casein expression, mammary gland epithelial cells, and mammary gland tissue differentiation on mice mammary gland epithelial cells, miR-101a and miR-199a* involved in the expression and regulation of mammary gland differentiation in mammary gland epithelial cells through regulating Cox-2 (cyclooxygenase-2) (Chakrabarty et al., 2007; Cohen et al., 2008; Daikoku et al., 2008; Tanaka et al., 2009).
In recent years, our Key Laboratory of Northeast Agricultural University Dairy Science did some work of miRNA's role in mammary gland development and lactation and obtained some results (Table1).
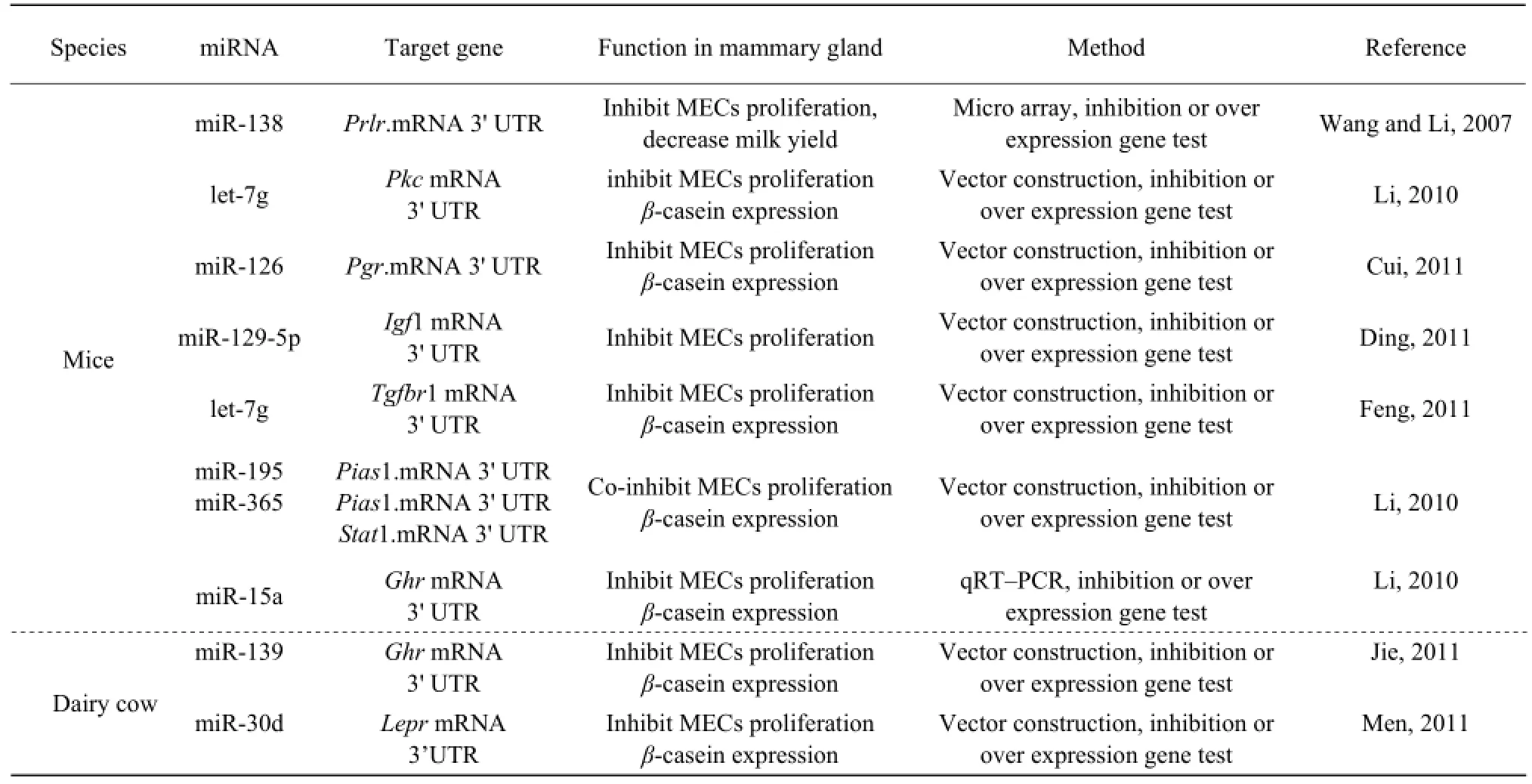
Table 1 Studies on miRNAs in mammary gland by Key Lab of Dairy Science, Ministry of Education
One study showed that miR-126 targeted at progesterone receptor in mammary gland epithelial cells Ding (2011) tested miR-129-5p targeting at IGF-1 in mouse mammary gland epithelial cells through fluorescent element enzyme gene report vector; Feng (2011) applications of bioinformatics technology and construction 3’UTR vector proved that let-7g regulated mice mammary gland development and lactation through targeting at Tgfbr1, and further influenced mammary gland epithelial cells of lactation (Li, 2010). Jie (2011) confirmed that miR-139 targeting at GHR of 3' UTR sequences in cows mammary gland epithelial cells might play a role in cow ductal epithelial lactation, and further regulation mechanism needed to be further discussed. Li's (2010) analysis on miR-15/miR-16 in cow mammary gland at different developmental stages indicated that differential expression of miR-15/miR-16 might control GHR expression, but regulation mechanism needed for further research; Men's (2011) analysis on LEPR genes showed that miR-30d targeting gene was in cow mammary gland. The studies showed that miRNAs involved in mammary gland development and lactation, thus played lactation readjustment function, but the target gene regulation mechanism still needed to be further discussed.
