不同林龄马尾松(Pinus massoniana)人工林碳氮磷分配格局及化学计量特征
2014-02-08崔宁洁刘小兵张丹桔张健刘洋邓长春纪托未陈亚梅
崔宁洁,刘小兵,张丹桔,张健,2,刘洋*,邓长春,纪托未,陈亚梅
1. 四川农业大学生态林业研究所,四川 成都 611130;2. 生态林业工程重点实验室,四川 雅安 625014;3. 峨眉山市林业局,四川 峨眉山 614200
不同林龄马尾松(Pinus massoniana)人工林碳氮磷分配格局及化学计量特征
崔宁洁1,刘小兵3,张丹桔1,张健1,2,刘洋1*,邓长春1,纪托未1,陈亚梅1
1. 四川农业大学生态林业研究所,四川 成都 611130;2. 生态林业工程重点实验室,四川 雅安 625014;3. 峨眉山市林业局,四川 峨眉山 614200
为了解长江上游低山丘陵区马尾松(Pinus massoniana)人工林生态系统的C、N、P分配格局及化学计量特征,本文采用时空互代的方法,在宜宾高县来复林区选取三种不同林龄(5年生幼龄林、14年生中龄林、39年生成熟林),但立地条件相近、样地情况基本一致的马尾松(Pinus massoniana)人工林作为研究对象,对马尾松针叶、凋落物及土壤中的C、N、P含量及w(C):w(N):w(P)化学计量特征进行测定和分析。结果表明,(1)C、N、P含量均表现为针叶>凋落物>土壤,且在三个库之间差异显著;(2)林龄对针叶、凋落物、土壤的C、N、P及w(C):w(N)、w(C):w(P)计量比均有显著影响。(3)土壤C、N、P含量在成熟林中最高;针叶和凋落物的C含量在成熟林中最低,N、P含量则在中龄林中最高。(4)随林龄增加马尾松对N、P的利用效率降低,针叶、凋落物及土壤的w(C):w(N)与 w(C):w(P)均表现为下降。(5)马尾松针叶w(N):w(P)比值在14.37~15.53之间,说明该地区马尾松人工林受N和P的共同限制,但林龄对N、P养分限制的影响不显著。为提高该区马尾松人工林的生产力,建议在人工林的抚育管理中要适当增加N肥和P肥,同时也可在马尾松人工林引入豆科固氮植物以提高地力。该研究将马尾松针叶、凋落物及土壤结合起来探究随林龄增长C、N、P养分元素的分配格局及化学计量特征的变化,有助于全面、系统地揭示马尾松人工林生态系统的养分循环,对指导马尾松人工林生产,调节和改善林木生长环境,提高系统的养分利用效率及林地生产力具有重要意义。
马尾松人工林;林龄;化学计量特征
生态化学计量学是在生态学背景下,研究生物系统和生态过程中多种化学物质平衡问题的一门科学,主要强调活有机体主要组成元素(特别是C、N和P)的化学计量特征关系(Elser等,1996)。目前,化学计量学作为一种新的生态学研究工具已经被应用于从分子到种群、群落以及生态系统等各个层次(Zhang等,2003)。C、N和P元素对生物的生长、发育以及行为都起着非常重要的作用。C 是构成植物体内干物质的最主要元素;N作为关键元素,在氨基酸、蛋白质、核酸等物质的生物合成、提高植物的光合作用等方面起着重要作用;P是机体组织的基本元素,是核酸和酶的组成部分(李征等,2012)。植物组织中C、N、P化学计量比的变化可以揭示植物叶片养分浓度和养分限制性关系,以及不同植物对养分的需求和利用状况。w(C):w(N)和w(C):w(P)表述了C(生物量)与养分的比值关系,可以简单地理解为单位养分的生产力,即养分利用效率(Vitousek,1982)。w(C):w(N)和w(C):w(P)还反映了植物的生长速度,一般较低的w(C):w(N)和w(C):w(P)说明有机体具有较高的生长速率。叶片w(N):w(P)作为土壤环境限制性元素的判断标准,不仅是决定群落结构和功能的关键性指标,也可以作为对生产力起限制性作用的营养元素的指示剂(刘万德等,2010)。
