Effect of stripe rust on the yield response of wheat to nitrogen
2014-02-28*
*
aClimate Change Cluster(C3),Faculty of Sciences,University of Technology,Sydney,Australia
bNSWDepartment of Primary Industries,Tamworth,NSW 2340,Australia
cSchoolof Environmental and Rural Science,University of New England,Armidale,NSW 2351,Australia
dCooperative Research Centre for Spatial Information,P.O.Box 672,Carlton South,VIC 3053,Australia
ePrecision Agriculture Research Group,University of New England,Armidale,NSW 2351,Australia
Effect of stripe rust on the yield response of wheat to nitrogen
Rakhesh Devadasa,e,*,Steven Simpfendorferb,David Backhousec,David W.Lambd,e
aClimate Change Cluster(C3),Faculty of Sciences,University of Technology,Sydney,Australia
bNSWDepartment of Primary Industries,Tamworth,NSW 2340,Australia
cSchoolof Environmental and Rural Science,University of New England,Armidale,NSW 2351,Australia
dCooperative Research Centre for Spatial Information,P.O.Box 672,Carlton South,VIC 3053,Australia
ePrecision Agriculture Research Group,University of New England,Armidale,NSW 2351,Australia
A R T I C L E I N F O
Article history:
Received 17 February 2014
Received in revised form 4 May 2014
Accepted 5 May 2014
Available online 23 May 2014
Puccinia striiformis
Nitrogen(N)is the most important fertiliser element determining the productivity of wheat. N nutrition is known to affect the levelof stripe rustinfection,with higher N associated with increased disease severity.Stripe rust,caused by Puccinia striiformis f.sp.tritici,is a major yield-limiting disease of wheat in Australia.This paper describes experiments designed to investigate the agronomic response to the interaction of various levels of N application and stripe rust severity in wheat varieties differing in response.Experimental plots were established in crop seasons 2006 and 2007 on the Liverpool Plains of northern NSW,Australia. Yield,biomass,grain protein content(GPC)and harvest index(HI)data were recorded. Increased rates of N increased the severity of stripe rust during grain filling.Napplication also increased yield and GPC in all varieties in both years.Stripe rust reduced the yield of the rust-susceptible wheat varieties,and GPC and proportion of added N recovered in the grain were also reduced in one yearbutnotthe other.Itwas evidentfromourexperimentthatstripe rust caused yield loss accompanied by either no change or reduction in GPC,indicating that the totalamount of Nentering the grain was reduced by stripe rust.The effects ofstripe ruston N yield are most likely associated with reduced uptake of N during grain filling.
©2014 Crop Science Society of China and Institute of Crop Science,CAAS.Production and hosting by Elsevier B.V.Allrights reserved.
1.Introduction
Of the three main rusts affecting wheat,stripe rust,caused by Puccinia striiformis f.sp.tritici(Pst),is the one that has proved the most difficult to manage in Australia.There are a limited number ofresistance genes available in adapted varieties,and new pathotypes that overcome the most widely deployed genes have arisen at frequent intervals.Outbreaks of allthree wheat rusts are highly dependent on weather conditions,with management relying on a combination of plant resistance,
*Corresponding author at:Climate Change Cluster(C3),Faculty ofSciences,University ofTechnology,Sydney,Australia.Tel.:+61 2 9514 8351.
E-mailaddresses:rakhesh.devadas@uts.edu.au(R.Devadas),steven.simpfendorfer@dpi.nsw.gov.au(S.Simpfendorfer), dbackhou@une.edu.au(D.Backhouse),dlamb@une.edu.au(D.W.Lamb).
Peer review under responsibility of Crop Science Society of China and Institute of Crop Science,CAAS.reducing“environmental risk”factors and the tactical application of fungicides if required.One important aspect of environmental risk is that associated with nitrogen management.
