杉木老龄林群落的种—面积关系1)
2014-03-06蔡锰柯林开敏赵长存郑文辉刘圣恩
蔡锰柯 林开敏 赵长存 郑文辉 刘圣恩
(福建农林大学,福州,350002) (国家林业局杉木工程技术研究中心) (福建农林大学)
何宗明 郭玉硕
(国家林业局杉木工程技术研究中心) (福建农林大学)
种—面积关系的研究在群落生态学中有着悠久的历史,虽然由于研究区域的差异性,样方内物种数目随取样面积的扩大而增加的速度也各不相同[1-2],但从总体上看,在面积扩大的初期阶段,物种种类增加的速度较快,到了中后期逐步减缓甚至出现物种种类保持稳定的变化趋势。植物群落生态学家将这种变化的规律称为“生态学中少有的定理之一”[3],而将变化规律所形成的曲线叫做种—面积曲线。从植物的种—面积曲线首次绘出至今[4],许多学者为探究物种与面积之间的关系付出了巨大的努力。种—面积关系的定量化研究,最早由Arrhenius[5-6]在1921—1923年提出。他采用巢式系列样方进行试验样地的设置并统计出不同面积样方中的物种数量。在数据的整理过程中发现,物种种类和样地面积呈现出幂函数关系进而提出S=c×Az的关系式。随着种—面积理论的不断发展,许多数学和生态学家尝试运用数学表达式或数学方法构造出同实验或观察数据吻合的种—面积模型[7],乔秀娟[8]等在天山南北坡植物种—面积关系的研究中采用logS=logc +zlogA,要元媛[1]等认为采用是描述山西霍山植物群落种—面积关系较好的模型。经过生态学家们的不断探索至今,用以呈现不同类型的物种数目与样地面积间关系的曲线模型及关系式已经被大量提出[9],成为一种既能反映物种数目和取样面积间关系量化的研究方式[10],又能用以衡量群落中的物种组成特征和结构变化规律的测度方法。为不同群落中物种多样性、结构多样性的调查提供理论基础和研究依据[11-12]。
杉木(Cunninghamia lanceolata (lamb.)Hook.)在我国有着悠久的栽培历史,也是我国最重要的速生用材树种。由于长期以来不合理的育林制度和杉木自身的生长特性,造成杉木林群落出现物种结构单一和土壤退化等现象,不仅直接制约杉木林的可持续发展,也破坏了群落中物种的多样性[13-14]。目前国内关于杉木老龄林群落虽然已有一些研究[15-16],但有关该群落在种—面积关系方面的研究则尚未见报道。本研究以现存量稀少的杉木老龄林为研究对象,通过建立60 m×60 m 的大样地开展乔木和灌木植物的全面调查,分析杉木老龄林群落物种数目和取样面积间的关系。同时依据调查数据对不同的种—面积曲线模型进行拟合,筛选出适合分析杉木老龄林群落中植物物种数目和面积关系的最佳模型,并确定该群落的最小取样面积,为杉木林群落生物多样性恢复和重建提供科学的理论依据和实践指导。
1 研究区概况
