引种欧洲云杉自由授粉家系种实性状的多样性1)
2014-03-06马建伟胡勐鸿张宋智雷曙光王军辉
马建伟 胡勐鸿 张宋智 雷曙光 王军辉
(甘肃省小陇山林业实验局林业科学研究所,天水,741022) (国家林业局林木培育重点实验室(中国林业科学研究院林业研究所))
欧洲云杉(Picea abies (L.)Karst)又名挪威云杉。天然分布主要在中欧和北欧,大致分布在北纬42° ~69.5°,是典型的山地树种[1-2]。由于材质硬,纤维长,纹理细密,质地均匀,用途广。其既是西欧、北欧波罗的海沿岸国家,俄罗斯和加拿大的重要工业用材树种,也是重要的建筑用材树种和产脂树;还因树形优美、树冠常绿,常用作圣诞树和园林绿化[3];其针叶和嫩枝不仅可做云酒,药用价值也备受关注。世界各地广为引种,其中日本北海道、美国东北部和加拿大东部引种的欧洲云杉优于乡土树种[4]。我国庐山、长春、辽宁熊岳、呼和浩特、北京、延安、青岛、杭州、湖北建始、沙坝落叶松/云杉国家良种基地均有引种栽培[1],表现出较强的适应性,而且比国内云杉种生长快[2-3]。
球果和种子是针叶树种有性繁殖系统的重要组成部分,是生存和繁衍适应性的集中体现,具有受遗传控制较强的特征,同一物种内球果和种子性状被认为是相对稳定的,最具有区分和比较意义的[5];但是它们长期在自然界不同的选择压力下表现出较大的适应性[6]。具有丰富的遗传变异,球果大小反映了雌性生殖器官的营养状况,是母体营养状况和用于有性生殖投入的体现[7];球果大小与种子大小和质量密切相关[8-9],种子性状的大小表征了种子内营养物质的多少[10],种子性状变异的大小影响到种子的传播和萌发[11],对幼苗的定居和存活有很大的影响[12-13],与子代后期生长及适应性显著相关[14],从而影响到种群更新[15]。国外对欧洲云杉[16-19]、白云杉[20-23]、东方云杉、黑云杉[24-25]、西加云杉[26]等地理种源、群体间、群体内的种实性状、授粉效率、种子园管理与产种质量等进行过研究;国内对青海云杉[27]、天山云杉[28]、川西云杉[29]、粗枝云杉[30]等地理种源群体间和群体内种实性状遗传变异进行过研究,一致的结论认为云杉群体间、群体内(即家系间)种实性状存在着丰富的遗传变异,群体内的遗传变异大于群体间。物种遗传有固有的形态特征,许多针叶树种研究者通过分析针叶、球果和种子的表型性状来探讨性状变异、分化及其与环境因子的关系[31]。球果和种子是容易测得的表型性状,表型性状是基因型和环境共同作用的结果[17],表型性状的遗传多样性反映了家系(群体)的遗传多样性,通过简单、容易测得的表型性状研究家系(群体)的遗传多样性因简便、快速和节省费用,至今仍然是重要而其有效的方法[32]。由于缺乏材料,关于家系种实性状研究从未见报道。为认识引入材料的遗传复杂性、变异大小,文中对沙坝落叶松/云杉国家良种基地44 个欧洲云杉家系(单株)种实性状表型多样性进行了研究,评价了欧洲云杉家系之间的种实性状的遗传变异,期望为欧洲云杉种子园营建、多目标杂交育种和遗传改良提供依据。