Nitrogen(N)nutrition is known to affect the level of stripe rust infection,with higher N associated with increased disease severity[1,2].Different mechanisms have been suggested to be involved in this response.Some studies suggest that increased crop density and canopy density associated with N fertilisation creates a more favourable microclimate for stripe rust development[2,3].Other studies suggest that the effect of N on stripe rust is mediated via increased N content of the host tissue acting as a substrate for pathogen growth,rather than via changes in canopy microclimate[4,5].
Diseases can also affect the way in which the crop uses nitrogen[6].In general,controlling rusts with fungicides increases the protein content of wheat grains.The mechanisms for this are uncertain,but it has been suggested that rusts have a greater proportional effect on nitrogen mobilisation into the grain than on the supply of photosynthate[6].
Adding nitrogen to a wheat crop in the presence of stripe rust could thus increase the severity of the disease,and the disease itself could then reduce the amount of nitrogen exported in the grain.Understanding the interaction of these factors is important in assessing the productivity impacts of rust management,namely,yield and quality(protein).
This paper describes replicated plot experiments designed to investigate the agronomic response(grain yield and quality) resulting from the interaction of various levels of N application and stripe rust severity in wheat varieties of differing levels of response.
2.Materials and methods
2.1.Experimental plot design
Field experiments were conducted over two consecutive seasons at the Breeza Research Station(New South Wales Department of Primary Industries)located on the Liverpool Plains of northern New South Wales(NSW),Australia(150°25′31″E and 31°10′54″S).Plots were sown with varieties Baxter, Ellison and Hybrid Mercury(HM)in 2006.In 2007,varieties Ellison and H45 were grown.Among these varieties,HM and H45 were considered highly susceptible,Baxter moderately resistant and Ellison resistant to pathotype(134 E16 A+),which was the dominant pathotype in eastern Australia during the years in which the experiments were conducted.In both years wheat was grown in experimental plots of 10 m length and 1.8 m width.Spacing between rows was 40 cmand sowing rate was adjusted based on grain weight and germination of the various wheat varieties so as to attain a target plant population of 100 plants m-2.In both years,N rates of 0,50,100,200 or 300 kg ha-1were established by application of granular urea prior to sowing.The trial areas in both years deliberately followed a long fallow from a previous sorghum crop to ensure low starting soil N reserves.Soil N levels were measured to 1.2 m prior to sowing in each year with a total of 64 kg ha-1nitrate Navailable in 2006 and 42 kg ha-1nitrate N in 2007.
All plots were inoculated with Pst spores prior to a rain event during tillering in each season to supplement natural inoculation with wind-blown spores from neighbouring fields. Low-disease plots were then established in each trial by treatment of seed with fluquinconazole(Jockey-Bayer Crop Science at 450 mL 100 kg-1seed)prior to sowing and foliar applications of tebuconazole(Folicur-Bayer Crop Science at 290 mL ha-1)at the start of booting(GS32)and full flag leaf emergence(GS39).In 2006 the fungicide treatment was applied to all varieties,but in 2007 it was applied only to the susceptible variety H45 because Ellison was highly resistant to the dominant pathotype at the time of the trial.
The experimental design in 2006 was a split-plot design with fungicide treatment as the main plot factor,and variety and nitrogen as the subplot factors.In 2007 a randomised complete block design was used.There were four replicates in both years.
2.2.Disease assessment
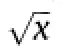
2.3.Yield and biomass parameters
In both years,a 1.5 m segment of each row was randomly cut at ground level from each plot just prior to harvest.These samples were used to determine biomass,after drying at 50°C for 48 h,and grain yield.Finalgrain yield was also obtained at maturity by harvesting each 10.0 m×1.8 m plot with a Kew experimental plot header.Grain protein concentration was determined by NIR reflectance.The trial was harvested 145 DAS in 2006 and 154 DAS in 2007.Data were analysed by ANOVA.
