梁子湖团头鲂的年龄和生长特征
2014-02-15宋文祝东梅王艺舟王卫民
宋文,祝东梅,王艺舟,王卫民
(华中农业大学 水产学院,湖北 武汉430070)
团头鲂Megalobrama amblycephala 隶属于鲤形目Cypriniformes、鲤科Cyprinidae、鲌亚科Culterinae、鲂属Megalobrama,俗称武昌鱼,是分布于长江中下游大中型湖泊的较大型经济鱼类[1]。中国对团头鲂基础生物学的研究开展得较早:易伯鲁[2]于1955年首次将团头鲂作为一个新种提出;1960年,曹文宣[3]对梁子湖团头鲂的基础生物学做了详细的论述;1990年,罗云林[4]整理了鲂属鱼类分类资料,肯定了团头鲂作为鲂属一种的分类地位;1992年,熊金林等[5]对低龄团头鲂鳞片上的环纹与生长特征进行了研究;1995年,谢从新等[6]对网湖的3种团头鲂年龄、生长进行了研究。
梁子湖位于长江中游南岸,湖区地处武汉、黄石、鄂州和咸宁四市的中间(114° 31' 19″~114°42'52″E,30°04'55″ ~30°20'26″N),流域面积为2083 km2,湖泊平均水深为2.54 m[7],是团头鲂重要的原种场之一。团头鲂是梁子湖的主要经济鱼类。鱼类的年龄与生长特性容易受气候、环境、水文、地质等因素的影响[8],近年来,梁子湖由于水利建设、生态资源过度开发,引起了其生态环境的变化,因此,亟需了解梁子湖现今团头鲂的种群数量和结构的变化。本研究中,作者对梁子湖团头鲂年龄和生长特征进行了初步研究,并根据其年龄和生长的相关资料综合评估了团头鲂种质资源状况,旨在为团头鲂资源保护及利用提供科学依据。
1 材料与方法
1.1 材料
2011年7月—2012年6月,每月于湖北省鄂州市梁子湖收集渔民采用刺网捕捞的野生团头鲂共310 尾。
1.2 方法
1.2.1 团头鲂生长指标的测量 对鱼体长、体质量和鳞径等生长参数进行测量,并以鳞片作为年龄鉴定材料。参考李文静等[9]的方法,取侧线以上2 ~3 行背鳍正下方形状规则、环片清晰的鳞片8 ~12 枚,用1 mol/L KOH 浸泡4 ~5 h,去除鳞片上附着的黏液和角质蛋白,再用清水漂洗2次,通风处晾干,然后置于两载玻片之中制成压片。在Olympus TH4-200 型解剖镜下读取年龄,用Qcapture Plus-In 成像系统拍照,测量鳞径R(R表示从鳞焦中心到前区与下侧或上侧交界边缘处的直线距离,此处年轮清晰,易于确定测量点)以及鳞焦到每个年轮处的轮径ri(依据鳞焦到年轮处的先后顺序依次记为r1,r2,…),精确到0.01 mm。
1.2.2 年龄的鉴定年龄组分组参照邓中粦等[10]的方法:0+~1 龄,记为1 龄鱼,1+~2 龄,记为2龄鱼,依次类推。
1.2.3 生长参数的计算