1 研究地自然概况
小陇山林业实验局沙坝国家落叶松/云杉良种基地位于甘肃小陇山林区腹地,隶属甘肃省天水市秦州区娘娘坝镇。地理坐标为105°51'27″~105°54'51″N,34°07'28″~34°10'22″E,海拔1 560 ~2 019 m,属大陆性季风气候,为温带湿润区,气候特点是冬无严寒夏无酷暑,春季气温回升快,常有倒春寒和春旱发生。年均气温7.2 ℃,极端最高气温32 ℃,极端最低气温-27 ℃,年均降水量1 012.2 mm,平均相对湿度78%;平均年日照时间1 553 h;≥10 ℃的有效积温2 480 ℃,无霜期154 ~185 d。引种林地处167 林班山麓,为沟谷河滩地,地势平坦,坡度2°左右,土壤为棕色森林土,母质为河流冲积物,土层厚度30 ~50 cm,多含砂、砾和石块。地被物(苔藓、地衣)盖度70%,灌木盖度30%,林龄26 a,林分郁闭度0.9。
2 材料与方法
研究材料为林科院从国外引进的种子,1983年在沙坝育苗,1985年造林。采自沙坝落叶松/云杉国家良种基地26年生欧洲云杉引种林中2009年结实的44 个家系的自由授粉成熟球果。
球果长、宽,鲜、干质量测定:每个家系随机选取20 个(不足20 个的全部选取)生长正常的球果,采用钢卷尺测量球果长CL,精度0.1 cm,采用电子游标卡尺测量球果宽(最宽处)CW,精度0.01 mm,计算球果长/宽。采用电子天平称量每个球果的鲜质量CF,W,球果风干后,每个家系随机称取10 个(不足10 个的全部称量)球果的风干质量CD,W,精度0.01 g。
球果可育种鳞的测定:种鳞分为可育种鳞和不育种鳞[33]。云杉位于球果基部和顶部1 ~2 圈为不育种鳞,没有结种能力;可育种鳞位于球果的中部,含正常发育胚珠,体现了球果结种的潜在能力。每个家系随机测量10 个发育正常球果的可育种鳞数FS;可育种鳞密度FS,D是球果可育种鳞数与球果长度的比(个·cm-1),体现了球果长度与球果可育性的关系。
球果种子潜能和种子效率:种子潜能SP是球果在生物学理论上能够产生种子的最大数目;种子效率SE则反映了球果实际生产种子的情况,其受遗传与环境因子影响。欧洲云杉球果每一个可育种鳞能够生产的种子数为2 粒,按照以下公式计算种子潜能和种子效率。
种子潜能=2 ×该球果的可育种鳞数,
种子效率=(球果生产的健全种子数/SP)×100%。
球果产种数与产种质量:每个家系随机选取10个(不足10 个的全部测量)生长正常的球果,测量每个球果生产的种子总数CS,N,并用水选法测量球果生产种子的饱子数,它反映了受精成功发育成种的胚珠数量。计算饱子率=(饱子数/种子总数)×100%。用电子天平称取单个球果生产的饱子质量CS,W。
种鳞性状的测定:每个家系随机选取1 个生长正常的球果,从基部1/3 以上(球果中部)随机测量10 个可育种鳞的种鳞长SS,L、种鳞宽SS,W,计算种鳞长/宽。