目前,国外对于w(C):w(N):w(P)的化学计量学特征研究较多(Redfield,1958;Elser等,2000;McGroddy等,2004;Reich和Oleksyn,2004),国内相关研究起步虽晚但发展迅速,已对不同植被类型、不同空间尺度和不同演替阶段的植物叶片的化学计量特征先后进行了研究(Han等,2005;He等,2006;He等,2008;高三平等,2007;刘兴诏等,2010;吴统贵等,2010;阎恩荣等,2010;王晶苑等,2011),对土壤及凋落物养分含量的研究也较多(刘万德等,2010;王晶苑等,2011;杨会侠等,2010),但将这三者联系起来探讨这些化学计量特征变化的研究却很少。土壤作为地上植物营养元素的主要来源,其养分含量的变化及分布情况会对植物体的生长发育产生很大的影响(刘兴诏等,2010);叶片是地上植物进行光合作用,进行C同化、C积累以及贮存其他养分的重要器官;凋落物是森林生态系统树木与土壤的一个联结点,参与了系统能量与养分转移过程,在平衡与维持系统结构功能和长期生产力中发挥着重要的作用(杨会侠等,2010)。随林龄的增加,森林生态系统的组成、内部环境及土壤性质也会发生改变,进而影响养分的分配格局,将叶片、凋落物及土壤结合起来探究随林龄增长主要养分元素的分配及化学计量特征的变化将有助于全面、系统地揭示森林生态系统的养分循环及化学计量特征。
世界工业人工林正以200~300万hm2/a的速度发展,目前面积已超过5000万hm2(张丹桔等,2013)。马尾松(Pinus massoniana)具有耐干旱瘠薄、适应能力强等特点,是我国长江上游低山丘陵区退耕还林的主要人工林树种,在该区森林生态系统中占据重要地位。本研究以四川省宜宾低山丘陵区不同林龄的马尾松人工林为研究对象,对马尾松针叶、林下凋落物及土壤的C、N、P含量及w(C):w(N):w(P)化学计量比进行测定和分析,探讨随林龄增加马尾松人工林生态系统中养分元素的分配格局与物质循环特征,对指导马尾松人工林生产,调节和改善林木生长环境,提高系统的养分利用效率及林地生产力具有重要意义,同时可以为马尾松人工林的土壤肥力管理提供参考。

表1 林分基本概况Tab 1 The basic situation of sample plot
1 材料与方法
1.1 研究区概况
四川省宜宾市高县来复镇森林经营所林区,位于四川盆地南缘,高县北部,东经104°21′~104°48′、北纬28°11′~28°47′,地处川云公路中段,宜高路、来沙路在此交汇。气候类型属中亚热带湿润季风气候,多年平均温度17.5 ℃,年均降水量1070.4 mm,气候温和,雨量充沛。该研究样地选自于来复辖区内川云中路毛顛坳的人工马尾松林,海拔高度在400~550 m之间,土壤为铁铝土性质老冲积黄壤。选择地形地貌、海拔、母岩、土壤类型、坡度、坡位等立地条件相近的不同林龄的马尾松人工林为研究对象,5年生代表幼龄林,14年生代表中龄林,39年生代表成熟林(表1)。马尾松人工林乔木层以马尾松为主,林下层植物(乔木层林冠以下的灌木、草本以及蕨类植物)较密,主要灌木有枹栎(Quercus serrata)、梨叶悬钩子(Rubus pirifolius)、野桐(Mallotus japonicus)等;主要草本有芒(Miscanthus sinensis)、芒萁(Dicranopteris dichotoma)、荩草(Arthraxon hispidus)和商陆(Phytolacca Americana)等。
1.2 样品采集和分析
2011年8月,采用空间代替时间的方法,在宜宾市来复森林经营所毛颠坳林区选定立地条件相近,林龄分别为5a、14a和39a生的马尾松人工林,分别代表人工林演替的幼龄林阶段、中龄林阶段和成熟林阶段。每个年龄林分的马尾松人工林面积大约为3~8 hm2不等,在不同年龄的马尾松人工林内随机设置3个20 m×20 m的样方,在每个样方内挑选5株生长良好的健康马尾松树,采集新鲜的马尾松针叶,并在相应的马尾松林下选择1 m×1 m的5个小样方收集地表凋落物,将采集好的样品混合均匀后放入纸质档案袋中,做好标记带回实验室。经过105 ℃杀青、65 ℃烘干、机械磨碎后测定马尾松叶与凋落物的C、N、P含量。同时在采集凋落物的相应小样方下采集表层(0~20 cm)的土壤样品,5个样点的土样充分混合均匀后,取样带回实验室,土壤样品风干后,去除根、石头等杂物,研磨并过0.25 mm的网筛后测定C、N、P。该试验中的马尾松针叶、凋落物及土壤的C、N、P含量测定均统一参照《中华人民共和国林业行业标准LY/T-1999》。样品用硫酸-高氯酸消煮,有机质采用重铬酸钾氧化外加热法测定,全氮用凯式定氮法测定,全磷用钼锑抗比色法测定。
