青海云杉无性系种子园雌雄球花空间分布与管理
2014-03-06何海金胡勐鸿牟小梅
何海金 孙 斌 胡勐鸿 牟小梅
(甘肃省小陇山林业实验局,陇南,742303)
青海云杉(Picea crasifolia Mill)是我国青藏高原东北边缘特有的森林建群树种,主要分布在青海、甘肃、宁夏、内蒙古及四川西部[1];水平分布在32°40' ~41°30'N,98°40' ~112°30'E,海拔1 750 ~3 550 m,但多集中在2 200 ~3 200 m;甘肃祁连山为适中分布区。青海云杉寿命较长、树干通直、树姿雄伟、材质优良,不仅是重要的建筑和纸浆材树种,更是祁连山、贺兰山和大青山等水源涵养林的主要组成树种,是具有很大优势而又相当稳定的基本森林类型,其水源涵养林是该区人民赖以生存和发展的生态保证[2-6];又由于青海云杉耐旱、抗寒、适应性强,是西部生态脆弱区生态恢复建设、城市绿化的主要树种之一。随着天然林保护工程和退耕还林还草工程的实施,对青海云杉良种需求量进一步扩大,青海云杉遗传改良起步较晚,种子园管理和研究工作处在起始阶段[7]。种子园内各组成亲本开花结实的不平衡性直接影响种子园的产量和品质[8],良种难以满足造林用种的需求。为种子园管理,国外以天然和人工林分为对象,进行过雌雄球花在树冠中的分布模式以及与雌雄球花产量关系的研究[9-11]。国内对油松(Pinus tabulaeformis carr)、余甘子(Phyuanthus emblica L.)、杉木(Cuuninghamia lanceolata)、思茅松(Pinus kesiya var.langbianensis)等树种进行过雌雄球花空间分布的研究[12-15]。赖焕林等[16-17]对马尾松(Pinus massoniana Lamb)嫁接和实生植株分枝特性与种子园产量进行过研究。云杉母树雌雄球花数量的多少反映了结实潜力的大小,雌雄球花在树冠中的分布受到本身的遗传特性、树冠结构、发枝生物学特性以及光结构等因子的影响,马建伟[18]、雷军等[7]分别对粗枝云杉(Picea asperata Mast)、青海云杉开花生物学特性、雌雄球花的分布、数量有过报道,但缺乏种子园无性系水平的分析。根据青海云杉种子园树体管理的需要,在沙坝落叶松/云杉国家良种基地,以青海云杉嫁接无性系种子园开花结实的无性系分株为研究对象,研究了青海云杉树冠结构、分支特性、雌雄球花的空间分布特征以及无性系间、内树冠不同方位雌雄球花数量。对准确预测青海云杉种子产量,进行整形修剪,促进雌雄球花的合理分布,达到稳产、丰产具有重要意义。
1 研究区自然概况
甘肃小陇山沙坝国家云杉/落叶松良种基地,位于天水市秦州区娘娘坝镇,地理坐标为105°51'27″~105°54'51″N,34°07'28″~34°10'22″E,海拔1 560 ~2 019 m。属大陆性季风气候,为温带湿润区,冬无严寒夏无酷暑,春季气温回升快,常有倒春寒和春旱发生。年均气温7.2 ℃,极端最高气温32 ℃,极端最低气温-27 ℃,≥10 ℃的有效积温2 480 ℃,无霜期154 ~185 d。年均降水量460 ~800 mm,降水集中分布在7、8、9月份,一般夏秋雨占年降水量的70% ~80%,年蒸发量989 ~1 658 mm,平均相对湿度78%,平均年日照时间1 553 h。土壤为棕色森林土,土壤有机质质量分数为1.71% ~5.35%,pH值为5.72 ~7.90,土层厚度30 ~50 cm,土壤肥力较高[19]。