种子性状的测定:种子脱离后每个家系随机选取10 粒种子,用电子游标卡尺测量种子长SL、宽SW,种翅长SW,L、宽SW,W,精度0.01 mm。计算种子长/宽、种翅长/宽。
千粒质量的测定:采用电子天平测量每个家系的千粒质量,3 次重复,精度0.01 g。
种子发芽势和发芽率的测定:净种后每个家系随机选择100 粒种子,温水(30°左右)浸泡24 h,待充分吸水后,每20 粒为1 次重复,5 次重复。将每个重复的供试种子间隔整齐摆放在有定性滤纸的培养皿内,培养皿放置恒温发芽箱内,发芽箱温度控制在最适宜温度25 ℃左右[34]。观察发芽过程,以胚根长度达到种子等长、胚芽长度达到种子一半作为种子萌发标准。种子发芽后每天记录发芽种子粒数,计算种子发芽达到高峰时(7 d)的发芽势G 和种子完全发芽(15 d)时的发芽率Gr。
数据统计分析:数据采用SPSS16.0 进行方差分析(GLM)、相关分析。方差分析时百分率做arcsinxi转换,家系间表型分化系数(Vs,t)的分析参照葛颂等[31]对群体的定义和分析。
3 结果与分析
3.1 球果大小和质量性状
方差分析结果表明,球果长、球果长/宽家系间、家系内差异达显著水平,球果宽家系间差异显著,家系内差异不显著(表1)。欧洲云杉家系球果平均长、球果平均宽、球果平均长/宽分别为(14.639 4 ±1.795 5)cm、(27.187 7 ±3.008 6)mm、(5.411 7±0.605 9),变幅分别为10.0 ~19.50 cm、14.69 ~36.77 mm、3.80 ~7.90,总体变异系数由大到小的顺序为球果长(12.27%)、球果长/宽(11.20%)、球果宽(11.07%)(表2)。球果长、球果宽、球果长/宽家系间的表型分化系数分别为89.01%、78.54%和94.95%(表1)。
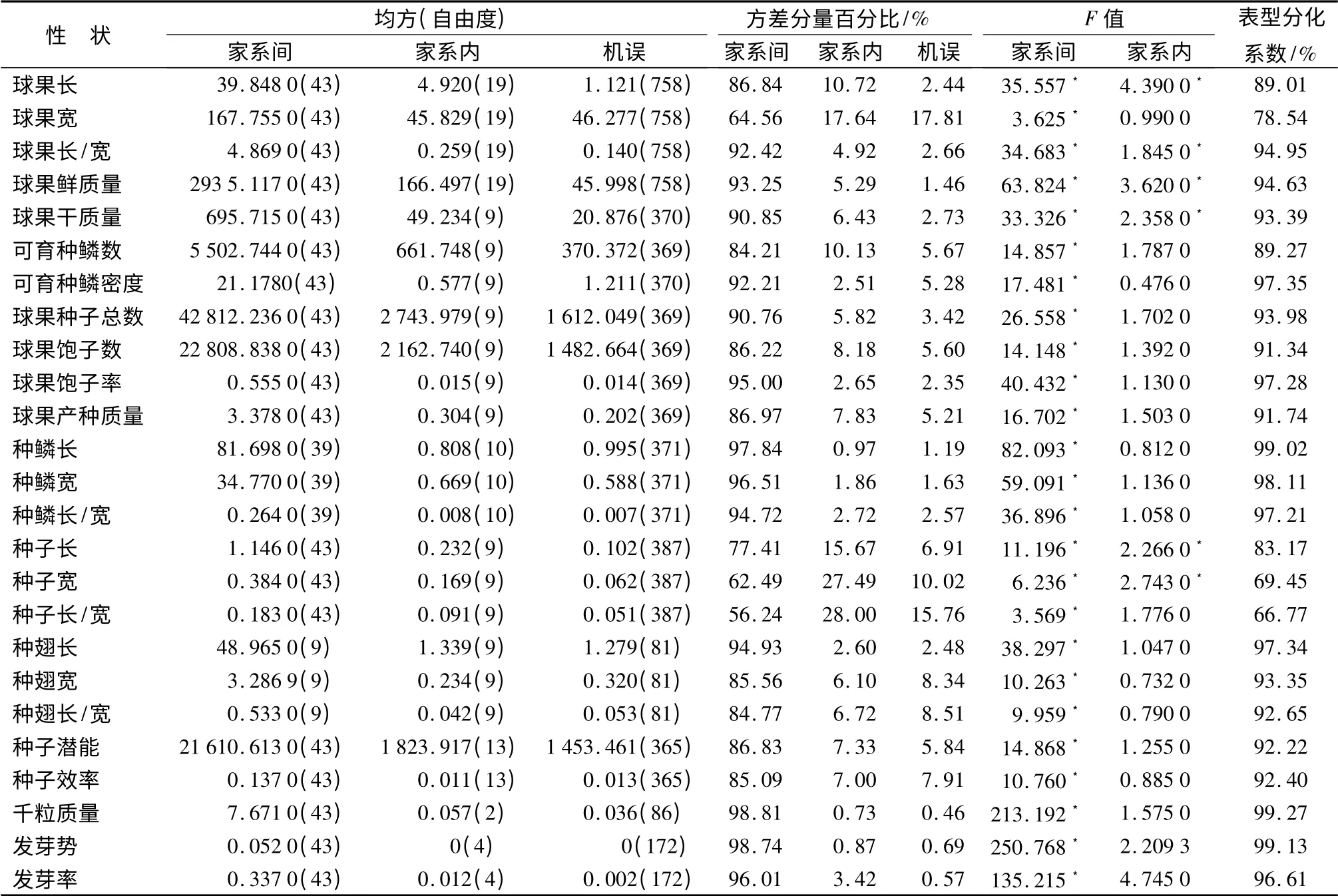
表1 欧洲云杉家系种实性状方差分析
方差分析结果表明,家系间、家系内球果鲜质量和干质量差异均达显著水平(表1)。家系间球果鲜质量平均为(47.951 9 ±14.128 9)g,变幅13.53 ~100.22 g;干质量平均为(34.538 9 ±9.487 6)g,变幅为14.69 ~68.30 g,总体变异系数由大到小的顺序为球果鲜质量(29.46%)、干质量(27.47%)(表2);家系间球果鲜质量、干质量的表型分化系数分别为94.63%和93.39%(表1)。
球果长、球果宽、球果长/宽、球果鲜质量、球果干质量性状家系间的方差分量远远大于家系内的方差分量,球果大小和质量性状的变异主要受到家系间遗传因素的控制;是母体营养状况和用于有性生殖投入的体现。家系内的差异受环境因子影响,如球果在树冠上的方位、冠层、光照、风力、授粉状况等。同一家系内环境对球果长的影响较球果宽的影响大,球果宽是较为稳定的性状。
3.2 球果可育种鳞、种子潜能、种子效率
