艺术设计中创造性思维的fMRI研究:一项基于智能CAD的探索*
2014-02-02范亮艳范晓芳罗位超吴功航尹大志吕朱君明徐冬溶
范亮艳 范晓芳 罗位超 吴功航 严 序 尹大志吕 岳 朱君明 徐冬溶
(1华东师范大学认知神经科学研究所, 脑基因组重点实验室和磁共振重点实验室, 上海 200062)
(2华东师范大学信息学院 计算机系, 上海 200062) (3浙江大学 第二附属医院 神经外科, 杭州 310009)
(4 哥伦比亚大学医学院/纽约精神疾病研究所 磁共振中心, 纽约 10032)
1 引言
联想学派的Mednick (1962)在他著名的远距离联想理论中认为“在创造性过程中, 大脑中已经存在的思想以不同寻常的、创新的和有用的组合方式联系在一起。我们已有的每一个映像或概念与其它映像或概念联系在一起。”
人类之所以能够进行创造, 是因为事物之间能够基于一定的联系进行相互的结合与重组。一个事物变成另一个事物, 是因为结合了其它事物, 如果一个事物不与其它任何事物结合, 就永远不会发生变化, 创造也不可能发生。客观世界为人类提供了进行创造的前提和条件, 提取客观世界中的任何一个事物, 它的全部属性都可以归属到这个高维空间中的某些维度上, 而该属性所具备的值都可以定位到这一维度的某个点上, 正是这样的一些坐标点组成了人们所看到的具备不同属性不同特征的各种事物。这就为不同事物之间建立联系和进行组合提供了可能, 高维空间中的这些不同的维度就是不同事物之间进行彼此联系的中介。通过对处于不同维度上的事物的不同属性的值进行各种变化与重组,即可不断地生成新事物。创造性思维的本质就是客观世界的高维空间里, 人们以事物不同的属性所属的维度作为中介, 进行交叉、重组与综合, 进而产生新颖性的一种认识活动。
近年来, 随着功能磁共振技术等神经科学研究手段的不断发展与成熟, 针对创造性思维中发散性思维、顿悟这几个研究领域的神经机制的探索逐渐多了起来, 但是对于艺术设计及创作的研究不多。
Solso (2001)是较早采用神经成像手段研究艺术家创作过程的科学家之一。实验要求专业画家和普通被试创作一系列的脸部画像并记录其脑激活情况。结果发现, 无论是专业画家还是普通人, 在任务进行中右后侧顶叶脑区都呈现血流量的增加,而这个区域通常与面部识别处理有关。但也发现专业画家对应脑区的激活程度比普通人低。此外, 专业画家的右前侧脑区激活程度要高于普通人, Solso(2001)认为这是由于艺术家通常采用“高级指令”认知功能来观察和创作脸部画像的缘故。Bengtsson,Csíkszentmihályi和 Ullén (2007)通过即兴钢琴演奏任务考察了音乐创作中的脑活动。实验要求钢琴家在一段简单8拍旋律乐谱的基础上进行即兴变奏创作, 然后凭记忆对自己创作的旋律进行复弹。实验将即兴变奏、回忆复弹以及静息态分别作了对比,发现在包含创造过程的即兴演奏中, 激活脑区包括右背外侧前额叶, 前运动辅助区, 前运动皮层的背侧喙部以及颞上回的左后侧, 并且即兴变奏越为复杂, 前运动辅助区的激活程度也就越高。Bengtsson等 (2007)认为这些区域在音乐创作中构成网络相互联系, 彼此影响。同样采用了钢琴即兴演奏任务,Limb和Braun (2008)让6名爵士乐演奏家弹奏简单的普通旋律和复杂的爵士旋律, 然后在此基础上进行即兴创作。结果发现背外侧前额叶的广泛抑制,激活主要集中在内侧前额叶皮层, 这与之前Bengtsson等(2007)的研究结果存在一定出入。Limb和 Braun (2008)认为脑区呈现的这种激活-抑制模式反映了即兴创作中自我监控系统受到了抑制, 从而使得内部激发的行为得到了更为自然流畅地展现。