1.3 数据统计分析
采用EXCEL2003作图,SPSS19.0进行数据统计,分别对不同林龄的马尾松叶、凋落物及土壤C、N、P、w(C):w(N)、w(C):w(P)、w(N):w(P)进行单因素方差分析(Oneway ANOVA)和多重比较(LSD),并对马尾松针叶、凋落物、土壤的C、N、P含量,w(C):w(N)、w(C):w(P)、w(N):w(P)进行Pearson相关分析。
2 结果与分析
2.1 马尾松人工林针叶、凋落物、土壤的C、N、P含量
C含量表现为针叶>凋落物>土壤,针叶C的质量分数在470~526 g·kg-1之间,凋落物C的质量分数在388~453 g·kg-1之间,土壤C的质量分数则在13~34 g·kg-1之间(图1)。随林龄增加,马尾松针叶的C含量降低,不同林龄马尾松人工林针叶C含量差异显著(p<0.05),幼林和中龄林马尾松针叶的C含量差异不大,但均和成熟林有显著差异。凋落物C含量也随林龄增加而降低,不同林龄马尾松人工林林下凋落物C含量差异显著(p<0.05),多重比较显示,幼龄林和成熟林马尾松林下凋落物C含量存在显著差异。不同林龄马尾松人工林土壤C含量差异显著(p<0.05),有机质在成熟林阶段出现积累。
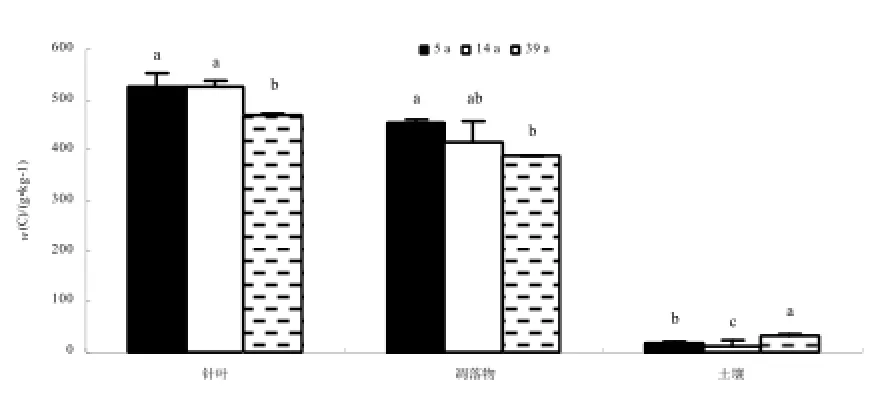
图1 不同林龄马尾松人工林针叶、凋落物、土壤C含量。不同小写字母代表不同林龄间的差异显著,下同Fig. 1 The C content of needle, litter, soil in different forest age masson pine plantation.The different lowercase letters stand for differences between the different forest age, the same below
N含量表现为针叶>凋落物>土壤,针叶N的质量分数在9.52~10.53 g·kg-1之间,凋落物N的质量分数在7.21~8.92 g·kg-1之间,土壤N的质量分数在0.63~1.58 g·kg-1之间(图2)。方差分析表明,林龄对马尾松针叶、凋落物及土壤N含量均有显著影响(p<0.05),马尾松针叶及凋落物的N含量均随林龄先增后降,土壤N含量则是先降后增,多重分析表明,幼龄马尾松针叶N含量分别与中龄林、成熟林具有显著差异,中龄马尾松凋落物N含量分别与幼龄林、成熟林有显著差异,成熟马尾松人工林林下土壤N含量显著高于幼龄林和中龄林。

图2 不同林龄马尾松人工林针叶、凋落物、土壤N含量Fig. 2 The N content of needle, litter, soil in different forest age masson pine plantation
P含量表现为针叶>凋落物>土壤,马尾松针叶P的质量分数在0.63~0.74 g·kg-1之间,凋落物P的质量分数在0.48~0.53 g·kg-1之间,土壤P的质量分数在0.23~0.43 g·kg-1之间(图3)。单因素方差分析显示,林龄对马尾松针叶P、凋落物P及土壤P含量均有显著影响(p<0.05)。多重比较显示,中龄马尾松针叶P含量分别与幼龄林、成熟林具有显著差异,中龄马尾松林下凋落物P含量与幼龄林具有显著差异,幼龄马尾松林下土壤P含量分别与中龄林、成熟林差异显著。

图3 不同林龄马尾松人工林针叶、凋落物、土壤P含量Fig. 3 The P content of needle, litter, soil in different forest age masson pine plantation
2.2 马尾松人工林针叶、凋落物、土壤的w(:C)w(N)、w:(C)w(P)和w:(N)w(P)