试验林地设在福建省南平市延平区的福建农林大学西芹教学林场花竹沟。从地理位置看,福建省南平市是武夷山脉北段东南侧,因其独特的气候特点,植物年均生长周期可达300 d,属于杉木的中心产区。土壤类型主要以红壤、黄壤和红黄壤为主,土层也较厚。由于境内复杂的地形条件,该区域的成土母岩包括沉积岩和变质岩。
由于地处中国东南部湿润森林区中亚热带常绿阔叶林带,该区域属中亚热带季风气候。夏季日照时间较长,冬季日照时间较短,年均日照时数为1 590 h。全年雨量明显,年均降水量为1 735 mm,多集中于5、6月份,具有温暖湿润,多雨多雾等气候特征。年平均温度为19.3 ℃,1月平均温度(极端最低为-5.8 ℃)较低,7月平均温度(极端最高为41 ℃)较高;全年无霜期302 d,极少下雪;年均风速为1.1 m·s-1。独特的气候条件使其十分适宜杉木人工林的生长。
本研究课题组在对杉木树种长期的研究过程中,针对杉木人工林所存在的一系列生态问题,结合国内外学者所提出的关于人工林的新理论和新方法,对杉木人工林群落实施近自然改造和调控,逐步建立起一些近自然的杉木林经营模式,其中杉木老龄林群落(林龄约为73 a)就是在一代杉木人工成熟林基础上,通过延长杉木轮伐期(杉木轮伐期一般为25 a),并以实施保护和促进天然更新为主要技术,逐步形成的一种近自然的杉阔混交经营模式。
2 研究方法
2.1 样地设置及调查方法
2013年3月,在福建省南平市的福建农林大学西芹教学林场花竹沟中的杉木老龄林内设置一个面积为3 600 m2(60 m×60 m)的大样地。并将标准地划分为144 个5 m ×5 m 的样方网格。调查时以最小的样方网格为基本单位,调查记录每个5 m ×5 m小样方内所有出现木本植物的种类名称、个体数量、胸径(地径)、高度、冠幅等指标,并对每一木本植物个体进行挂牌和定位,并测量其坐标。
2.2 种—面积关系研究的样方构建方式
Scheiner[17]在对种—面积关系的研究过程中发现,种—面积关系的构建方式包括组合样方法、巢式样方法及隔离生境。本研究在综合分析杉木老龄林群落的实际情况后,采用巢式样方法(图1)和邻域随机连续组合样方法(图2)对3 600 m2的样地进行组合[7],并比较2 种不同的样地组合方式所拟合出的种—面积模型。
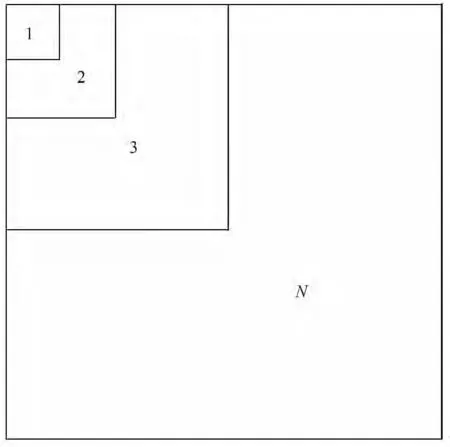
图1 巢式样方法
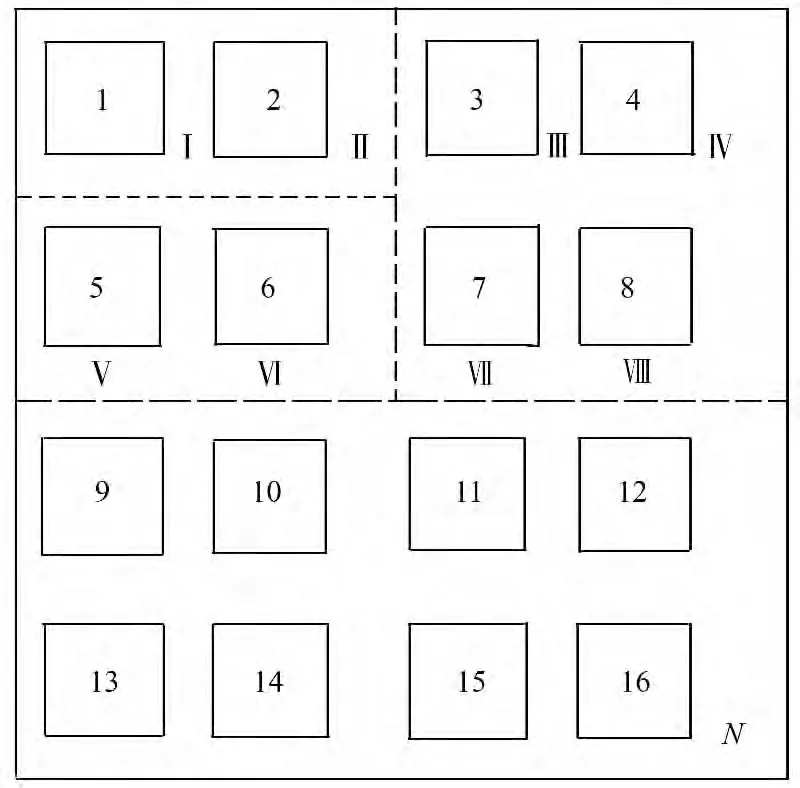
图2 组合样方法
2.3 种—面积数学模型的选取及最小取样面积的确定
本研究选取10 种常见的种—面积曲线作为杉木老龄林群落的拟合模型:
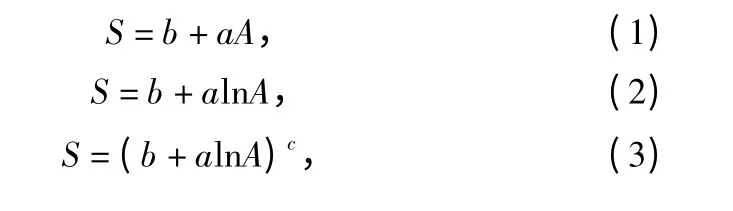
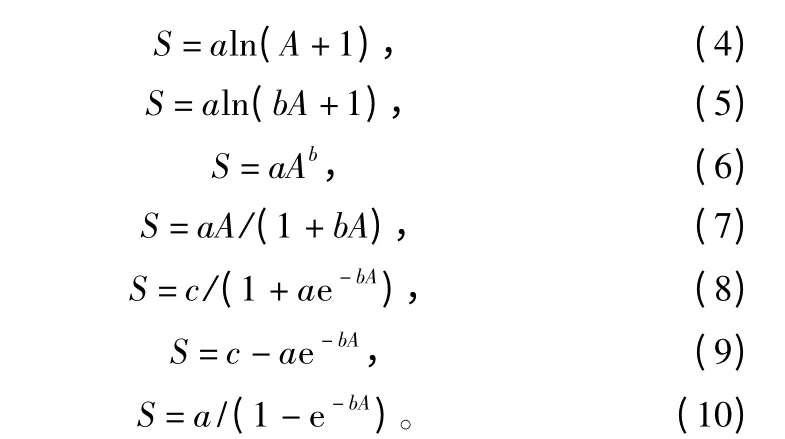