The amount of N harvested in the grain protein was calculated from yield and grain protein content,using a conversion factor of protein content of 5.61 times amino acid N content[8].N in protein was used rather than total grain N (which is about 1.05 times higher)because commercial prices are based on protein content.The Mitscherlich diminishing returns function,
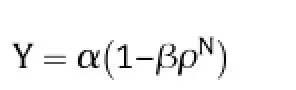
where Y represents grain protein N yield and N represents nitrogen application rate,was fitted to response curves for the susceptible varieties in each year using nonlinear regression in PASWStatistics version 18.This function was shown to give good fits to the response of yield and protein content of wheatin field trials from northern New South Wales[9].The parameters are interpreted as estimates of maximum yield (α),responsiveness to added N(β)and curvature of the response(ρ)[9].
3.Results
3.1.Rust infection
Stripe rust was the only foliar disease detected.No rust symptoms developed on the resistant variety Ellison in either year.In 2006 stripe rust severity at GS 75 was high in the susceptible variety HM,and was significantly(P<0.05) reduced by about half by fungicide treatment(Fig.1).Severity was very low in the moderately resistant variety Baxter. Nitrogen had a significant effect on rust severity,with severity increasing in both HM and Baxter as N rate increased(Fig.1).
Severity ofstripe rustwas also high inthe susceptible variety H45 in 2007(Fig.2).The fungicide treatment was more effective (P<0.0001)in reducing severity than in 2006.Although there was a trend for increased severity with increasing N,this was not significant(P=0.1).
3.2.Yield and biomass production in 2006
There were no significant effects of fungicide,variety or nitrogen on vegetative biomass in 2006.Mean biomass was 6.22 t ha-1.The effect on grain yield of the interaction between variety and N application rate was significant(P<0.05)in 2006. Grain yield was the highest in Ellison,and in HMwith fungicide treatment(Fig.3).Yield was reduced in HM without fungicide treatment,and was lowest in Baxter.Fungicide treatment had negligible effect on yield in Baxter.There was a highly significant(P<0.001)effect of N on yield,with yield increasing as N increased.There were no significant interactions between N and other factors.There was a highly significant(P<0.001) effect of variety on harvest index,with Baxter(0.39)having a lower harvest index than HM(0.43)or Ellison(0.44).The effects of other factors and interactions on harvest index were not significant.
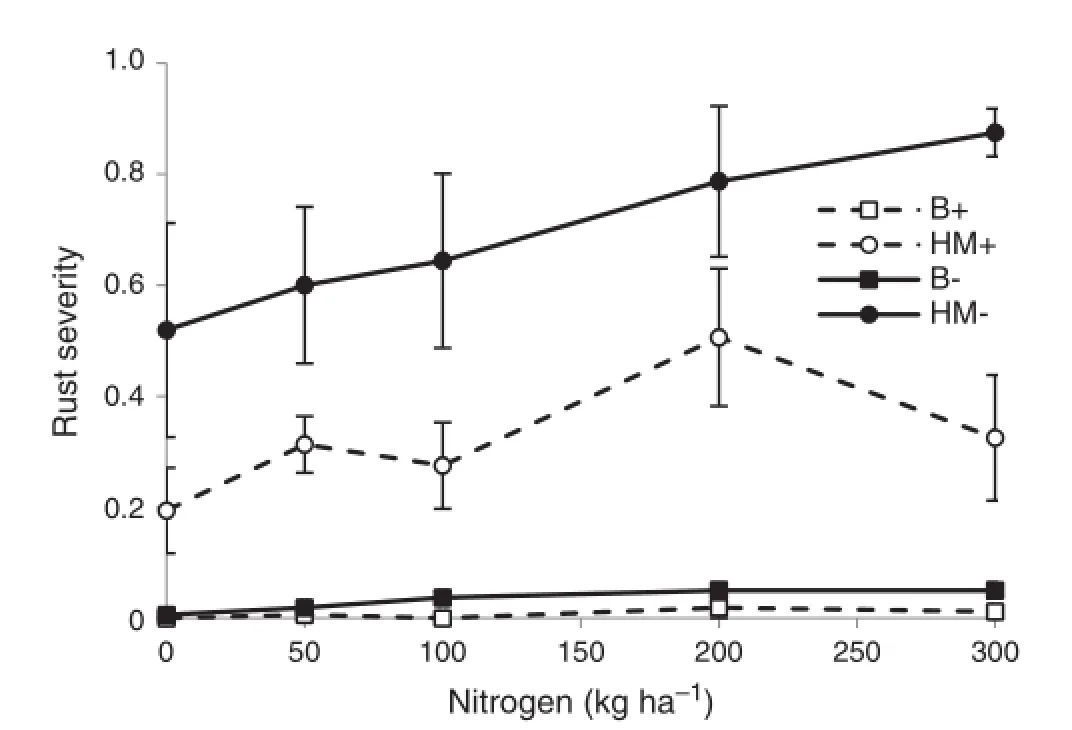
Fig.1-Severity of stripe rust in Baxter(B)and Hybrid Mercury (HM)wheat during grain filling in 2006 at different nitrogen levels with(+)and without(-)fungicide treatment.Error bars show±standard errors(n=4).