马尾松针叶的w(C):w(N)比值在45.07~55.22之间,凋落物w(C):w(N)比值在46.72~62.94之间,土壤w(C):w(N)比值在20.87~26.21之间(图4)。林龄对马尾松针叶、凋落物及土壤w(C):w(N)均有显著影响(p<0.05),马尾松针叶w(C):w(N)随林龄增加而下降,凋落物与土壤的变化一致,均是随林龄先下降后又有所回升。不同林龄的马尾松针叶w(C):w(N)均具有显著差异,幼龄马尾松人工林的林下凋落物及土壤w(C):w(N)均显著高于中龄林和成熟林的。
马尾松针叶的w(C):w(P)比值在699.82~830.37之间,凋落物w(C):w(P)比值在751.16~924.64之间,土壤w(C):w(P)比值在41.92~81.76之间(图5)。方差分析表明,林龄对马尾松针叶、凋落物及土壤的w(C):w(P)影响显著(p<0.05),马尾松针叶与林下凋落物w(C):w(P)均随林龄增加而下降,土壤w(C):w(P)则是中龄马尾松最低。

图4 不同林龄马尾松人工林针叶、凋落物、土壤w(C):w(N)比Fig. 4 The w(C):w(N) ratio of needle, litter, soil in different forest age masson pine plantation

图5 不同林龄马尾松人工林针叶、凋落物、土壤w(C):w(P)比Fig. 5 The w(C):w(P) ratio of needle, litter, soil in different forest age masson pine plantation
马尾松针叶w(N):w(P)比值在14.37~15.53之间,凋落物w(N):w(P)比值在14.72~16.68之间,土壤w(N):w(P)比值在2.11~3.75之间(图6)。从图中可以看出马尾松针叶w(N):w(P)随林龄增加先下降后上升,但是方差分析显示,林龄对马尾松针叶的w(N):w(P)影响不显著(p>0.05),对凋落物及土壤的w(N):w(P)影响显著(p<0.05)。多重比较显示,中龄马尾松林下凋落物的w(N):w(P)要显著高于幼龄和成熟林马尾松,土壤的w(N):w(P)则刚好与凋落物相反。
2.3 马尾松人工林针叶、凋落物、土壤C、N、P及其比值的相关性分析

图6 不同林龄马尾松人工林针叶、凋落物、土壤w(N):w(P)比Fig. 6 The w(N):w(P)ratio of needle, litter, soil in different forest age masson pine plantation
从相关分析结果可以看出(表2,表3),马尾松针叶P含量与凋落物P含量的相关性达到显著水平,针叶C含量与土壤C、N含量分别呈极显著和显著负相关,针叶N含量与土壤P含量显著正相关,土壤C含量与土壤N、P含量分别呈极显著和显著正相关。针叶w(C):w(N)与针叶w(C):w(P)存在极显著正相关,凋落物w(C):w(N)及凋落物w(N):w(P)存在显著正相关,针叶w(C):w(P)还与凋落物w(C):w(N)存在显著正相关,凋落物w(C):w(N)与土壤w(C):w(P)也存在显著正相关,凋落物w(N):w(P)与土壤w(C):w(N)、w(C):w(P)、w(N):w(P)均存在显著负相关。
3 讨论
(1)不同林龄马尾松林针叶、凋落物、土壤的C、N、P含量
研究表明,土壤C、N、P含量随林龄增加而得到积累,均在马尾松成熟林体现出最大值。据野外的多样性调查显示,随着马尾松人工林的演替进行,马尾松林下植物多样性增加,林内及土壤微环境发生变化。凋落物和根系分泌物是土壤有机质的最主要来源(Chapin III等,2011),土壤有机质是土壤肥力的物质基础,是各种营养元素特别是氮元素的主要来源,故土壤理化性质的变化、凋落物养分归还等因素均会影响土壤C、N、P含量。成熟马尾松人工林组成成分复杂且具有较明显的垂直结构,此阶段凋落物的主要组分不仅仅是马尾松枝叶,灌木、草本的枯落物在总的凋落物组分中的相对比例提高,阔叶树种及草本的凋落物能够在土壤动物与微生物的作用下较快分解并将养分释放到土壤中(李志安等,2004),Zhou等和Luyssaert等的研究也表明土壤有机碳积累与植被群落年龄存在正相关关系(Zhou等,2006;Luyssaert等,2008)。