其中:公式(1)—公式(6)模型的曲线为非饱和曲线,公式(7)—公式(10)模型的曲线为饱和曲线。在表达式中A 用以指代样地面积,S 表示对应样地的物种数目。a、b、c 均为种—面积模型中的参数。
2.4 数据处理
不同样地面积条件下的物种数目采用Excel2007 进行数据初步处理,模型的拟合采用SPSS17.0 进行。
3 结果与分析
3.1 不同样方构建方式的种—面积关系
3.1.1 不同样方构建方式的种—面积曲线比较
根据144 个样方的资料对杉木老龄林群落所有的木本植物进行统计并绘出种—面积曲线图,由图3可知杉木老龄林群落中物种数目随样地面积的增大而增加。从组合样方法的种—面积关系来看,样地面积在(25 ~500)m2的过程中,物种数目随样地面积的增大,呈现快速增加的趋势;从(500 ~1 125)m2,物种数目进入一个较快的增长阶段;从(1 125 ~3 300)m2,物种数目进入缓慢增加阶段;从(3 300~3 600)m2的过程中,物种的数目已经没有变化,基本趋于稳定的阶段。另一方面,以巢式样方法的种—面积关系来看,样地面积在(25 ~400)m2的过程中,物种数目随样地面积的增大,呈快速增加的趋势;从(400 ~1 225)m2的范围,物种数目进入一个较快增长的阶段;从(1 225 ~3 025)m2,物种的数目进入缓慢增加阶段;到了3 025 m2直至最后的3 600 m2的样地中,物种的数目已经不再增加。因此,从两种方法的种—面积曲线来看,(1 000 ~1 200)m2范围可作为老龄杉木林群落的最小取样面积。
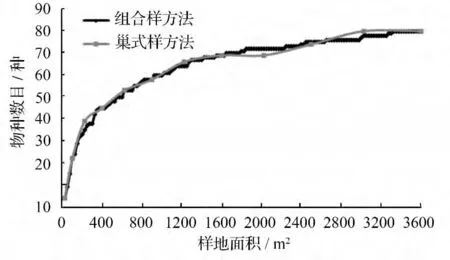
图3 杉木老龄林群落中不同构建方式的种—面积曲线
此外,比较两种方法的种—面积曲线,巢式样方法与组合样方法的种—面积曲线的变化趋势基本较为一致,均可用以描述杉木老龄林群落的种—面积关系。
3.1.2 不同样方构建方式种—面积数学模型的拟合与评价
依照巢式样方法和组合样方法对杉木老龄林144 个5 m×5 m 的小样方进行重新排序和统计,初步画出两种样方构建方式的种—面积曲线,然后采用10 个数学模型对种—面积曲线进行拟合(表1)。从巢式样方法和组合样方法的种—面积曲线模型拟合结果来看,模型(2)、(6)及(7)的拟合结果均较好(R2>0.85),其次为模型(3)、(4)、(1)、(5)、(10)(0.69 <R2<0.81),而采用模型(8)、(9)对种—面积曲线进行拟合的效果均较差(R2<0.41)。说明(模型(2))、(模型(7))及(模型(6))明显优于其余7 个种—面积数学模型,可用于描述杉木老龄林群落的种—面积关系。