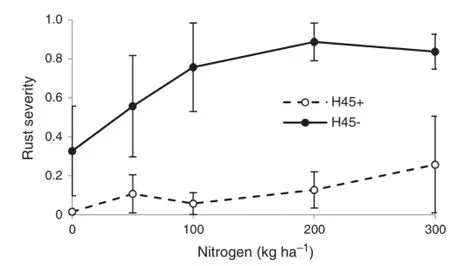
Fig.2-Severity ofstripe rust in H45 wheat during grain filling in 2007 at different nitrogen levels with(+)and without(-) fungicide treatment.Error bars show±standard errors(n=4).
3.3.Yield and biomass production in 2007
In 2007,vegetative biomass was significantly(P<0.001) higher in Ellison than in H45(Fig.4).Fungicide had no effect on biomass of H45.Increasing nitrogen significantly(P<0.01) increased biomass.
Yield of Ellison was higher than that of H45(Fig.5).Yield of H45 was significantly(P<0.001)increased by fungicide treatment.Nitrogen application significantly(P<0.001)increased yield.Harvest index was significantly(P<0.001)higher in H45 with fungicide(0.45)than in H45 without fungicide(0.41)or Ellison(0.41).There were no effects of N or interactions on harvest index.
3.4.Grain protein concentration
In 2006,there was a significant(P<0.001)effect of nitrogen on grain protein content(GPC).GPC increased with increasing N, but with little difference between 200 and 300 kg ha-1rates of N application(Fig.6).There was a significant(P<0.05)variety-by-fungicide interaction,with GPC in HM being increased from a mean of 11.2%to 11.7%by fungicide treatment.

Fig.3-Yield of Baxter(B),Ellison(E)and Hybrid Mercury(HM) wheat in 2006 at different nitrogen levels with(+)and without (-)fungicide treatment.Error bar shows pooled standard error (n=4).
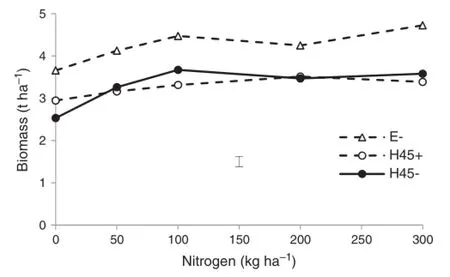
Fig.4-Vegetative biomass of Ellison(E)and H45 wheatin 2007 at different nitrogen levels with(+)and without(-)fungicide treatment.Error bar shows pooled standard error(n=4).
In 2007,fungicide had no significant effect on GPC in H45 (Fig.7).There was a significant interaction between N rate and variety,with the increase in GPC with N being slightly greater in Ellison than H45.