同时,成熟林的马尾松生长相对缓慢,干物质合成减少,对C、N、P的需求降低,土壤中C、N、P的积累量远大于消耗量,故C、N、P在成熟林时期得到积累。马尾松成熟林针叶中的C含量最低,推测与马尾松的生长规律有关。一般来说,马尾松在幼龄、中龄阶段生长较快,不断合成干物质,而到了成熟林阶段其生长逐渐减慢,故幼林及中龄林阶段马尾松C含量较高,成熟林时C含量下降。由于中龄林的马尾松生长速率较快,需要较多的rRNA以增加蛋白质的合成,最终导致叶片N含量升高。而rRNA又是植物的一个主要P库(刘万德等,2010;高三平等,2007),故马尾松针叶N、P含量均表现为中龄林最高。
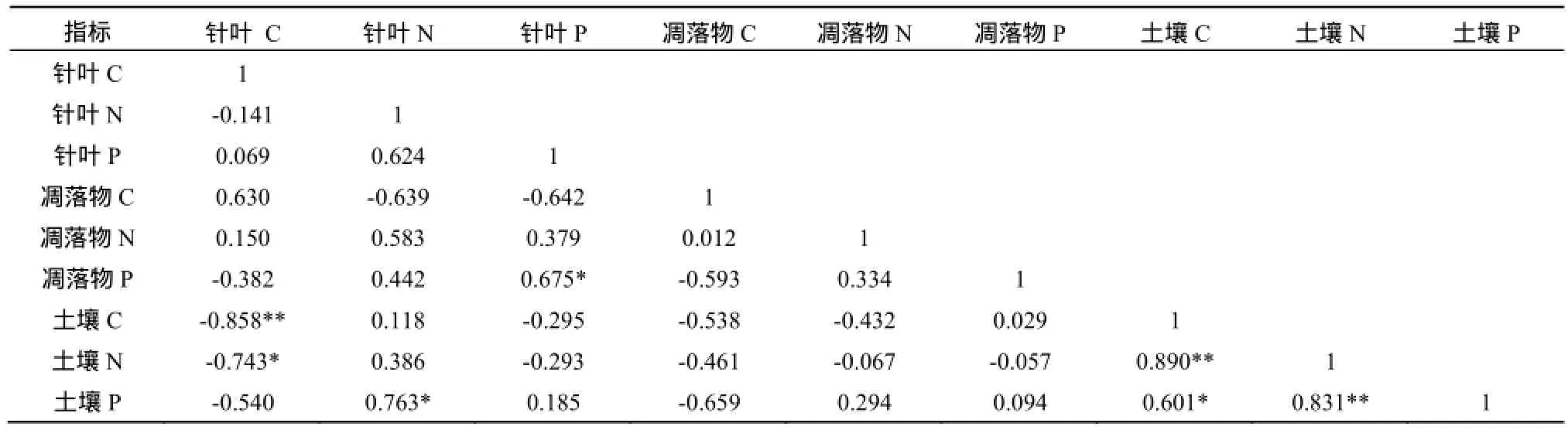
表2 C、N、P的相关性分析Tab 2 The correlation analysis between each element and the stoichiometric ratios of leaf

表3 化学计量比的相关性分析Tab 3 The correlation analysis between stoichiometric ratios
凋落物C含量则是马尾松幼龄林最高:一是和幼林马尾松针叶的C含量较高有关,因为马尾松针叶是凋落物组分的主要来源,相关分析结果也表明针叶C含量与凋落物C含量存在正相关关系;二是早期马尾松的凋落物组分比较单一,凋落物中含有较多的木质素及单宁,单宁会抑制微生物对凋落物的分解,降低凋落物的分解速率(Rothstein等,2004),土壤较低的养分含量也会影响凋落物的分解。有研究表明,土壤养分含量越低,凋落物的w(C):w(N)比越高,难分解的化合物越多,微生物数量及酶活性越低,凋落物的分解则越慢(郭剑芬等,2006)。凋落物N、P含量则是中龄林最高,这和此阶段马尾松针叶较高的N、P浓度相关。同时,14年生马尾松的林分密度较大,林下凋落物较多,较高的郁闭度使林下光照减少,光照不足也会降低凋落物的分解速度与养分释放(庞学勇等,2004;刘洋等,2006)。
(2)不同林龄马尾松林针叶、凋落物、土壤的w(C):w(N)、w(C):w(P)和w(N):w(P)
植物叶片的w(C):w(N)和w(C):w(P)表征植物吸收营养同化C的能力,一定程度上反映了单位养分供应量所能达到的生产力、植物对营养的利用效率以及植物的生长速度(刘万德等,2010;Wardle等,2004),因此具有重要的生态学意义。生长速率假说认为,生物体在生长发育过程中,能够通过改变它们的w(C):w(N):w(P)比值来适应自身生长速率的改变(Elser等,2003;Makino等,2003),生物个体的生长速率与体内的氮磷比、碳磷比具有负相关的关系(Elser等,1996)。本研究结果显示,马尾松针叶的w(C):w(N)比与w(C):w(P)比随林龄增加而降低。按照生长速率假说理论,该结果表明马尾松幼龄时期的生长速率低于中龄林和成熟林,而这和以往得出的马尾松的生长规律是不符的,可见生长速率假说的应用还有待进一步验证。这也可能是由于该地区马尾松生长发育的早期土壤相对贫瘠,可供植物吸收利用的N、P养分较少,使马尾松的生长发育受到一定的限制,造成马尾松w(C):w(N)比与w(C):w(P)比的失衡。
较高的w(C):w(N)与w(C):w(P)同时代表植物对N、P的利用率较高(Vitousek,1982),以往研究也证实了植物在营养元素供应缺乏的情况下往往具有较高的养分利用效率,是植物适应贫瘠养分状态的一种生存策略(Bowman,1994),这一观点恰好符合上文做出的推断。N和P作为植物生长的必需矿质营养元素和生态系统常见的限制性元素,在植物体内存在功能上的联系(Redfield,1958)。植物w(N):w(P)可用作N饱和以及P缺乏的诊断指标,不仅被用于确定养分限制的阈值,也可以用来反映土壤对植物生长的养分供应状况(Koerselman和Meuleman,1996)。