对比两种方法的种—面积模型拟合结果(表1)可知:不同方法拟合的数学模型R2值及模型参数的值差异不大,说明两种方法均可用于描述杉木老龄林群落的种—面积关系。但从表1中还可看出,同一数学模型的两种方法拟合度还是略有差异,在模型(2)、(6)、(7)中,巢式样方法构建数学模型的拟合度略高于组合样方法,在模型(1)、(3)、(4)、(5)、(8)、(9)、(10)中,组合样方法构建数学模型的拟合度则略高于巢式样方法。

表1 两种方法对杉木老龄林群落的种—面积模型拟合结果
3.2 不同层次植物的种—面积关系
依照巢式样方法初步画出杉木老龄林群落乔木层及灌木层的种—面积曲线,从图4可知,乔木层与灌木层物种的数目均随样地面积的增大而增加,但乔木层与灌木层的种—面积曲线变化趋势存在一定差异。从乔木层的种—面积关系来看,样地面积在(25 ~400)m2的过程中,物种数目随样地面积的增大,呈现出快速的增加趋势;从(400 ~900)m2,物种数目的增长进入较快的阶段;从(900 ~2 500)m2的过程,物种数目的增长进入较为缓慢的阶段,从(2 500 ~3 600)m2的过程中,物种的数目已经没有变化。另一方面,从灌木层的种—面积关系来看,样地面积在(25 ~400)m2的过程中,物种数目随样地面积的增大,呈较为快速的增长变化;从(400 ~1 225)m2,其物种数目进入较快增长的阶段;从(1 225 ~3 025)m2,物种数目的增加基本进入较为缓慢的阶段;到了(3 025 ~3 600)m2的样地中,物种的数目已经不再增加。因此,从乔、灌木层的种—面积曲线来看,(700 ~900)m2范围可作为乔木层的最小取样面积,(1 200 ~1 300)m2则可作为灌木层的最小取样面积。
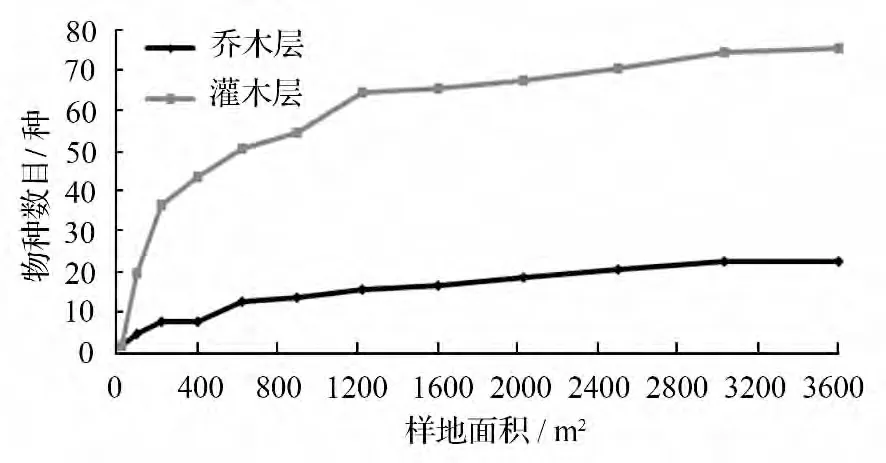
图4 灌木层与乔木层的种—面积曲线
基于模型(1)—(10)对杉木老龄林群落乔木层和灌木层的种—面积关系进行拟合,其结果可知:从乔木层的拟合结果来看,模式(2)(R2=0.937 3)、(6)(R2=0.979 7)具有较高的拟合度,其次为模型(1),其余模型对乔木层种—面积关系的拟合度则较差(R2<0.72);而从灌木层的拟合结果来看,模型(2)具有较高的拟合度,其R2的值为0.994 4,其次为模型(6),其R2的值为0.815 1,其余模型对灌木层种—面积关系的拟合度则较差(R2<0.72)。

表2 杉木老龄林各层种—面积拟合结果
4 结论与讨论
巢式样方法与组合样方法绘出的群落木本植物的种—面积曲线变化趋势较为一致,其物种数目均随样地面积的增大而增加。在取样面积增加初期,物种数目均呈快速或较快增长趋势,随后呈较为缓慢的增长,最后趋于稳定。此外,从两种方法的种—面积曲线来看,(1 000 ~1 200)m2的范围可作为老龄杉木林群落的最小取样面积。