2 材料与方法
研究材料为青海云杉嫁接无性系种子园中正常开花结实的23年生各无性系分株。株行距6 m ×6 m,无性系顺序错位排列,平均树高9.6 m,平均胸径17.6 cm,郁闭度0.6,活枝下高1.45 m。
在青海云杉种子园随机选择12 个无性系,每个无性系选3 个分株。参照李国峰等[12]对油松雌雄球花空间分布特征的研究。花期(4月23日—5月2日)按无性系分株调查雌雄球花量,分别调查无性系分株轮生枝层数、每个轮生枝层的轮生枝数量(个),每个轮生枝层选择树冠S(南向)、N(北向)的1 个轮生枝,调查轮生枝枝序(从主干轴向外生长发育的顺序)、分枝角(轮生枝与主干的夹角)、每个枝序上雌雄球花数量、着生雌雄球花的枝龄(2 级以上侧枝生长发育的枝龄)。
计算12 个无性系分株的雌雄球花比(H(雌)∶ H(雄))、选择典型无性系绘制雌雄球花在树冠N 向垂直剖面上的数量分布图,根据典型无性系不同冠层、不同枝序上的雌雄球花量,绘制不同冠层和枝序上雌雄球花数量变化曲线图;采用SPSS16.0 对无性系间雌雄球花数量和无性系内树冠S、N 雌雄球花数量进行方差分析(GLM)、多重比较(LSD)。
3 结果与分析
3.1 青海云杉树冠特征和分枝特性
青海云杉树冠呈圆锥状(或伞形),自下而上由轮生枝构成树冠主体,1年一层轮生枝,轮生枝层明显。轮生枝在树冠各方位的分布比较均匀,一般是4 ~7 个,但是个别无性系轮生枝较多,可达8 ~10个。轮生枝与树干的夹角从基部向上逐渐缩小,越往顶部分枝角越小。但有的无性系自下而上分枝角没有变化,有的无性系分株由于下部轮生枝下垂分枝角≥90°,轮生枝先端上翘。青海云杉的分枝特性为:一级侧枝(轮生枝)上可分生二级侧枝,二级侧枝上分生三级侧枝,依次类推,可分生2 ~5 级,枝条数一般为3 ~5 条;但是4 ~5 级侧枝一般为2 个枝条。在下年度各级侧枝上的侧芽又会萌发,生长新的侧枝,侧芽萌发的枝条不对称,分散在各级侧枝上,侧芽萌发枝条的数量与侧枝的长度和健康状况有关,侧枝越长,侧枝上的芽数就多,萌发的侧枝数越多;但是侧枝上有些侧芽不萌发枝条,枝条上有隐芽,当枝条受到刺激后,如机械损伤、风折等时,刺激隐芽萌发,生长出枝条。隐芽萌发的枝条比较细弱。随着树龄的增大,枝序增加,树冠内堂由于光照不足,多数侧枝干枯。只有1 ~12 a 的枝条能够正常发育。
3.2 青海云杉雌雄球花在枝条上的着生方式
青海云杉雌雄球花芽在上年度已经完成分化。雄球花着生在1 ~9 a 枝龄先端1年生冬梢上,着生方式有顶生、侧生、基部簇生3 种着生方式;雄球花着生在比较细弱的枝条上。雌球花着生在1年生枝条顶端,通常为1 枚、单生,或1 ~3 枚。
3.3 雌雄球花在树冠上的垂直分布