以往研究认为,叶的w(N):w(P)<14反映植物受N限制,w(N):w(P)>16反映植物受P限制,14<w(N):w(P)<16表示受N、P的共同限制(Drenovsky和Richards,2004)。本研究结果显示,马尾松不同生长阶段叶的w(N):w(P)比值均处于14到16之间,说明马尾松生长受N、P的共同限制。马尾松幼、中龄林叶的w(N):w(P)比值接近14,说明受N的限制比较严重,到了成熟林阶段,w(N):w(P)有所上升,说明植物受N的限制情况有所缓解,而受P的限制加剧。在人工林的抚育管理中,特别是在马尾松幼林时期,建议要合理施用N、P肥以改善土壤养分供给状况。同时也可引入固氮植物以提高地力,促进马尾松人工林的生长发育。虽说不同生长阶段的马尾松针叶N、P含量均随林龄增加有显著变化,但w(N):w(P)却保持相对稳定,这说明植物自身具有调节元素需求与养分吸收平衡的能力,验证了植物的内稳态维持机制。
(3)马尾松林针叶、凋落物、土壤的C、N、P含量和w(C):w(N)、w(C):w(P)、w(N):w(P)的相关性
相关分析结果显示,马尾松针叶、凋落物、土壤的主要化学计量指标之间存在着紧密的相关关系,说明了生态系统内部C、N、P循环是在植物、凋落物和土壤三个库之间运输和转换的(Mcgroddy等,2004)。土壤是植物体生长所需营养元素的主要来源,从土壤中吸收的养分共同支持有机体的生长发育,植物通过叶片的光合作用源源不断地合成有机物质;植物在完成自身生活史后以凋落物的形式将养分归还于地面。凋落物经雨水淋溶、土壤动物破碎及微生物分解作用,一部分有机质损失,但是还有相当一部分的有机质及N、P等养分元素被释放到土壤中,故凋落物分解是土壤养分库的主要来源之一。这也导致了森林生态系统植物叶>凋落物>土壤的养分格局。
叶的w(C):w(N):w(P)的变化能影响到土壤养分关系,反过来土壤对植物的养分有效性也会产生反馈(Mcgroddy等,2004)。凋落物和针叶C、N、P含量及计量比多随林龄增加呈现相似的变化规律,说明了凋落物的养分承自植物叶,二者具有非常紧密的关联。凋落物中C、N、P含量明显低于马尾松叶,这反映了叶在衰落前后养分的再吸收状况,其实也是植物在养分供应有限的环境中进化出来的保持养分的一种机制(Drenovsky和Richards,2004)。植物、凋落物和土壤之间w(C):w(N):w(P)的差异意味着总生长效率的变化,其差异大小,代表了生产者及土壤微生物为维持生态平衡面临的养分竞争格局(Hobbie和Gough,2002)。目前,将植物、凋落物、土壤作为一个完整的系统来探讨C、N、P元素化学计量比在整个系统中的变化格局及其相互作用的研究还很少,其C、N、P平衡及养分转化的内在机制还需要进一步的探索。
4 结论
马尾松人工林C、N、P含量表现为针叶>凋落物>土壤,三个库之间差异显著,不同林龄马尾松针叶、林下凋落物、土壤C、N、P及w(C):w(N)、w(C):w(P)计量比均有显著差异,说明林龄影响着马尾松人工林生态系统的养分分配及化学计量特征。马尾松针叶的C含量在成熟林最低,N、P含量在中龄林最高。土壤C、N、P含量均在成熟林达到最高,说明随林龄增加马尾松人工林的土壤肥力得到较好改善。到成熟林阶段,马尾松针叶、凋落物及土壤的w(C):w(N)与w(C):w(P)有所下降,随林龄增加马尾松对N、P的利用效率降低。马尾松针叶w(N):w(P)比值在14.37~15.53之间,说明该地区马尾松人工林受N和P的共同限制。为了提高该区马尾松人工林的生产力,建议在人工林的抚育管理中适当增加N肥和P肥,同时也可在马尾松人工林内引入豆科固氮植物,以提高地力。马尾松针叶、凋落物、土壤的C、N、P含量及计量比存在着紧密的相关关系,说明该马尾松人工林生态系统的C、N、P元素在植物、凋落物和土壤三个库之间不断的运输转换,但其内在的维持机制需要进一步的深入研究。
BOWMAN W D. 1994. Accumulation and use of nitrogen and phosphorus following fertilization in two alpine tundra communities[J]. Oikos, 70(2): 261-270.