此外, 在即兴演奏的过程中也观察到了执行音乐演奏行为的感觉运动区广泛的激活。Kowatari等人(2009)采用功能成像手段, 引入了物品设计的新颖范式来研究创造性思维。实验要求设计专业和其它专业的 40名被试进行钢笔设计。结果发现专业组激活的脑区更为有限, 相关专业训练会在艺术创作时对大脑左侧顶叶产生抑制, 这种抑制通过胼胝体等其它联结扩散到右顶叶和左侧前额叶, 大范围的抑制在右侧前额叶形成了一个优势中心。Kowatari等(2009)认为广泛的抑制实际上是对无关记忆信息干扰的抑制, 通过优势集中诱发对相关经验的提取、重构、整合, 完成艺术创作的整个过程。
Bechtereva等(2004)采用PET技术对被试的言语创造力进行了研究, 实验中给定被试 16个单词,要求被试将这16个单词按照自己的逻辑联系起来,编成一个故事。结果发现顶颞区与言语创造有密切关系。
近期Gilbert, Zamenopoulos, Alexiou和Johnson(2010)的一项研究把设计创造力的探讨引入到更为复杂综合的实际层面。实验通过调整指导语让 18名有相关设计经验的被试进行室内设计布置, 一种条件下有着对设计结果的诸多明确要求, 另一种条件下则更为开放自由。通过脑激活数据的比对, 观察到了自由设计与限定条件相比右背侧前额叶的显著激活, 但这种差异只在任务初期对任务的学习和领悟阶段出现, 在正式操作阶段未观察到这种差异。
综合各种研究, 发现前额叶对于艺术创作起到不可或缺的作用, 但艺术创作是通过前额叶与其它脑区的广泛联结协同作用来实现的, 后顶叶、顶颞区、边缘系统等可能也都参与进艺术创作的过程。
由于实验条件的限制, 在艺术创作活动时不方便进行脑成像扫描, 因此, 这类实验目前经常采取的形式是在整个实验过程中被试只需要想象自己将如何创作, 同时仪器对脑活动进行扫描和记录,待扫描结束后被试再将刚才想到的结果画下来或写出来。这种方式解决了脑成像实验仪器内无法进行创作活动的问题, 也排除了因为运动等不相关变量而可能产生的干扰。但是, 扫描过程所记录脑图像并非真正创作状态下的, 而且与实际创作过程的差异不得而知。其次这些实验任务都缺乏一个目标明确的模型, 只是一个传统的行为学实验套用磁共振影像学方法, 进行传统行为学方法研究的一个延伸。因此, 如果能在现有的理论基础上, 建立一个定量化可计算模型, 并在这个模型上开发一个与脑成像设备兼容的心理实验任务平台, 使得被试能够在扫描过程中完成创作任务, 并实时记录创作过程中的脑活动数据, 从而进行定量化的创造性思维研究。
人类的设计过程, 最集中地体现了人类的创造性能力。在计算机日益广泛应用的今天, 计算机辅助设计(CAD)系统被称为“设计师的助手”。但是大部分的 CAD系统, 仅仅只是人类创造工作的一个被动助手, 所有的创造性工作是人类设计师完成的。比如, Word处理软件只是替代纸张和笔墨, 但是写作的内容是人类完成的; Photoshop替代了颜料和画板并不能生成图片的内容。因此, 智能CAD就是要让 CAD系统具有真正的智能, 让计算机软件不仅仅是替代性工具, 而且是创造性工具。类比推理是这种创造性的方法之一。常规的类比推理是从已知的一个事物的因果推广到另外一个部分未知的对象上, 取得原先不知道的结论。类比学习和类比推理对人类历史的发展起到了重要作用, 将已有的知识扩展到多个未知领域, 并产生出知识。多源类比生成的概念和模型(Xu & Pan, 1995)和综合推理模型(潘云鹤, 1996)是对类比推理的推广并建立的一个可计算智能模型。在这个模型中, 首先需要从个体(源)提取多方面的属性, 这些属性构成了在高维空间中的单个属性维度。于是, 这个高维空间内或者外的任何一点, 都是这些属性的变化和组合所对应的一个崭新对象, 即潜在的创新。该模型正是利用这种原理在不同的个体间建立联系从而完成进一步的生成, 这反映了本研究对于创造性思维本质的推测。同时, 这个模型借助于抽象思维的符号推理方式, 由类比源定义出推理的基准空间,成功地将形象思维的因素转化到可计算的模型中去, 第一次实现了形象思维的可计算化。