青海云杉雄球花比雌球花多,通常球花数H(雌)∶ H(雄)=1∶ 5.6;但是有偏雄无性系,雄球花远比雌球花多,球花H(雌)∶ H(雄)=1∶ (24.53 ~39.64),无性系间球花H(雌)∶ H(雄)比的变异系数为59.38%。由图1可以看出,雌雄球花在树冠中的分布特征是雄球花分布在树冠中下部,有花冠层占68.75%,无性系间变幅62.5% ~75.0%,变异系数为9.09%。雄球花在树冠中的垂直分布有2种情况(图2),其一是雄球花在树冠最低的几个轮生枝层数量多,越往上的几个冠层雄球花较少,直到没有,如73 号无性系;另一种是由于受到光照等因子影响最低的1 ~2 个冠层没有雄球花分布(如136号无性系),或者很少(如191 号无性系),但在中间冠层雄球花数量分布较多,集中分布在3 ~9 轮层间,越往上面的冠层雄球花分布数量越少,直到没有,多数无性系雄球花的分布属于这种类型。轮层间雄球花数量变化与光照、枝条数量有关。调查发现枝条稀疏、内堂通风透光性好的无性系,最低的几个冠层雄球花分布数量多。但随着树龄的增加,最上面冠层的轮生枝虽然光照充足,但是枝条数量少,多为营养枝,所以雄球花少。

图1 191 号无性系树冠N 向垂直剖面上雌雄球花的数量分布
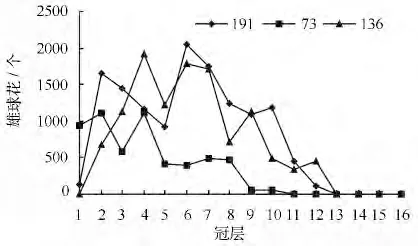
图2 青海云杉雄球花在树冠上的垂直分布
雌球花在树冠中的垂直分布与雄球花相反,分布在树冠中上部顶层轮生枝以下,有花冠层占总冠层的66.67%,有些无性系雌球花的分布占到总冠层的93.0% ,无性系间有花冠层的变异系数为35.49%;比雄球花有花冠层的变异系数大。雌球花在树冠中的垂直分布有3 种(图3),第一,雌球花集中分布在树冠中上部,在接近地面的轮生枝上没有雌球花分布,绝大多数无性系属于这种分布形式;第二,顶层轮生枝以下均有雌球花分布,各轮层数量变化不大,但是这种分布的无性系较少;第三,在接近地面的轮层有少数雌球花,但是多数雌球花集中在树冠中上部。
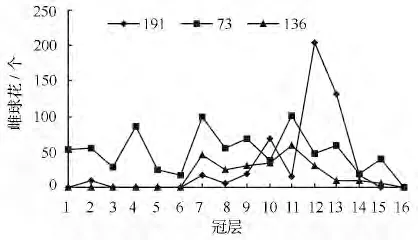
图3 青海云杉雌球花在树冠上的垂直分布
云杉雌雄球花的这种分布特征是长期自然选择的结果,受到无性系本身强的遗传控制,在自然条件下能减少自交几率、有利于种子的飞散和传播。云杉雌球花的这种空间分布格局,表明云杉雌球花生长发育过程中对光照和养分条件要求比雄球花高。但不利于种子园采种,增加了采种难度。因此,在种子园整形修剪时进行截顶,降低结实层,减小采种难度。
3.4 雌雄球花在树冠上的水平分布
青海云杉雄球花大多无性系水平分布集中在2 ~8 枝序间(图1、图4),随着枝序的增加,靠近主干(内堂枝条干枯)很少或者没有雄球花分布。但不同无性系间有差别,有些无性系分株在靠近主干(内堂)有少数雄球花分布,如191 号无性系,雄球花在1 ~14 枝序间有分布;73 号无性系靠近地面的几个轮生枝2 ~13 枝序间有雄球花分布,但集中分布在3 ~8 枝序间,在5 枝序间分布最多;136 号无性系雄球花在2 ~6 枝序间分布最多。雄球花在树冠中的水平分布从外向内,随着枝序的增加雄球花分布数量减少,雄球花集中分布在3 ~8 枝序间,由于光照不足靠近主干没有雄球花分布。青海云杉无性系雄球花的分布多数属于这种类型。但是枝条稀疏,内堂通风透光较好的无性系分株,内堂也有雄球花分布。多数无性系靠近主干由于光照不足,枝条干枯,丧失生殖能力。所以,在云杉种子园树体整形修剪时进行疏枝、短截,可增加树冠内通风透光,刺激隐芽萌发枝条,增加树冠内雄球花着生的枝条数。