其中:X1和X2分别为年龄t1、t2时的体长L1、L2或体质量W1、W2。
1.3 数据处理
使用Microsoft Excel 2010 软件记录和处理数据,用Photoshop CS 软件进行图像处理,用SAS 9.1.3 统计软件分析数据。
2 结果与分析
2.1 鳞片的年轮特征
团头鲂年轮为疏密切割型,侧区切割明显,前区呈现疏密现象。后区和侧区交界呈现收缢现象,收缢程度与年龄呈现正相关(图1-A)。
年轮具有完整连续的特征,侧区轮纹结构最清晰,前区较清晰,后区环片破碎、特化成疣粒组织,并在密环处形成细密的环状脊,造成一定程度的变形,但就整体而言,在后区仍可见到年轮的延伸。年轮常出现环片破碎现象,偶见环纹断裂、融溶、聚合的现象。宽带与窄带交替出现,宽带所占比例较大,宽带和紧连的窄带构成1 个生长年带(图1-B)。
少部分个体在鳞焦附近呈现一部分环轮的幼轮现象,幼轮出现概率较小,且不明显,不呈现年轮的疏密切割特征,仅出现1 ~2 个环片缺失(图1-C)。多数3 龄以上团头鲂在侧区可见生殖轮,生殖痕位于正常年轮内侧密带上(图1-D)。
2.2 团头鲂的种群组成
团头鲂渔获物体长为(242.37±58.87)mm,体长分布范围为74.1 ~362.8 mm。其优势体长为180 ~320 mm,占群体总数的81.29%。体长为180 mm 以下时的个体数量较少,这是由于限制渔具网目对小规格鱼的捕捞所致。体长为310 mm 以上的个体数量迅速减少,体长为350 mm 的个体仅占样本总数的0.65%。
从图2可见:团头鲂体长出现较明显的1 个主峰(260 ~270 mm),主要为4 龄个体;出现5 个次峰(90 ~100,140 ~150,230 ~240,300 ~310,340 ~350 mm),主要为1、2、3、5、6 龄个体。
团头鲂渔获物由1 ~6 龄共6 个龄组组成,优势年龄组为3、4 龄,占总渔获物的55.48%,6 龄个体较少,仅占群体总量的4.84%(表1)。
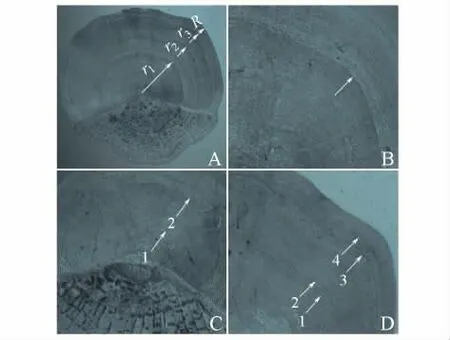
图1 梁子湖团头鲂的鳞片形态和年轮Fig.1 Form of scale and annual ring in bluntnose black bream Megalobrama amblycephala in Liangzi Lake
2.3 团头鲂的生长特征
2.3.1 鳞片生长特征
(1)体长与鳞径的关系。以团头鲂群体体长、鳞径作散点图(图3),选择相关系数最大的关系进行拟合。结果显示,线性拟合效果最佳,拟合方程为L=34.656R+25.21(n=310,R2=0.9621)。
(2)体长退算。团头鲂的体长与鳞径呈线性相关,且不通过原点,采用Rosa Lee 公式进行体长退算。结果显示,退算年龄越大,退算体长小于实际体长就越多,显现李氏现象[8](表2)。

图2 梁子湖团头鲂的体长分布Fig.2 Distribution of body length in bluntnose black bream Megalobrama amblycephala in Liangzi Lake