泛化上述模型, 当这个空间维度很高, 超出人类想象能力的时候, 这样的生成物就是潜在的创造。潜在创造产品的数目呈组合爆炸, 但是并非每个潜在结果都是可用的, 因此人类世界的真实发明创造, 就是在潜在结果上加了一定筛选标准的结果。在这样的意义上, 高维空间内的有意义发现就意味着创造。因此, 多源类比推理模型, 可以说是一个对创造性过程的抽象和简化。但此类关于创造性思维的研究, 更多的是从人工智能及其在创造性设计中的应用的角度出发的, 并没有从基础神经科学角度去揭示人类大脑的创造性思维本质。
这个可计算的模型使得 CAD进行创造性设计成为可能。根据这个可计算的智能CAD理论模型,本研究开发了一个人脸脸谱设计的智能CAD系统,其中包括两种任务, 一种是无限制的自由人脸创作,另一种则是带有限制条件的设计(范晓芳, 2012)。同时开发和实现了一个磁共振兼容的心理学实验平台, 让被试参与进行创造性设计过程, 同时使用磁共振扫描仪对被试的脑功能图像数据进行同步采集。通过分离实验过程中与创造性相关的活动片断并关联与之对应的脑图像数据, 定位相关的激活或抑制脑区, 分析和建立创造性思维脑活动的神经回路模型。这样的尝试将是方法学上的一个创新, 可能为创造性思维的神经成像研究开辟一个新的领域和方向, 使得创造性思维的神经成像研究获得一个新的途径, 填补该领域研究的部分空白。
根据以往的研究, 自由创作设计与限定条件的设计相比(Limb & Braun, 2008; Gilbert et al., 2010),更加显著地激活了前额叶, 因此本研究假设:人脸设计中, 自由创作会更多地激活前额叶。此外, 先前研究也发现, 艺术创作中颞顶区, 边缘系统等脑区也会参与, 因此本研究假设:人脸设计自由创作中, 颞叶与创造性思维有着密切联系, 可能会不断产生新颖观点; 边缘系统可能会使被试产生强烈的创作欲望。
2 研究方法
2.1 实验设计
传统fMRI实验在IFIS (Invivo, Orlando, FL)等控制系统中运行 E-prime (http://www.pstnet.com/eprime.cfm)程序, 作为实验的刺激程序。但是E-prime无法满足复杂程序的编写要求。因此本研究用C语言编写了基于智能CAD的“三源类比人脸生成系统”, 并搭建了一个与磁共振仪器兼容的实验平台。三源类比人脸生成系统的开发环境是Microsoft Windows 7.0, 编译软件是 Visual studio 2008, 共分为两个部分。
第一部分:建立人脸图片数据库。本系统选用大量的真实照片, 在照片上对五官, 脸型等要素进行特征点标注, 进而生成一张二维线框图, 作为实验素材在实验中呈现给被试, 是生成结果的类比源。这部分工作由主试在实验前提前完成(图1)。
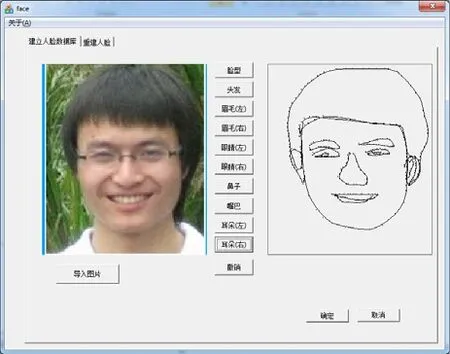
图1 三源类比人脸生成系统的人脸类比源采集功能模块注:导入人脸图片后, 选择要进行标注的部位, 然后在左侧图片上, 按顺时针或者逆时针一次点击该部位的轮廓,直至闭合, 此时右侧界面会同步显示点击到的特征点,并用平滑曲线将特征点连接起来, 所有部位标注完毕,即形成一张完整的人脸图片, 并保存, 供后续实验设计过程中作为类比源选取使用。