Conclusions
At present, there is little realization about cow mammary gland development and epigenetic lactation. However, many problems need us to further discuss, it is hard to clearly explain the composition of different diets, different environmental factors and different production performance of dairy cows under physiological changes (such as milk yield or milk protein, milk fat) of the mechanism. Furthermore, how the epigenetic mechanisms influence the development and lactation process of the cow mammary gland; how many miRNAs are in work, and what is their action mechanism? How diet composes and environmental factors influence miRNAs change? How they modulate milk yield and milk quality? Although Singh et al. (2010) included that miRNAs in epigenetic regulation might control 25% of the cow milk yield, there are still few researches on these epigenetic factor effect and controls of the quality of milk yield and milk quantity. Ultimately, in the near future, more and more in-depth understanding in how epigenetic regulates cow milk yield and milk quality mechanism will be got.
Avril SLD, Goldstein J, Stingl C, et al. 2009. Characterisation of microRNA expression in post-natal mouse mammary gland development. BMC Genomics, 10: 548.
Bernstein E , Hake S B. 2006. The nucleosome: a little variation goes a long way. Biochem Cell Bio, 84(4): 505-517.
Bestor T H. 2000. The DNA methyltransferases of mammals. Hum Mol Genet, 9(16): 2395-2402.
Campos E I, Reinberg D. 2009. Histones: annotating chromatin. Annu Rev Genet, 43: 559-599.
Chakrabarty A S, Tranguch T, Daikoku K, et al. 2007. MicroRNA regulation of cyclooxygenase-2 during embryo implantation. Proc Natl Acad Sci U S, 104(38): 15144-15149.
Cohen A M, Shmoish L, Levi U, et al. 2008. Alterations in microribonucleic acid expression profiles reveal a novel pathway for estrogen regulation. Endocrinology, 149(4): 1687-1696.
Cui W. 2011. MiR-126-3p targets progesterone receptors and controls development and lactation of mouse mammary gland. Northeast Agricultural University, Harbin.
Daikoku T Y, Hirota S, Tranguch A R, et al. 2008. Conditional loss of uterine Pten unfailingly and rapidly induces endometrial cancer in mice. Cancer Res, 68(14): 5619-5627.
Ding W. 2011. miR-129-5p and its target gene Igf-1in mouse mammary gland. Northeast Agricultural University, Harbin.
Fazi F A, Rosa A, Fatica V, et al. 2005. A minicircuitry comprised of microRNA-223 and transcription factors NFI-A and C/EBPalpha regulates human granulopoiesis. Cell, 123(5): 819-831.
Feng L. 2011. let-7g and its target gene in mouse mammary gland development and lactation. Northeast Agricultural University, Harbin.
Gu Z L, Eleswarapu S, Jiang H L. 2007. Identification and characterization of microRNAs from the bovine adipose tissue and mammary gland. FEBS Lett, 581(5): 981-988.
Gonzalez S, Pisano D G, Serrano M. 2008. Mechanistis principles ofchromatin remodelling guided by siRNAs and miRNAs. Cell Cyc, 7(16): 2601-2608.
Hossain A M, Kuo T, Saunders G F. 2006. Mir-17-5p regulates breast cancer cell proliferation by inhibiting translation of AIB1 mRNA. Mol Cell Bio, 26(21): 8191-8201.
Jaenisch R, Bird A. 2003. Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals. Nat Genet, 33: 245-254.