表1 梁子湖团头鲂不同年龄组的生长参数(平均值±标准差)Tab.1 Growth indices in bluntnose black bream Megalobrama amblycephala in different age classes from Liangzi Lake(珔x±S.D.)
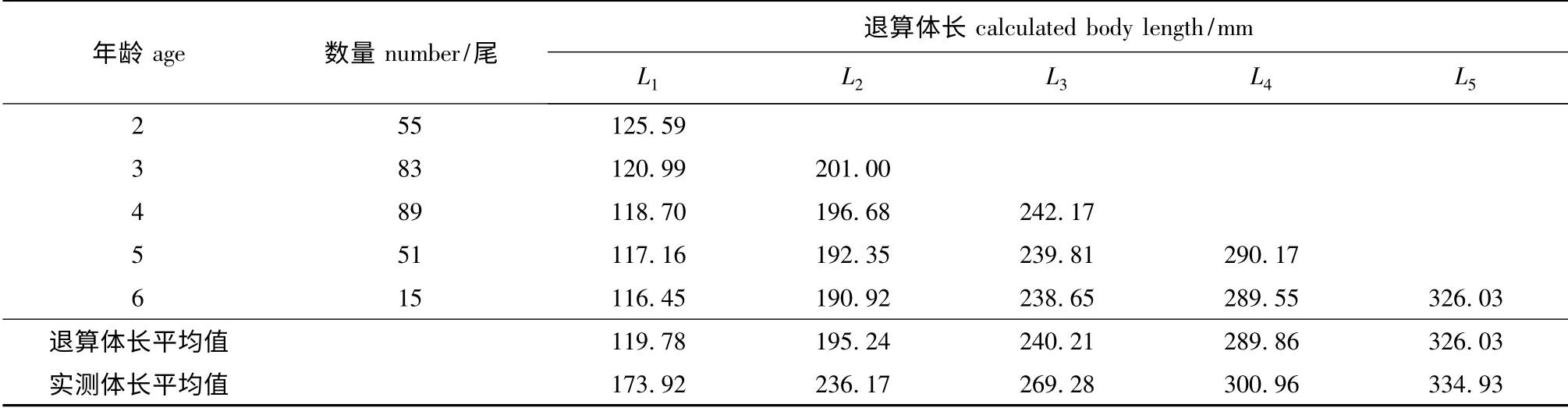
表2 梁子湖团头鲂的实测体长和退算体长Tab.2 The body length measured and calculated in bluntnose black bream Megalobrama amblycephala in Liangzi Lake

图3 梁子湖团头鲂鳞径与体长的关系Fig.3 Relationship between scale radius and body length in bluntnose black bream Megalobrama amblycephala in Liangzi Lake
2.3.2 体质量与体长的关系 根据团头鲂群体的体长(L)和体质量(W)作散点图(图4),对散点进行拟合,结果显示幂函数拟合效果最好,体质量与体长符合Taylor 公式,呈高度的幂相关,拟合方程为

关系式中幂指数b=3.1228,接近3,表明团头鲂群体体质量与体长的3次方呈线性关系,生长符合匀速生长型,满足Von Bertalanffy 方程的使用条件。

图4 梁子湖团头鲂体质量与体长的关系Fig.4 Relationship between body weight and body length in bluntnose black bream Megalobrama amblycephala in Liangzi Lake
2.3.3 生长方程和生长参数 使用Von Bertalanffy方程描述团头鲂的生长特性,体长(L)和体质量(W)的生长方程分别为

2.3.4 生长速度和加速度 为了研究团头鲂生长过程的变化特征,分别求出Von Bertalanffy[11-13]生长方程的一阶导数和二阶导数。
团头鲂体长和体质量的生长速度分别为

团头鲂体长和体质量的生长加速度分别为
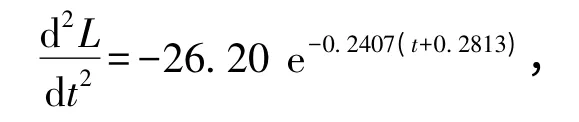

依据上述方程得到团头鲂体长与体质量的生长速度曲线和生长加速度曲线如图5所示。
从图5可见:团头鲂3 ~4 龄、5 ~6 龄时体长的生长速度略有减缓,未见拐点(图5-A),体长随年龄的增长逐渐趋近渐近体长;体长生长速度随年龄的增大不断减少(图5-C);而体长生长加速度却随着年龄的增大而逐渐升高,但d2L/dt2为负值,位于年龄轴下方(图5-E),表明团头鲂随着体长生长速度减缓,其递减速度逐渐趋缓。从图5还可见:团头鲂体质量的生长曲线随年龄的增长而增长(图5-B);体质量生长速度随着年龄的增加先增加后减少,呈现出现明显的拐点(图5-D);通过生长加速度方程,可以得出团头鲂生长拐点时间ti=ln b/K+t0=4.46龄,此时,Li=307.78 mm,Wi=607.70 g。当t<4.46 龄时,体质量生长速度不断增加,体质量生长加速度不断减少(图5-F),此时d2W/dt2为正值,位于年龄轴上方,表明4.46 龄以前是团头鲂体质量的增长阶段,尽管增长速度在减慢;在4.46 龄时,团头鲂的体质量生长速度达到最大值;当t>4.46 龄时,体质量的生长速度和生长加速度均呈现下降趋势,此时,d2W/dt2为负值,位于年龄轴下方,表明此时是团头鲂种群体质量生长递减阶段,且递减速度逐渐增加,约8 龄时,d2W/dt2降至最低点后又逐渐增加,表明随着体质量生长速度的进一步下降,递减速度也渐趋缓慢,团头鲂8 龄后开始进入衰老期。