The effect of stripe rust on the ability of the plant to make use of added N was determined by calculating the amount of N in the grain protein per hectare for the susceptible variety in each year.The Mitscherlich(exponentialdiminishing returns) equation gave significant fits to the response of this parameter to N application rates(Table 1).In both years,fungicide treatment increased the predicted maximum grain N yield by 15-20%.In 2006,fungicide also increased the responsiveness of HM to added N.
The fitted curves were used to predict how much N would be returned in grain protein for each unit of N added as fertiliser(Fig.8).In 2006,the proportion of fertiliser N returned as grain N was much lower in the no-fungicide treatment at all levels of N.In 2007,there was no appreciable difference in N return between fungicide and no-fungicide treatments at low levels of N,with slightly higher N return in the fungicide treatment at N levels above 200 kg ha-1.
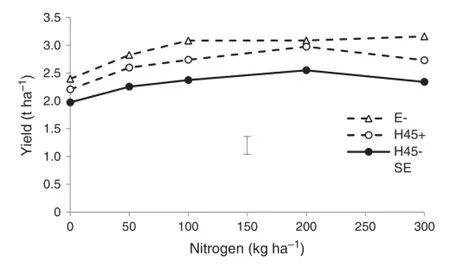
Fig.5-Yield of Ellison(E)and H45 wheat in 2007 at different nitrogen levels with(+)and without(-)fungicide treatment. Error bar shows pooled standard error(n=4).

Fig.6-Grain protein concentration of Baxter(B),Ellison(E)and Hybrid Mercury(HM)wheat in 2006 at differentnitrogen levels with(+)and without(-)fungicide treatment.Error bar shows pooled standard error(n=4).
4.Discussion
Only in the varieties HM and H45,which were susceptible to the dominant stripe rust pathotype present at the time of the field measurements,did fungicide treatment show a significant effect on any of the parameters measured.Stripe rust was also the only foliar disease seen in the plots.This observation indicates that reductions in biomass,yield and protein content in the no-fungicide treatments were due to the effect of stripe rust.Although fungicide treatment did not completely prevent rust infection,it afforded sufficient reduction in severity to discriminate the rust effect from variety and nitrogen effects.
Consistent with previous studies[1,2],increased rates of N increased the severity of stripe rust during grain filling.N application also increased yield and grain protein content in all varieties in both years,and generally there was no interaction between Nrate and disease.This finding suggests that stripe rust has the same effect on yield at all rates of N, even though rust severity increased as N rate increased.Thiscorrespondence may arise because higher levels of N lead to higher leaf area index(LAI[10]).Robert et al.[11]showed for leaf rust of wheat that photosynthesis in green parts of the leaf was unaffected by the presence of rust elsewhere in the leaf.It is possible that despite higher stripe rust severity at high N,with the higher LAIthe total amount of green leaf was not reduced.
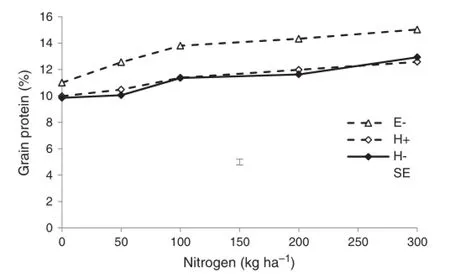
Fig.7-Grain protein content of Ellison(E)and H45 wheat in 2007 at different nitrogen levels with(+)and without(-) fungicide treatment.Error bar shows pooled standard error (n=4).