方差分析结果,家系间可育种鳞数、种鳞密度、种子潜能、种子效率差异达显著水平,家系内差异不显著。家系间表型分化系数为89.27% ~97.35%(表1)。家系内的表型分化仅占2.65% ~11.73%,可见球果的这些性状主要受到家系强的遗传控制。
可育种鳞数、种鳞密度、种子潜能、种子效率的平均值分别为(204.777 3 ±30.001 8)个、(13.871 8±1.798 7)个·cm-1、(409.872 0 ±59.402 7)粒、(61.852 1 ±12.569 8)%;变幅分别为115.00 ~319.00 个、7.47 ~22.07 个·cm-1、240.0 ~638.0粒、5.07% ~90.00%;总体变异系数由大到小的顺序为种子效率(20.32%)、可育种鳞数(14.65%)、种子潜能(14.49%)、可育种鳞密度(12.97%)(表2)。
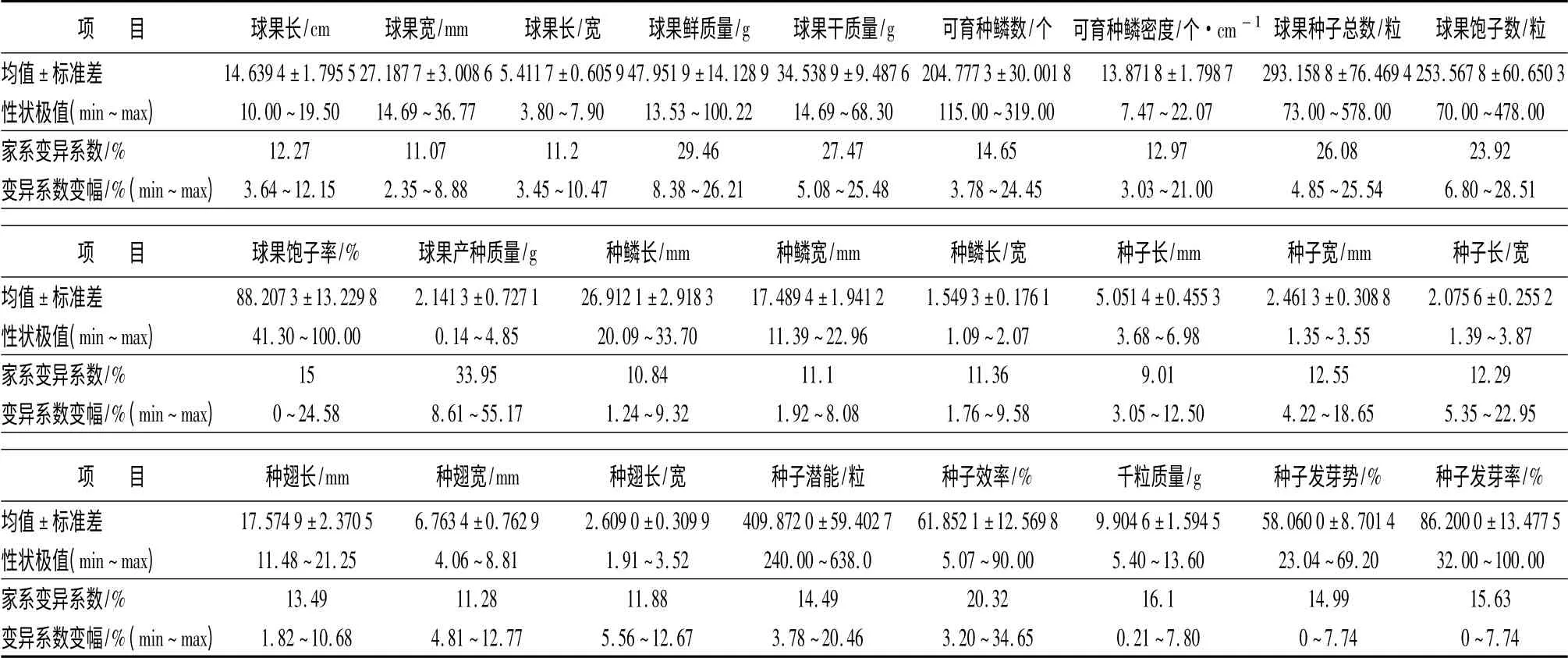
表2 家系种实性状均值和变异系数
家系间球果平均生产种子的潜在能力为409粒,最大可达到638 粒,种子潜能的变异系数最小,种子效率的变异系数最大。种子潜能是受家系遗传影响,家系间种子效率的差异受遗传和环境双重因子影响,如树体位置、花粉的传播方式、花粉量、风力、授粉效率等因素。种子效率平均为61.85%,因此,在可受期进行人工辅助授粉,可提高种子效率,同时也提高了球果生产种子量。
3.3 球果产种数与产种质量
方差分析结果表明,球果生产种子总数、饱子数、饱子率、球果产种质量家系间差异达显著水平,家系内差异不显著,家系间表型分化系数为91.34% ~97.28%(表1)。
家系间球果生产种子总数、饱子数、饱子率、球果产种质量的平均值为(293.158 8 ±76.469 4)粒、(253.567 8 ± 60.650 3)粒、(88.207 3 ± 13.229 8)%、(2.141 3 ±0.727 1)g,变幅分别为73.0 ~578.0 粒、70.0 ~478.0 粒、41.30% ~100.00%、0.14 ~4.85 g,总体变异系数由大到小的顺序为球果产种质量(33.95%)、种子总数(26.08%)、饱子数(23.92%)、饱子率(15.00%)(表2)。
球果生产种子总数是饱子数和空子数(有完整的种翅、种壳、色泽,与饱子相同)之和,饱子数是球果受精成功后发育成有胚珠、胚乳的胚珠数量;空子数的形成与营养状况、病虫危害、自交有关,是家系间交配系统、传粉效率、营养状况和环境等诸多因素的综合响应。球果生产饱子数的多少影响饱子率,从而影响球果产种质量。球果饱子率的变幅为41.30% ~100.00%,有些球果的饱子率可以达到100.00%,因此,可授期进行人工辅助授粉或改善环境和营养状况可提高授粉效率。在进行云杉种子园营建时,合理配置交配系统、减少自交,是提高球果饱子率的有效手段。
3.4 种子性状
3.4.1 种子大小