CHAPINIII F S, MASTONP A, MOONEY H A. 2011. Principles of terrestrial ecosystem ecology[M]. New York: Springer: 202.
DRENOVSKY R E, RICHARDS J H. 2004. Critical N:P values: predicting nutrient de ficiencies in desert shrublands[J]. Plant and Soil, 259(1-2),59-69.
ELSER J J, ACHARYA K, KYLE M, et al. 2003. Growth rate-stoichiometry couplings in diverse biota[J]. Ecology Letters, 6(10): 936-943.
ELSER J J, DOBBERFUHL D R, MACKAY N A, et al. 1996. Organism size, life history, and N:P stoichiometry[J]. BioScience, 46(9): 674-684.
ELSER J J, FAGAN W F, DENNO R F, et al. 2000. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature, 408(6812): 578-580. HAN W X, FANG J Y, GUO D L, et al. 2005. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 168(2): 377-385.
HE J S, FANG J Y, WANG Z H, et al. 2006. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J]. Oecologia, 149(1): 115-122.
HE J S, WANG L, FLYNN D F B, et al. 2008. Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes[J]. Oecologia, 155(2): 301-310.
HOBBIE S E, GOUGH L. 2002. Foliar and soil nutrients in tundra on glacial landscapes of contrasting ages in northern Alaska[J]. Oecologia, 131(3): 453-462.
KOERSELMAN W, MEULEMAN A F. 1996. The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology, 33(6): 1441-1450.
LUYSSAERT S, SCHULZE E D, BÖRNER A, et al. 2008. Old growth forests as global carbon sinks[J]. Nature, 455(7210): 213-215.
MAKINO W, COMER J B, STERNER R W, et al. 2003. Are bacteria more like animals than plants? Growth rate and resourse dependence of bacterial C:N:P stoichiometry[J]. Functional Ecology, 17(1): 121-130.
MCGRODDY M E, DAUFRESNE T, HEDIN L O. 2004. Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial redfield-type ratios[J]. Ecology, 85(9): 2390-2401.
REDFIELD A C. 1958. The biological control of chemical factors in the environment[J]. American Scientist, 46(3): 205-211.
REICH P B, OLEKSYN J. 2004. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences of the United States of America, 101(30): 11001-11006.
ROTHSTEIN D E, VITOUSEK P M, SIMMONS B L. 2004. An exotic tree alters decomposition and nutrient cycling in Hawaiian montane forest[J]. Ecosystem, 7(8): 805-814.
VITOUSEK P. 1982. Nutrient cycling and nutrient use efficiency[J]. American Naturalist, 119(4): 553-572.
WARDLE D A, WALKER L R, BARDGETT R D. 2004. Ecosystem properties and forest decline in contrasting long-term chronosequences[J]. Science, 305(5683): 509-513.
ZHANG L X, BAI Y F, HAN X G.. 2003. Application of N:P stoichiometry to ecology studies[J]. Acta Botanica Sinica, 45(9): 1009-1018.
ZHOU G Y, LIU S G, LI Z, et al. 2006. Old growth forests can accumulate carbon in soils[J]. Sciences, 314(5804): 1417.
高三平, 李俊祥, 徐明策, 等. 2007. 天童常绿阔叶林不同演替阶段常见种叶片 N、P 化学计量学特征[J]. 生态学报, 27( 3): 947-952.
郭剑芬, 杨玉盛, 陈光水, 等. 2006. 森林凋落物分解研究进展[J]. 林业科学, 42(4), 93-99.
李征, 韩琳, 刘玉虹, 等. 2012. 滨海盐地碱蓬不同生长阶段叶片C、N、P 化学计量特征植物[J]. 植物生态学报, 36(10): 1054-1061.
李志安, 邹碧, 丁永祯, 等. 2004. 森林凋落物分解重要影响因子及其研究进展[J]. 生态学杂志, 23(6): 77-83.
刘万德, 苏建荣, 李帅锋, 等. 2010. 云南普洱季风常绿阔叶林演替系列植物和土壤 C, N, P化学计量特征[J]. 生态学报, 30(23): 6581-6590.
刘兴诏, 周国逸, 张德强, 等. 2010. 南亚热带森林不同演替阶段植物与土壤中 N、P的化学计量学[J]. 植物生态学报, 34(1): 64-71.
刘洋, 张健, 冯茂松. 2006. 巨桉人工林凋落物数量、养分归还量及分解动态[J]. 林业科学, 42(7): 1-10.
庞学勇, 刘庆, 刘世全, 等. 2004. 川西亚高山云杉人工林土壤质量性状演变[J]. 生态学报, 24(2), 261-267.
王晶苑, 王绍强, 李纫兰, 等. 2011. 中国四种森林类型主要优势植物的C:N:P化学计量学特征[J]. 植物生态学报, 35(6): 587-595.