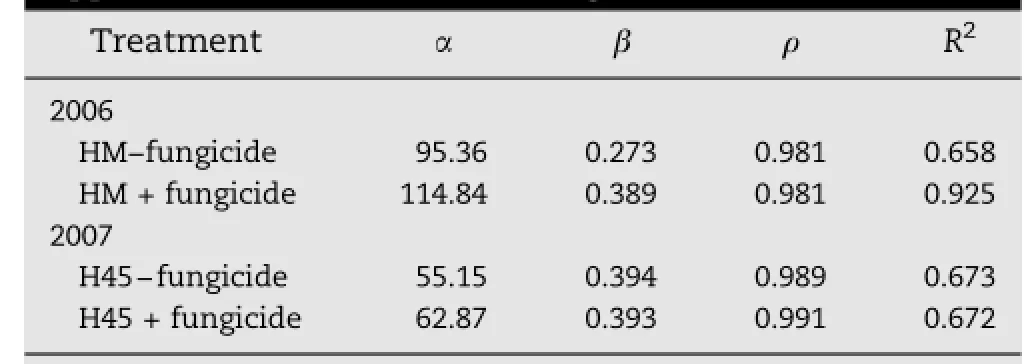
Table 1-Parameters of the Mitscherlich equation fitted to the response of grain protein N yield(kg ha-1)for wheat varieties Hybrid Mercury(HM)and H45 to nitrogen application with and without fungicide treatment.
Stripe rust reduced yield of the susceptible wheatvariety in both years,but it reduced grain protein content only in HMin 2006.This difference could be due either to environment,with yields in 2006 being almost twice as high as in 2007,or to genotype.
The effect of stripe rust on the proportion of added N recovered in the grain differed between the two years.In 2006, when both yield and GPC were reduced by disease,the rate of return on added N was approximately halved.This was a much larger effect than would be expected from a 10% reduction in yield and a reduction in mean grain protein from11.7%to 11.2%by the presence of stripe rust.However,in 2007,when yield was reduced by disease,protein content was unaffected.These conditions resulted in almost no difference in the marginal N yield in grain with the addition of varying N rates.
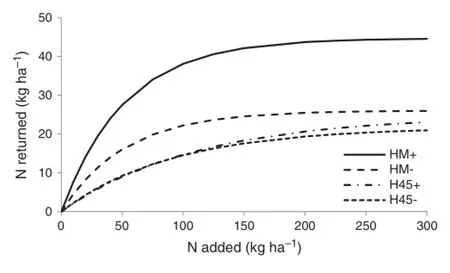
Fig.8-Modelled rate of N returned as grain protein relative to Nadded as fertiliser for Hybrid Mercury(HM)and H45 wheat with(+)and without(-)fungicide treatment.
The mechanisms by which rusts reduce N yield remain uncertain.Yield reductions are due to loss of photosynthetic area[11].Normally,reduced carbohydrate available for grain filling would be expected to increase relative protein content, as is typically seen when necrotrophic foliar diseases reduce yield[6].However,our experiments with stripe rust showed a reduction in yield accompanied by either no change or a reduction in protein content,indicating that the total amount of N entering the grain was reduced.There are three possible mechanisms for this effect.One is removalof Nfromthe plant tissue by the pathogen,principally as spores.Robert et al.[12] found that N content of leaf rust spores was lower,and C content higher,than those of wheat leaves,suggesting that rusts do not remove N from the plant at a higher rate than C. The other mechanisms are reduced uptake of N and reduced remobilisation from vegetative tissue into the grain after anthesis.Both uptake and remobilisation are reduced by late infections with foliar diseases[13].However,Bancal et al.[13] showed that variation in late N uptake had a greater effect on N yield than did variation in remobilisation in wheat crops affected by leaf rust and Septoria tritici blotch.
The effects of stripe rust on N yield found in this study were thus most likely due to reduced uptake of N during grain filling. The largest effects of stripe rust on N yield relative to N input were seen in 2006,which was the year with greater yield. Presumably the lower yields in 2007 reflected a reduction in assimilation after anthesis,accompanied by a reduced demand for post-anthesis N uptake.This hypothesis could account for differences in N use efficiency between seasons,although the possibility of genotype effects cannot be discounted.
Stripe rust clearly has the ability to affect the economics of N fertilisation,but such an effect was not consistent between the trials.The effects of genotype and environment on N use in the presence of rust should be explored further.