Jie J. 2011. miR-139 and its target gene in cow mammary gland. Northeast Agricultural University, Harbin.
Kedde M, Agami R. 2008. Interplay between microRNAs and RNA-binding proteins determines developmental processes. Cell Cycle, 7(7): 899-903.
Kong W, He L, Coppola M J, et al. 2010. MicroRNA-155 regulates cell survival, growth, and chemosensitivity by targeting FOXO3a in breast cancer. J Biol Chem, 285(23): 17869-17879.
Kong W, Yang H, He L, et al. 2008. MicroRNA-155 is regulated by the transforming growth factor beta/Smad pathway and contributes to epithelial cell plasticity by targeting RhoA. Mol Cell Biol, 28(22): 6773-6784.
Lee Y S, Dutta A. 2006. MicroRNAs: small but potent oncogenes or tumor suppressors. Curr Opin Investig Drugs, 7(6): 560-564.
Lewis B P, Burge C B, Bartel D P. 2005. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell, 120(1): 15-20.
Li H M. 2010. Role of miR-15a in mammary gland and mammary epithelial cells of dairy cow. Northeast Agricultural University, Harbin.
Li S C, Tang P, Lin W C. 2007. Intronic microRNA: discovery and biological implications. DNA Cell Biol, 26(4): 195-207.
Li Y, Tian L, Wang C M, et al. 2010. Expression of let-7g in development, lactation and involution of the murine mammary gland. J Dairy Sci, 93(1): 483.
Mansfield J H, Harfe B D, Nissen R J, et al. 2004. MicroRNA-responsive 'sensor' transgenes uncover Hox-like and other developmentally regulated patterns of vertebrate microRNA expression. Nat Genet, 36(10): 1079-1083.
Mattick J S, Amara P P, Dinger M E, et al. 2009. RNA regulation of epigenetic processes. Bioessays, 31(1): 51-59.
Men J. 2011. miR-30d and its target gene in cow mammary gland. Northeast Agricultural University, Harbin.
Rijnkels M, Kabotyanski E, Montazer-Torbati M B, et al. 2010. The epigenetic landscape of mammary gland development and functi onal differentiation. J Mammary Gland Biol Neoplasia, 15(1): 85-100.
Rodriguez A S, Griffiths-Jones J L, Ashurst, et al. 2004. Identification of mammalian microRNA host genes and transcription units. Genome Res, 14(10A): 1902-1910.
Silveri L G, Tilly J L, Vilotte, et al. 2006. MicroRNA involvement in mammary gland development and breast cancer. Reprod Nutr Dev, 46(5): 549-556.
Singh K, Erdman R A, Swanson K M, et al. 2010. Epigenetic regulation of milk production in dairy cows. J Mammary Gland Biol Neoplasia, 15(1): 101-112.
Tanaka TS, Haneda K, Imakawa S, et al. 2009. A microRNA, miR-101a, controls mammary gland development by regulating cyclooxygenase-2 expression. Differentiation, 77(2): 181-187.
Vasudevan S, Tong Y, Steitz J A. 2007. Switching from repression to activation: microRNAs can up-regulate translation. Science, 318(5858): 1931-1934.
Wang C, Li Q. 2007. Identification of differentially expressed micro-RNAs during the development of Chinese murine mammary gland. J Genet Genomics, 34(11): 966-973.
Watson C J, Khaled W T. 2008. Mammary development in the embryo and adult: a journey of morphogenesis and commitment. Development, 135: 995-1003.
Weber M J. 2005. New human and mouse microRNA genes found by homology search. Febs J, 272(1): 59-73.
S512.1
A
1006-8104(2014)-01-0070-05
Received 4 December 2012
Support by the Natural Science Foundation of China (31072103)
Li Qing-zhang (1953-), male, professor, supervisor of Ph. D student, engaged in the research of lactation biology. E-mail: qzl@neau.edu
杂志排行

Journal of Northeast Agricultural University(English Edition)的其它文章
- Study on Relationship Between Cucumber Germplasm and Propamocarb Residue Using Subjective Rating Technique
- Cloning and Expression Analysis of Mlo Gene from Pericallis hybrida B. Nord.
- Simulation of in situ Root Decomposition of Two Barley Cultivars
- Vertical Migrating and Cluster Analysis of Soil Mesofauna at Dongying Halophytes Garden in Yellow River Delta
- Effect of MSTN Propeptide and shRNA Co-expression Vector on Proliferation of Skeletal Muscle Satellite Cells
- Application of Protein Feed Processed by Microbial Fermentation to Dairy Cow