家系间种子平均长、种子平均宽、平均种子长/宽的总体平均值依次为(5.051 4 ±0.455 3)mm、(2.461 3 ±0.308 8)mm、2.075 6 ±0.255 2,变幅分别为3.68 ~6.98 mm、1.35 ~3.55 mm、1.39 ~3.87。总体变异系数由大到小的顺序为种子宽(12.55%)、种子长/宽(12.29%)、种子长(9.01%)(表2)。方差分析结果表明,种子长、种子宽家系间、家系内差异显著,种子长/宽家系间差异显著,家系内差异不显著。种子长、种子宽、种子长/宽家系间表型分化系数分别为83.17%、69.45%和66.77%(表1)。表明家系遗传因素对种子性状的影响占优势,其次受到家系内环境因素的影响。
3.4.2 种翅性状
种翅家系平均长、宽、长/宽分别为(17.574 9 ±2.370 5)mm、(6.763 4 ±0.762 9)mm、2.609 0 ±0.309 9,变幅分别在11.48 ~21.25 mm、4.06 ~8.81 mm、1.91 ~3.52,总体变异系数由大到小的顺序为种翅长(13.49%)、种翅长/宽(11.88%)、种翅宽(11.28%)(表2)。经方差分析发现,种翅性状在家系间差异显著,家系内差异不显著,表型分化系数为92.65% ~97.34%(表1)。表明种翅性状主要受家系强的遗传因素控制,环境因素的影响非常微弱。
经观察,云杉种子旋转飘落,种翅的大小影响种子传播速度、传播距离。对种子的定居、萌发、后期生长产生影响,这是其长期适应自然的结果。
3.5 种子发芽势、发芽率、千粒质量
家系间种子平均发芽势和发芽率分别为(58.060 0 ±8.701 4)%和(86.200 0 ±13.477 5)%,发芽势和发芽率的总体变异系数分别为14.99%和15.63%(表2)。方差分析结果显示,种子发芽势、发芽率家系间差异达显著水平,表型分化系数分别为99.13%和99.61%(表1),表明发芽势和发芽率主要受家系间强的遗传基因控制。经解剖未发芽的种子多数为空粒。千粒质量是种子的品质反映,欧洲云杉平均千粒质量为(9.904 6 ±1.594 5)g,变幅为5.40 ~13.60 g,总体变异系数为16.10%(表2)。家系间千粒质量的方差分析结果显示,家系间千粒质量差异达显著水平,表型分化系数为99.27%(表1)。千粒质量主要受强的家系遗传因素控制。因此,在杂交育种、种子园营建时以千粒质量为主要选择指标,选择千粒质量大的家系,有利于子代后期生长。
3.6 种实性状间的相关性