本文选择10 个数学模型对杉木老龄林群落的种—面积曲线进行拟合,结果表明:模型(2)、(6)、(7)的拟合度较高,可用于描述杉木老龄林群落的种—面积关系。这与张蕊[4]等在研究东祁连山亚高山草甸不同样地的种—面积曲线拟合结果及刘灿然等[18]在研究北京东灵山地区植物群落种—面积曲线的拟合结果有所不同。这可能是由于本实验的研究对象为杉木老龄林群落木本植物,群落内乔、灌木层的复杂程度较高,林下植被也更丰富。另一方面,本研究是建立在一次性调查3 600 m2样地内物种数目的数据基础上,而研究北京东灵山地区植物群落与东祁连山亚高山草甸的种—面积关系时,其样地面积在(64 ~2 500)m2的范围内。因此,本文研究的杉木老龄林群落种—面积关系关于尺度的依赖性与东祁连山和北京东灵山有所不同,这可能是导致本文研究结果产生差异的主要原因之一。此外,植物群落在空间分布上的差异也可能导致同其它地区的研究结果不同。
乔、灌木层的种—面积关系研究结果表明:在取样面积增加初期,乔木层及灌木层物种数目均呈快速或较快增长,随后进入较为缓慢的增长阶段,最后趋于稳定,但乔木层与灌木层的种—面积曲线变化趋势存在一定差异。此外,(700 ~900)m2可作为乔木层的最小取样面积,(1 200 ~1 300)m2可作为灌木层的最小取样面积。从不同数学模型的拟合结果而言,模型(2)、(6)、(1)对乔木层种—面积曲线的拟合度较高(R2>0.87),模型(2)、(6)则对灌木层的种—面积曲线拟合度较高(R2>0.81)。这可能是因为在延长一代杉木人工成熟林的轮伐期之后,杉木老龄林是以实施保护和促进天然更新为主要经营措施,对林地的人工扰动较少,有利于林地内天然阔叶树种及灌木的天然更新和侵入,但二者侵入和生长所需的时间不相同,乔木层阔叶树种的天然更新需要较长周期,而灌木层物种则可以在较短的时间内恢复与生长。因此,在选取模型时,应根据群落类型、林下植被的生长及更新等实际情况选择合适的模型。
由于杉木老龄林群落草本植物物种数目及数量较大,调查需要很大的工作量,因此本文仅对样地内木本植物进行全面调查,至于草本植物还有待今后进一步研究。在对杉木老龄林的物种数目进行调查后,仅通过得到的数据拟合10 个种—面积模型,在本文选取的模型之外,是否还有能够反映杉木老龄林群落中种—面积关系的模型,其拟合结果与本研究结果相比是否具有更好的拟合度或预测结果,其结果有待进一步的探讨。
[1] 要元媛,闫明,毕润成.山西霍山植物群落种—面积曲线与物种多样性的关系[J].生态学杂志,2013,32(1):39-44.
[2] 沈有信,刘文耀,崔建武.滇中喀斯特森林土壤种子库的种—面积关系[J].植物生态学报,2007,31(1):50-55.