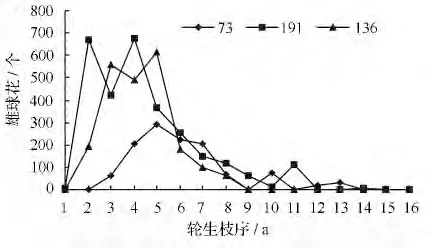
图4 雄球花在树冠上的水平分布
雌球花在树冠上的水平分布不同于雄球花,雌球花在树冠上的水平分布比较一致,集中分布在1 ~4 枝序间;但是个别无性系在1 ~7 枝序间、甚至10 枝序上也有雌球花分布(如73 号无性系)(图5)。
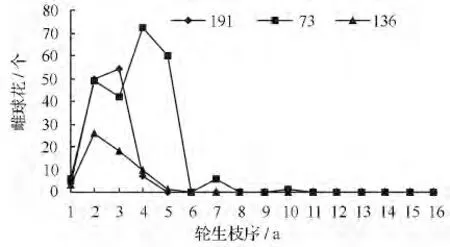
图5 雌球花在树冠上的水平分布
3.5 无性系间雌雄球花数量分析
经方差分析,12 个无性系间雌雄球花数量差异达显著水平,无性系内分株间差异不显著(表1)。由表1可看出,青海云杉不同的无性系由于遗传基础的差异,雌雄球花数量不同,同一无性系分株间由于遗传基础一致,雌雄球花数量差异不显著。雌雄球花数量主要受不同无性系强的遗传控制。
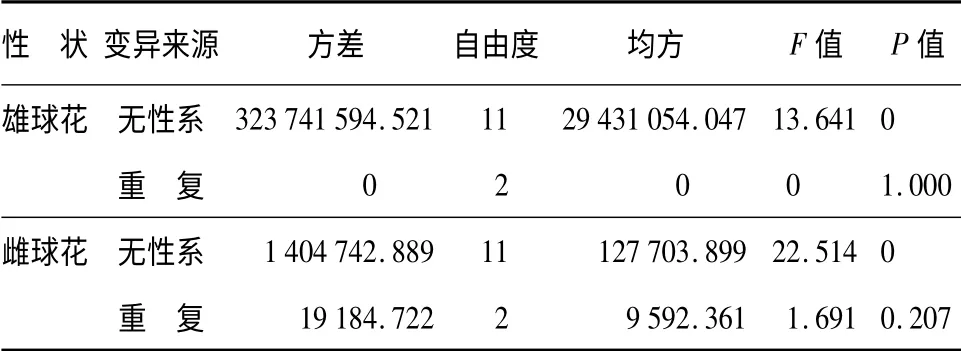
表1 青海云杉无性系雌雄球花量方差分析
无性系间雄球花量的总体平均值为10 127.375枚,总体变异系数为32.16%,变异系数的变幅为0.44% ~44.51%,雄球花量除32、97 号无性系变异系数较大外,其余无性系雄球花量的变异系数较小;雌球花量的总体平均值为537.778 枚,总体变异系数为39.12%,变异系数的变幅为0.79% ~47.58%,130号无性系雌球花量的变异系数最大,其余无性系雌球花量的变异系数均小于总体变异系数。说明无性系分株由于受相同的开花基因控制,所以,雌雄球花量的变异系数小,个别无性系分株间变异系数大,主要是受到树冠结构、光照等环境因子影响。无性系间雌雄球花量多重比较结果(表2):除雄球花量73、76、124、32 号无性系间,97、32 号无性系间,130、136、58、183、191、156、16 号无性系间差异不显著;雌球花量73、32、76、124 号无性系间,76、124、97、130、136、58 号无性系间,183、191、156、16 号无性系间差异不显著外,其余无性系间雌雄球花量差异显著。

表2 青海云杉无性系雌雄球花多重比较
3.6 无性系内树冠不同方位雌雄球花数量分析
对无性系内(以191 号无性系为例)树冠S、N雌雄球花量进行方差分析,结果表明,雌球花(F =1.070,P=0.341)和雄球花(F =0.010,P =0.922)差异不显著。表明同一无性系不同分株由于遗传基础一致,雌雄球花在树冠不同方位的分布比较均匀。
4 结论与讨论
青海云杉雄球花分布在树冠中下部,有花冠层占68.75% ,无性系间有花冠层的变异系数为9.09%;雌球花的分布与雄球花相反,雌球花主要分布在树冠中上部顶层轮生枝以下,有花冠层占66.67%,有的无性系有花冠层占93.0%左右,有花冠层的变异系数为35.49%。这与雷军等[7]对青海云杉、马建伟等[18]对粗枝云杉雌雄球花空间分布的结论一致。云杉属树种雌雄球花在空间的这种分布格局是受云杉属树种的遗传特性、树冠结构、发枝生物学特性控制。