Acknowledgements
The authors gratefully acknowledge the receipt of postgraduate funding from the University of New England(UNE)and Cooperative Research Centre for Spatial Information(CRCSI), Australia.The CRCSI was established and supported under the Australian Governments Cooperative Research Centres Program.The authors also thank the NSW Department of Primary Industries,for the establishment of experimental plots at Breeza Research Station in NSW.
R E F E R E N C E S
[1]G.J.Ash,J.F.Brown,Effect ofnitrogen nutrition of the host on the epidemiology of Puccinia striiformis f.sp.tritici and crop yield in wheat,Australas.Plant Pathol.20(1991)108-114.
[2]D.L.Danial,J.E.Parlevliet,Effects of nitrogen fertilization on disease severity and infection type of yellow rust on wheat genotypes varying in quantitative resistance,J.Phytopathol. 143(1995)679-681.
[3]A.Darwinkel,Grain production of winter wheat in relation to nitrogen and diseases:I.Relationship between nitrogen dressing and yellow rust infection,Z.Acker.Pflanzenbau.149 (1980)299-308.
[4]R.J.Bryson,R.Sylvester-Bradley,R.K.S.Scott,N.D.Paveley, Reconciling the effects ofyellow rust on yield ofwinter wheat through measurements of green leaf area and radiation interception,Asp.Appl.Biol.42(1995)9-18.
[5]S.Neumann,N.D.Paveley,F.D.Beed,R.Sylvester-Bradley, Nitrogen per unit leaf area affects the upper asymptote of Pucciniastriiformis f.sp.tritici epidemics in winter wheat,Plant Pathol.53(2004)725-732.
[6]J.P.R.E.Dimmock,M.J.Gooding,The influence offoliardiseases, and their controlby fungicides,on the protein concentration in wheat grain:a review,J.Agric.Sci.138(2002)349-366.
[7]C.Wellings,H.Bariana,Assessment scale for recording stripe rust responses in field trials,Cereal Rust Report Season 2004, Plant Breeding Institute-Cereal Rust Laboratory,University of Sydney,2004.
[8]J.Mosse,Nitrogen to protein conversion factor for 10 cereals and 6 legumes or oilseeds-a reappraisalofits definition and determination-variation according to species and to seed protein-content,J.Agric.Food Chem.38(1990)18-24.
[9]I.C.R.Holford,A.D.Doyle,C.C.Leckie,Nitrogen response characteristics of wheat protein in relation to yield responses and their interactions with phosphorus,Aust.J.Agric.Res.43 (1992)969-986.
[10]I.Pearman,S.M.Thomas,G.N.Thorne,Effects of nitrogen fertilizer on growth and yield of spring wheat,Ann.Bot.41 (1977)93-108.
[11]C.Robert,M.-O.Bancal,B.Ney,C.Lannou,Wheat leaf photosynthesis loss due to leaf rust,with respect to lesion development and leaf nitrogen status,New Phytol.165(2005) 227-241.
[12]C.Robert,M.-O.Bancal,C.Lannou,Wheat leaf rust uredospore production and carbon and nitrogen export in relation to lesion size and density,Phytopathology 92 (2002)762-768.
[13]M.-O.Bancal,R.Roche,P.Bancal,Late foliar diseases in wheat crops decrease nitrogen yield through N uptake rather than through variations in N remobilization,Ann.Bot.102(2008) 579-590.
Yield
Grain protein
Nitrogen deficiency
杂志排行

The Crop Journal的其它文章
- Brief Guide for Authors
- Genome-wide analysis and identification of cytokinin oxidase/dehydrogenase(CKX)gene family in foxtail millet(Setaria italica)
- Optimal yield-related attributes of irrigated rice for high yield potentialbased on path analysis and stability analysis
- Effect of different levels of nitrogen deficiency on switchgrass seedling growth
- Evaluation of maize inbred lines currently used in Chinese breeding programs for resistance to six foliar diseases
- Effects of free-air CO2enrichment on adventitious root development of rice under low and normal soil nitrogen levels