[3] Lomolino M V.Ecology’s most general,yet protean pattern:the species-area relationship[J].Journal of Biogeography,2000,27:17-26.
[4] 张蕊,陈军强,侯尧宸,等.亚高山草甸植物群落植物物种数与取样面积的关系[J].生态学杂志,2013,32(9):2268-2274.
[5] Arrhenius O.Species and area[J].Journal of Ecology,1921,9:95-99.
[6] Arrhenius O.Statistical investigations in the constitution of plant associations[J].Ecology,1923,4:68-73.
[7] 唐志尧,乔秀娟,方精云.生物群落的种—面积关系[J].生物多样性,2009,17(6):549-559.
[8] 乔秀娟,唐志尧,安尼瓦尔·买买提,等.天山南北坡植物种—面积关系[J].干旱区研究,2011,28(1):54-59.
[9] 姜俊.吉林蛟河针阔混交林种—面积关系及种间联结性研究[D].北京:北京林业大学,2012.
[10] 汤孟平,周国模,施拥军,等.天目山常绿阔叶林群落最小取样面积与物种多样性[J].浙江林学院学报,2006,23(4):357-361.
[11] 吴晓莆,朱彪,赵淑清,等.东北地区阔叶红松林的群落结构及其物种多样性比较[J].生物多样性,2004,12(1):174-181.
[12] 代力民,王青春,邓红兵,等.二道白河河岸带植物群落最小面积与物种丰富度[J].应用生态学报,2002,13(6):641-645.
[13] 林开敏,章志琴,叶发茂,等.杉木人工林下杉木、楠木和木荷叶凋落物分解特征及营养元素含量变化的动态分析[J].植物资源与环境学报,2010,19(2):34-39.
[14] 王祖华,李瑞霞,郝俊鹏,等.间伐对杉木人工林不同根序细根形态的影响[J].东北林业大学学报,2011,39(6):13-19.
[15] 郭恩莹,吴保国,张瀚,等.杉木人工林缓冲区类竞争指数[J].东北林业大学学报,2012,40(10):34-42.
[16] 高明,朱玉杰,董希斌,等.采伐强度对大兴安岭用材林生物多样性的影响[J].东北林业大学学报,2013,41(8):18-21.
[17] Scheiner S M.Six type of species-area curve[J].Global Ecology Research,2003,3:361-367.
[18] 刘灿然,马克平,于顺利,等.北京东灵山地区植物群落多样性研究:种—面积曲线的拟合与评价[J].植物生态学报,1999,23(6):490-500.