图5 梁子湖团头鲂体长和体质量的生长、生长速度和生长加速度曲线Fig.5 The curves of growth,growth speed,and accelerated rate in bluntnose black bream Megalobrama amblycephala from Liangzi Lake
2.3.5 生长指数 利用各龄鱼的体长、体质量退算值,计算得到各项生长指标。从表3可见:随着年龄的增长,体长和体质量的年相对增长率、生长比速、生长指标呈现递减的趋势,表明团头鲂随着年龄的增加,生长逐渐趋于缓慢。

表3 团头鲂不同时期全长和体质量的生长指数Tab.3 Growth indices of total length and body weight in bluntnose black bream Megalobrama amblycephala
3 讨论
3.1 年龄、体长和体质量特征
鱼类年龄主要采用鳞片[14]、耳石[15]、鳃盖骨[16]、脊椎骨[17]、鳍条[18]等各种钙化组织进行鉴定。鲂属多采用鳞片作为年龄鉴定材料[5-6,19],本试验中,为了在获取团头鲂年龄材料后保持鱼体的完整性,选择鳞片作为年龄鉴定材料。团头鲂鳞片幼轮不明显,退算幼轮在体长50 ~60 mm 时出现,这是由于其繁殖从5月延续到8月,繁殖晚的个体由于适宜的生长季节短而形成。团头鲂3 龄以上通常可以观察到明显的生殖轮,表明3 龄以上个体已参与繁殖活动。
鱼类体长与年龄呈正相关关系,鱼类各年龄组众数不同,使得体长分布随年龄组呈现波峰。梁子湖团头鲂的体长分布呈现1 个主峰和5 个次峰的分布特征,各个波峰对应各年龄组的团头鲂。1956—1957年,梁子湖团头鲂2 ~7 龄的体长平均值分别为305.0、383.7、427.4、464.3、450.0、470.0 mm[3],明显大于本研究中团头鲂的平均体长。梁子湖湖泊总面积由20世纪50年代的520 km2缩减为至今的304 km2[20],1955—2001年45年间梁子湖的水生植被有很大变化,覆盖率在2001年时下降至54.27%;虽然水生植物总生物量呈现恢复,但仍然低于历史最高水平;水生沉水植被优势群落发生变化(以微齿眼子菜和金鱼藻为优势群落)[21],梁子湖的环境变化,可能是造成样本群体团头鲂体长明显逊于1956年群体的原因之一。
3.2 梁子湖团头鲂的生长特征
体长与体质量的关系是鱼类研究的重要指标,反映了鱼类生活的环境条件和生长状况[11]。本研究中,梁子湖团头鲂的b 值为3.1228,略小于1956年梁子湖团头鲂的b 值(3.191),明显大于网湖团头鲂的b 值(2.5451),但是两者均接近于3,表明团头鲂的体长、体高、体宽为等速生长。本研究中得出的梁子湖团头鲂的渐近体长为452.36 mm,渐近体质量为2034.61 g,明显低于网湖团头鲂(491 mm,2703 g)[6];生长方程中的K 值为0.2407,小于网湖群体(0.310);拐点年龄为4.46,大于网湖群体(2.88),生长呈现减缓的现象。推测原因:一方面是由于梁子湖与网湖的环境差异较大造成的;另一方面可能是由于环境竞争压力大,以及过高的捕捞压力引起的。因为鱼类在环境竞争压力大、高强度的捕捞选择下,通常种质资源会偏向小型化的选择。
本研究中,梁子湖团头鲂渔获物中3、4 龄较多,占渔获物总数的55.48%,据伍献文[22]报道,团头鲂初次性成熟年龄为3 龄,梁子湖野生团头鲂渔获物中性成熟个体占多数。团头鲂生长拐点年龄为4.46 龄,此时体长Li=307.78 mm,体质量Wi=607.70 g。分析显示,团头鲂5 龄前生长较为迅速,5 龄左右时较为合适的捕捞年龄。而渔获物群体优势年龄组为2 ~5 龄,占总渔获物的89.68%,最大年龄为6 龄,仅占4.84%,表明梁子湖团头鲂资源利用是不合理的,过度捕捞使种群结构遭到破坏,资源量日趋减少,种群出现了明显的小型化趋势,严重影响了团头鲂野生资源的可持续性。目前,应采取限制捕捞规格和捕捞渔具规格,以及在繁殖季节加强保护团头鲂等措施,以利于团头鲂资源的恢复和增殖。