相关分析结果表明,球果产种质量与可育种鳞数显著相关,相关系数R =0.376(P =0.012)、与种子总数、种子效率呈极显著正相关关系,相关系数分别为R=0.471(P =0.001)、R =0.707(P =0)。可育种鳞与球果长、种子总数、饱子数呈极显著正相关关系,相关系数分别为R =0.495(P =0.001)、R=0.720(P =0)、R=0.632(P =0),与球果宽呈显著正相关关系,相关系数R =0.309(P =0.041)。种子发芽势、发芽率均与千粒质量呈极显著正相关,相关系数分别为R =0.886(P =0)和R =0.883(P=0),发芽势与发芽率也呈极显著正相关,相关系数为R=0.973(P =0)。球果长/宽与球果宽、种子长/宽与种子宽呈极显著负相关关系。除球果长/宽与千粒质量,种子长/宽与球果产种质量、千粒质量、球果长、球果宽、种子长不相关外,其他种实性状间呈极显著或显著正相关关系(表3)。
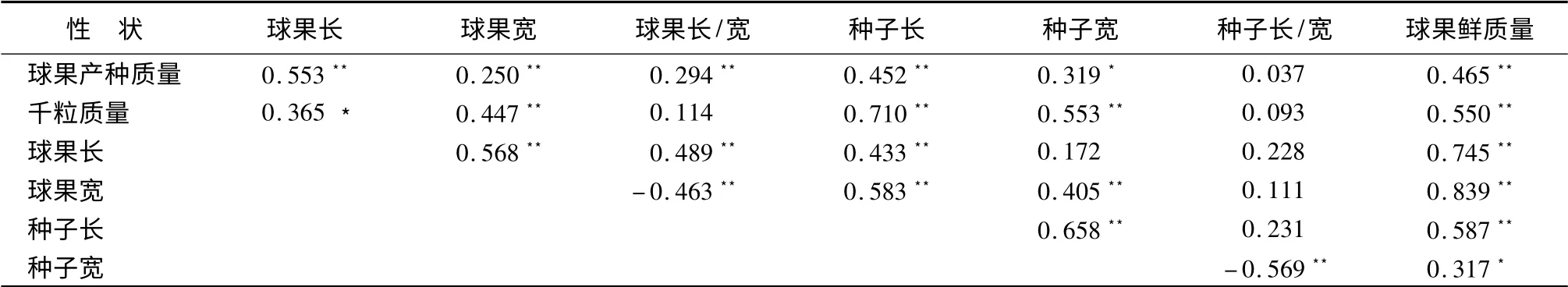
表3 种实性状间的相关分析
3.7 优良家系的筛选
种实性状间呈显著或极显著相关,种实性状受到家系强的遗传控制和微弱环境因子影响。千粒质量是种子品质的反映,是重要的种实性状之一,千粒质量的大小表征了种子内营养物质的多少,与种子发芽势、发芽率极显著相关,而且影响子代后期的生长及适应性,因此,以千粒质量的大小为选择依据,选择千粒质量≥11.05 g 的10 个家系作为营建种子园、多目标杂交育种的亲本,其他性状也将得到改良。
4 结束语
研究发现,引种欧洲云杉44 个家系在25 个种实性状中球果长,球果长/宽,球果鲜质量、干质量,种子长、宽丰富的遗传变异不仅存在于家系间,同样也存在于家系内。其余种实性状家系间具有丰富的遗传变异,家系内的变异不显著。种实性状家系间的表型分化系数为66.77% ~99.27%,表明种实性状主要受家系间强的遗传控制,其次受到微弱的环境影响;这与前人[19]对黑云杉、欧洲云杉的研究结论具有相似性。
松科球果大小与受精状况有关,受精状况好则球果就大[35]。受精不仅受到家系间的遗传控制,还与环境因子有关,由于云杉为风媒传粉,球果在树冠上的方位、冠层、树体营养状况、风力风向以及病虫危害等都影响球果和种子的生长发育,从而影响到球果鲜质量、干质量等性状的变异。球果长,种子长、宽,球果鲜质量、干质量等性状不仅受到家系间强的遗传控制,还受到家系内环境因子的影响。
球果性状是决定种子产量的决定因素,只要测得球果长(或球果鲜质量)和种子效率就可预测球果产种量,从球果产种量可以准确预测家系乃至种子园的产种量。欧洲云杉球果平均生产种子(293.158 8 ±76.469 4)粒,变幅为73.0 ~578.0 粒。饱子率的变幅较大,为41.30% ~100.00%。除环境因素外,自交是产生空子的主要原因[33]。由于云杉雌雄同株,球花量H(雌)∶ H(雄)=1∶ (4.5 ~50)[36],云杉花粉有双气囊,易于飞翔,所以产生自交的几率大。所以在种子园或母树林,为了提高云杉饱子率,要在可受期进行人工辅助授粉,在种子园营建时合理配置,减少自交。欧洲云杉种实性状丰富的遗传变异主要受家系间强的遗传因素控制,其次受到微弱环境因子影响。欧洲云杉种实性状间具有显著或极显著相关关系。千粒质量是种子品质的反映,是重要的种实性状之一,欧洲云杉千粒质量的平均值为(9.904 6 ±1.594 5)g。欧洲云杉家系间千粒质量的变幅为5.40 ~13.60 g。千粒质量的大小表征了种子内营养物质的多少,其性状变异的大小影响到种子的传播和萌发、幼苗的存活、子代后期的生长及适应性。根据欧洲云杉家系千粒质量的大小,选择了10 个千粒质量较大的家系作为营建种子园、多目标杂交育种的亲本。