经观察,欧洲云杉(Picea abies)、白云杉(Picea gluca)、川西云杉(Picea balfouriana)等云杉属树种,雌雄球花在树冠上的分布与青海云杉相似;雌球花集中分布在树冠顶端2/3 以上树冠外围1年生枝条上,雄球花与雌球花相反,垂直分布主要在树冠中下部,水平分布为树冠外围比树冠内多,靠近主干很少或没有雄球花分布。随着树龄的增加树冠内堂枝条干枯,结实层不断上移向外延伸,增加了采种的困难和危险。对青海云杉、粗枝云杉疏枝和枝条回缩后,树冠内堂光照增加,刺激隐芽萌发枝条,就增加了雌雄球花着生的枝条数。因此,在云杉种子园树体整形修剪时根据雌雄球花的分布规律和分枝特性,采取截顶、疏枝、枝条回缩等修剪措施,增加通风透光,促进树冠内堂隐芽萌发枝条,降低结实层,减小采种难度和危险。
云杉雌雄球花的这种空间分布格局是长期自然选择的结果。有利于减少云杉种内自交、种子的飞散和传播。雷军等[7]的研究认为,青海云杉无性系异型交配的几率大;由于云杉为风媒传粉,花粉有双气囊,易于飞翔,如果遇到高温大风天气,花粉上升增加了自交的几率,自交容易产生空子[21-22],这也许是云杉空子率多的原因之一,种子园在可授期进行人工辅助授粉,减少自交,提高种子产量和品质。
国内对马尾松(Pinus massoniana Lamb)、湿地松(Pinus elliottii Englem)、思茅松等无性系种子园的研究表明,无性系间结实能力由固有的遗传品质决定,同时受气候、园址、立地、密度、坡向、建园材料配置、经营管理、病虫害等各种因子的综合影响[22-26,15]。青海云杉雌雄球花数量的大小反映了结实潜力的大小。青海云杉雄球花比雌球花多,一般球花H(雌)∶ H(雄)=1∶ 5.6,这与粗枝云杉球花H(雌)∶ H(雄)的比接近,粗枝云杉球花H(雌)∶H(雄)=1∶ (4.5 ~5.0)[18],但存在偏雄无性系,H(雌)∶ H(雄)=1∶ (24.53 ~39.64),变异系数为59.38%。青海云杉无性系间雌雄球花数量差异显著[7],但是同一无性系分株间,同一无性系树冠不同方位雌雄球花分布比较均衡。在云杉种子园升级换代时尽可能选择雌雄球化分布比较均衡、有花冠层较高的无性系,合理配置建园材料。不仅提高种子产量和品质,也有利于采种。雌雄球花数量的变化将对种子园的产种量和遗传品质产生深刻的影响[27]。今后在云杉属树种遗传改良和种子园管理中,应进行雌雄球花分化时间、分化机理的研究[28-29],从分子水平进行开花控制基因、人工诱导处理,如ABA、NAA、GA1/3、GA4/7 等生物调控技术的研究,根据需要合理调整云杉雌雄球花在树冠上的分布和比例;又结实需要消耗大量营养物质[30],还需要进行施肥时间、施肥量、施肥种类对云杉种子产量和遗传品质影响的研究。
[1] 刘兴聪.青海云杉[M].兰州:兰州大学出版社,1992.
[2] 王娅丽,李毅,陈晓阳.祁连山青海云杉天然群体表型性状遗传多样性分析[J].林业科学,2008,44(2):70-77.
[3] 李萍,李毅.青海云杉遗传结构研究[J].甘肃农业大学学报,2005,40(3):363-367.
[4] 张立杰,赵文智,何志斌.青海云杉(Picea crassifolia)种群格局的分形特征及其影响因素[J].生态学报,2008,28(4):1383-1389.