王绍强, 于贵瑞. 2010. 生态系统碳氮磷元素的生态化学计量学特征[J].生态学报, 28(8): 3937-3947.
吴统贵, 陈步峰, 肖以华, 等. 2010. 珠江三角洲3种典型森林类型乔木叶片生态化学计量学[J]. 植物生态学报, 34(1): 58-63.
阎恩荣, 王希华, 郭明, 等. 2010. 浙江天童山常绿阔叶林、常绿针叶林与落叶阔叶林的 C:N:P 化学计量特征[J]. 植物生态学报, 34(1): 48-57.
杨会侠, 汪思龙, 范冰, 等. 2010. 不同林龄马尾松人工林年凋落量与养分归还动态[J]. 生态学杂志, 29(12): 2334-2340.
张丹桔, 张健, 杨万勤, 等. 2013. 一个年龄序列巨桉人工林植物和土壤生物多样性[J]. 生态学报, 33(13), 3947-3962.
The Distribution pattern of carbon, nitrogen and phosphorus and the
stoichiometry characteristics of Pinus massoniana plantation in different ages
CUI Ningjie1, LIU Xiaobing3, ZHANG Danju1, ZHANG Jian1,2, LIU Yang1*, DENG Changchuan1, JI Tuowei1, CHEN Yamei1
1. Institute of Ecology & Forestry, Sichuan Agricultural University, Chengdu 611130, China 2. Key Laboratory of Ecological Forestry Engineering in Sichuan Province, Sichuan Agricultural University, Ya’an 625014, China 3. The Forestry Bureau of Emei, Emei 614200, China
In order to understand the distribution pattern of carbon、nitrogen and phosphorus and the stoichiometry characteristics ofpinus massoniana plantation ecosystem at hilly area in upper reaches of the Yangtze River, we used the space-temporal exchanging method, and selected three different forest stand age(5 -year-old young growth, 14 -year-old half-mature forest, 39 -year-old mature forest ) of masson pine plantation in similar site conditions as the objects, and analyzed the concentration of carbon, nitrogen and phosphorus and w(C):w(N):w(P) stoichiometry characteristics in the needle leaf, litter and soil. The results showed that the concentration of C, N and P were significant difference between needle leaf, litter and soil which in order of needle leaf > litter > soil; forest age had significant impact on C, N, P concentrations and w(C):w(N), w(C):w(P) stoichiometry ratios; The C, N, P content of soil were the highest in the mature forest, while the C content of needle leaf and litter were lowest in mature forest, and the N, P content of needle leaf and litter was highest in the mid-maturation forest; N and P use efficiency of masson pine plantation reduced, as w(C):w(N) and w(C):w(P) ratio of needle leaf, litter and soil declined with forest age increasing; The w(N):w(P) ratio of needle leaf was between 14.37 and 15.53, which indicated that the masson pine plantation was restricted by N and P together, but the influence of N and P limitation was not significantly by forest age. To increase the productivity of pinus massoniana plantation in the area, we should appropriately increase N and P fertilization in the plantation tending management. In addition, we also could interplant the leguminous nitrogen-fixing plants to improve soil fertility. This study combined masson pine needles, litter and soil to explore the distribution pattern of C, N and P nutrient elements and the change of chemical measurement characteristics with forest age increasing, will help to reveal the nutrient cycling of pinus massoniana plantation ecosystem roundly and systematically, it is very significant to guide the production of pinus massoniana plantation, adjust and improve the growth environment of masson pine, improve the nutrient use efficiency and the forest land productivity of the system.
Pinus massoniana plantation; forest age; stoichiometry characteristics
S713;S718.55
A
1674-5906(2014)02-0188-08
崔宁洁,刘小兵,张丹桔,张健,刘洋,邓长春,纪托未,陈亚梅. 不同林龄马尾松(Pinus massoniana)人工林碳氮磷分配格局及化学计量特征[J]. 生态环境学报, 2014, 23(2): 188-195.
CUI Ningjie, LIU Xiaobing, ZHANG Danju, ZHANG Jian, LIU Yang, DENG Changchuan, JI Tuowei, CHEN Yamei. The Distribution pattern of carbon, nitrogen and phosphorus and the stoichiometry characteristics of Pinus massoniana plantation in different ages [J]. Ecology and Environmental Sciences, 2014, 23(2): 188-195.
国家科技支撑计划课题(2011BAC09B05);国家自然科学基金(31370628);教育部博士点基金项目(20115103120003;20115103120002);四川省科技支撑计划项目(12ZC0017);四川省科技厅应用基础项目(2012JY0047);四川省教育厅重点项目(11ZA079);四川省教育厅科技创新团队资助计划项目(11TD006)
崔宁洁(1989年生),女,硕士研究生,主要研究方向为森林生态与人工林调控。E-mail:sicaucnj@163.com∗
2013-10-25