家系间丰富的遗传变异具有较大的种质价值和遗传改良潜力。引种云杉种实性状上变异丰富,有广阔的遗传多样性,可以作为育种用的材料,不会影响子代的遗传幅度。
[1] 张立攻,张闯含.欧洲云杉引种研究[J].河北林学院学报,1995,10(2):122-126.
[2] 祁万宜,应中华,王军辉,等.几种针叶树引种试验研究[J].河北农业大学学报,2004,40(1):38-44.
[3] 马常耕.世界云杉无性系林业发展现状[J].世界林业研究,1993,11(6):24-31.
[4] Neuffer M G,Ehcoe J R.Paraffin oil technique for treating corn pollen with chemical mutagents[J].Maydica,1978,22:21-28.
[5] Ruby J L.The correspondence between genetic,morphological and climatic variation patter in scot pine[J].Silvae Genetica,1967,16(2):50-56.
[6] Wheeler N C,Guries R P.Population structure,genetic diversity,and morphological variation in Pinus contorta Dougl[J].Can J For Res,1982,12:595-606.
[7] 毛建丰,李 悦,刘玉军,等.高山松种实性状与生殖适应性[J].植物生态学报,2007,31(2):291-299.
[8] Josefa L,Juan A D,Ana O O,et al.Production and morphology of fruit and seeds in Genisteae (Fabaceae)of Southwest Spain[J].Bot J Linnean Soc,2000,132(2):97-120.
[9] 王军辉,顾万春,万军,等.桤木不同种源球果及种子性状的遗传变异[J].东北林业大学学报,2006,34(2):1-4,17.
[10] Raquel G R,Keith R P,Malcolm E R,et al.Effect of seed size and test colour on saponin content of Spanishlentil seed[J].Food Chem,1997,58(3):223-226.