[5] 张立杰,蒋志荣.青海云杉种群分布格局沿海拔梯度分形特征的变化[J].西北林学院学报,2006,21(2):64-66.
[6] 赵传燕,别强,彭焕华.祁连山北坡青海云杉林生境特征分析[J].地理学报,2010,65(1):113-121.
[7] 雷军,张宏斌,范菊萍.青海云杉无性系种子园开花习性[J].西北林学院学报,2012,27(6):70-74.
[8] 孔东升,马力,邓延安,等.青海云杉半同胞子代测定初探[J].西北林学院学报,2007,22(1):59-62.
[9] Jensen E C,Long J N.Crown structure of a codominant douglasfir[J].Can J For Res,1983,13(2):246-269.
[10] Caron G E,Powell G R.Patterns of cone distribution in crowns of young Picea mariana.I.effect of tree age on seed cones[J].Can J For Res,1992,22(1):46-55.
[11] Kathleen J T,Powell G R.Production and distribution of seed and pollen cones on Larix larcina trees in young plantations[J].Can J For Res,1991,21(4):446-454.
[12] 李国锋,李军,雷跃平,等.油松雌雄球花的空间分布特征[J].河南农业大学学报,1997,31(1):59-65.
[13] 王开良,姚小华,任华东,等.余甘子雌雄花空间分布规律研究[J].江西农业大学学报,2006,28(2):244-248.
[14] 陈晓阳,秦向华,潘奇敏,等.种子园优质高产技术[M].北京:中国林业出版社,1994:46-154.
[15] 段安安,许玉兰,王秀荣.思茅松无性系种子园雌雄球花分布特性及人工辅助授粉的研究[J].云南林业科技,2002,101(4):22-26.
[16] 赖焕林,邱进清,陈敬德,等.马尾松种子园嫁接植株的分枝与结实特性探讨[J].南京林业大学学报,1998,22(3):86-90.
[17] Lai Huan-lin,Wang Zhang-rong,Jiang Rui-rong.Branching and growth of plantings in fifth year of a seedling seed orchard of Masson pine(Pinus massonina Lamb.)[J].Journal of Forestry Research,2002,13(1):28-32.
[18] 马建伟,胡勐鸿,靳景春.云杉开花生物学和杂交技术[J].种子,2010,29(11):1-5.
[19] 张俊华,刘金鹏,巨天珍,等.天水小陇山森林土壤肥力特征分析[J].干旱区资源与环境,2004,18(8):103-106.
[20] 王晓茹,沈熙环.对由胚珠败育和空粒引起油松种子园减产的分析[J].北京林业大学学报,1989,11(3):60-65.
[21] 贾桂霞,杨俊明,沈熙环.落叶松种间交配结实力变异和自交衰退的研究[J].林业科学,2003,39(1):62-68.
[22] 秦国峰,汪名昌.马尾松开花结实规律的初步研究[J].林业科学研究,1991,4(3):328-332.
[23] 陈敬德.马尾松无性系种子园产量变异的研究[J].南京林业大学学报,1998,22(3):81-85.
[24] 王以珊,罗敏,曾令海,等.气候条件对马尾松种子园产量的影响[J].广东林业科技,2002,18(3):10-15.
[25] 洪永辉,陈惠敏,陈亚斌,等.影响马尾松种子园产量的种实害虫及综合防治[J].福建林业科技,2006,33(4):124-127.
[26] 吴际友,龙应忠.湿地松种子园无性系种实性状遗传与变异研究[J].林业科技通讯,1995(11):10-12.
[27] 陈友力.马尾松无性系种子园球花量变化研究[J].福建林业科技,2001,28(2):50-53.
[28] 江玲,王章荣.马尾松雌雄球花的形成与赤霉素和脱落酸及细胞分裂素的关系[J].南京林业大学学报,1998,22(3):61-65.
[29] 王玉荣,李爱民,吴鸿.马尾松雌球果的发生和早期发育研究[J].西北植物学报,2003,23(7):1182-1187.
[30] Pukkala T.Effect of seed production on the annual growth of Picea abies and Pinus sylvestris[J].Silva Fennica,1987,21(2):145-158.