[1]陈宜瑜.中国动物志:硬骨鱼纲鲤形目:中卷[M].北京:科学出版社,1998.
[2]易伯鲁.关于鲂鱼(平胸鳊)种类的新资料[J].水生生物学集刊,1955(2):115-122.
[3]曹文宣.梁子湖的团头鲂和三角鲂[J].水生生物学集刊,1960(2):57-82.
[4]罗云林.鲂属鱼类的分类整理[J].水生生物学报,1990,14(2):160-165.
[5]熊金林,杨干荣.低龄团头鲂鳞片上的环纹与生长特征的研究[J].淡水渔业,1992(6):20-22.
[6]谢从新,程文平,陈敬德.网湖三种鱼类的年龄与生长[J].水利渔业,1995(4):13-15.
[7]彭有轩,刘华,熊汉锋.梁子湖湿地保护与产业发展探析[J].湿地科学,2011,9(4):382-386.
[8]殷名称.鱼类生态学[M].北京:中国农业出版社,1995:21.
[9]李文静,王剑伟,谢从新,等.厚颌鲂的年龄结构及生长特性[J].中国水产科学,2007,14(2):215-222.
[10]邓中粦,余志堂,许蕴玕,等.汉江主要经济鱼类的年龄和生长[C]//中国鱼类学会.鱼类学论文集:第一辑.北京:科学出版社,1981:97-116.
[11]Ford E.An account of the herring investigations conducted at Plymouth during the years from 1924-1933[J].J Mar Biol Assoc U K,1933,19(1):305-384.
[12]Walford L A.A new graphic method of describing the growth of animals[J].Biol Bull,1946,90(2):141-147.
[13]Beverton R J H.Some observations on the principles of fishery regulation[J].J Cons Int Explor Met,1953,19:56-58.
[14]徐刚春,顾若波,闻海波,等.澄湖似刺鳊鮈的年龄和生长特征[J].中国水产科学,2009,16(3):307-315.
[15]陈新军,马金,刘必林,等.基于耳石微结构的西北太平洋柔鱼群体结构、年龄与生长的研究[J].水产学报,2011,35(8):1191-1198.
[16]吴立新,姜志强,秦克静,等.碧流河水库斑鳜年龄和生长的研究[J].大连水产学院学报,1996,11(2):30-38.
[17]李秀启,陈毅峰,李堃,等.抚仙湖外来黄颡鱼种群的年龄和生长特征[J].动物学报,2006,52(2):263-271.
[18]周成,朱国平,陈锦淘,等.印度洋南部大眼金枪鱼年龄鉴定及其与生长的关系[J].中国水产科学,2012,19(3):536-544.
[19]杨明生,王剑伟,李文静.厚颌鲂年龄材料的比较[J].动物学杂志,2004,39(2):58-61.
[20]彭有轩.湖北省湿地可持续发展研究[M].北京:中国科学技术出版社,2009:213-227.
[21]葛继稳,蔡庆华,李建军,等.梁子湖水生植被1955—2001年间的演替[J].北京林业大学学报,2004,26(1):14-20.
[22]伍献文.中国鲤科鱼类志:上卷[M].上海:上海科学技术出版社,1964.