[11] Latif K M,Putul B,Uma S,et al.Seed germination and seedling fitness in Mesua ferrea L.in relation to fruit size and seed number per fruit[J].Acta Oecol,1999,20(6):599-606.
[12] Eriksson O.Seed size variation and its effect on germination and seedling performance in the clonal herb Convallaria majalis[J].Acta Oecol,1999,20(1):61-66.
[13] Khurang E,Singh J S.Influence of seed size on seedling growth of Albizia procera under different soil water levels[J].Ann Bot,2000,86:1185-1192.
[14] Simons A M,Johnston M O.Variation in seed traits of Lobetia inflata (Campanuhceaee):sources and fitness consequences[J].American Journaf of Botany,2000,87(1):124-132.
[15] 陈益泰,李桂英,王惠雄.恺木自然分布区内表型变异的研究[J].林业科学研究,1999,12(4):379-385.
[16] Barzdajn W.Intra population diversity in spruce (Picea abies(L.)Karst.)in the mountains[J].Sylwan,1996,140(3):39-46.
[17] Barzdajn W.An assessment of diagnostic value of morphologicaltraits of Norway spruce (Picea abies (L.)Karst.)cones for discrimination of spruce provenances[J].Sylwan,1996,140(9):61-75.
[18] Bergmann F.The allelic distribution at an acid phosphatase locus in Norway spruce (Picea abies)along similar climatic gradients[J].Theoretical and Applied Genetics,1978,52(2);57-64.
[19] Borghetti M,Gianninir M.Geographic variation in cones of Norway spruce (Picea abies (L.)Karst.)[J].Silvae Genetica,1988,37(5/6):178-184.
[20] Khalil M A K.Genetic variation in estern white spruce (Picea glauca (Moench)Voss)populations[J].Can J For Res,1985,15:444-452.
[21] Li Peng,Beaulieu J,Bousquet J.Genetic structure and patterns of genetic variation among populations in eastern white spruce(Picea glauca)[J].CanadianJournal of Forest Research,1997,27(2):189-198.
[22] HO R H.Seed-cone receptivity and seed production potential in white spruce[J].For Ecol Manage,1984,9:161-171.
[23] HO R H.Effect of recepited pollination upon filled seed in white spruce[J].Can J For Res,1985,15:1195-l197.
[24] Khalil M A K.Genetic of cone morphology of black spruce (Picea mariana (Mill).B.S.P)in Newfoundland,Canada[J].Silvae Genetica,1984,33:101-109.
[25] Khalil M A K.Genetic variation in black spruce (Picea mariana(Mill)B.S.P.)in Newfoundland[J].Silvae Genetica,1984,24(4):88-96.
[26] Yeh F C,EI-kassaby Y A.Enzyme variation in natural populations of sitka spruce[J].Can J For Res,1980(10):415-422.
[27] 王娅丽,李毅.祁连山青海云杉天然群体的种实性状表型多样性[J].植物生态学报,2008,32(2):355-362.
[28] 臧润国,刘贵峰,巴哈尔古丽·阿尤甫.天山云杉球果大小性状的地理变异规律[J].林业科学,2009,45(2):27-32.
[29] 吴远伟,罗建勋,胡庭兴,等.川西云杉天然林分特点和种内群体分化的研究[J].西北农林科技大学学报:自然科学版,2008,36(9):81-86.
[30] 罗建勋,顾万春.云杉天然群体表型多样性研究[J].林业科学,2005,41(3):66-73.
[31] 葛颂.同工酶与林木群体遗传变异研究[J].南京林业大学学报,1988(1):68-77.
[32] Lagercrantz U,Ryman N.Genetic structure of Norway spruce(Picea bies):concordance of morphological and allozymic variation[J].Evolution,1990,44(1):38-53.
[33] 王晓茹,沈熙环.对由胚珠败育和空粒引起油松种子园减产的分析[J].北京林业大学学报,1999,11(3):60-65.
[34] 吴彦,刘庆,何海,等.光照与温度对云杉和红桦种子萌发的影响[J].应用生态学报,2004,15(12):2229-2232.
[35] 张恒庆,安利佳,祖元刚.天然红松种群形态特征地理变异的研究[J].生态学报,1999,19(6):932-938.
[36] 马建伟,胡勐鸿,靳景春,等.云杉开花生物学和杂交技术[J].种子,2010,29(11):1-5.