第二部分:三源类比人脸生成。选取图片库中的3张人脸图片, 以这 3张图片作为类比源, 这些类比源都具有不同的脸型, 长短不一的头发, 粗细不同的眉毛, 大小不一的眼睛, 高挺不同的鼻子, 厚薄大小均不同的嘴巴, 分别对构成人脸的五官等属性的参数进行调节和组合, 生成一张新的人脸(图2)。
通过改造, 成功搭建了适用于本实验的硬件实验平台(图 3), 使得刺激程序可以在扫描室外的一台笔记本电脑上运行, 信号同步到扫描仪内的显示器上, 呈现给被试, 被试的反应则通过自制的简易小键盘进行收集。整个过程主试可以通过扫描室外的电脑全程监控被试的操作过程, 行为数据也将被记录在这台电脑上。
以往对创造性思维神经机制的研究中, 基于任务模式的研究通常会采用将创造性任务与一般认知任务进行对比的范式。Gilbert等(2010)在 Goel和Grafman (2000)对脑损伤病人的研究基础上对范式提出了进一步的发展, 对同一任务的完成要求进行两种不同的变式, 分别成为“problem-solving task”和“ill-structure design task”。具体而言, 任务均为对一个房间进行布置,“problem-solving task”中设定了各种限制条件, 如“长的桌子要和大屏幕平行; 房间中入座的每个人要能看得到对方; 所有的家具都必须被使用上”等这样的指导语, 要求设计方案满足这些条件;“ill-structure task”则不进行具体限定, 只给定一些诸如“房间要宽敞; 房间可以用来集会、讨论”等模糊的指导语, 要求被试自由发挥制定设计方案。这种范式从更多的维度上保证了两种任务的同质性, 更加单纯地分离出了即兴创作和设计的成分。据此范式, 结合本研究的实验平台设计了两种不同的实验任务:

图2 三源类比人脸生成系统的人脸设计生成功能模块

图3 三源类比人脸生成系统的实验硬件平台
(1)基于 open-ends 的创造型人脸生成任务(Design Task):在创造型人脸生成任务中, 被试的任务即是根据界面中给出的三幅类比源图片, 完全根据自己的审美即兴发挥, 进行自由调节和组合,生成一张新的人脸图片。
(2)基于problem solving 的条件型人脸生成任务(Control Task):在条件型人脸生成任务中, 主试将事先使用多组图片进行调节、组合, 生成新图片,主试生成好的图片会在实验中呈现给被试, 被试的任务就是让自己生成的图片与给定的图片尽量匹配和吻合。任务界面见图4。
2.2 实验
2.2.1 实验材料
实验采集了86张2寸彩色正面证件照, 男女各半, 通过对这些照片进行特征点提取, 建立人脸图片库, 从中剔除五官相对位置偏差过大的图片, 最后保留了60张图片。使用这个图片库生成了45组design task和45组control task。召集41名被试参与了预实验, 通过测量其任务完成时间对任务难度进行匹配, 选定 12组 design task和 12组 control task。实验过程中各组任务的图片不会重复出现。

图4 三源类比人脸生成系统的任务界面
2.2.2 被试
23名(17女, 6男)在校大学生作为有偿被试参加了实验, 其中15人的数据最终有效。其余8组数据因为头动过大被剔除。被试均为右利手, 双眼视力或矫正视力正常, 无精神病或神经系统疾病史。所有被试均事先签署知情同意书。本研究得到了华东师范大学伦理委员会的批准。
2.2.3 fMRI实验要素
本研究采用的是组块式实验设计(block design)。刺激呈现流程如图5所示。通过预实验发现, 要创作出一幅自己满意的人脸, 被试至少需要120 s。为了让被试在给定任务下, 一直处于思考和创作状态, 每个任务的时间设定为120 s。

图5 刺激呈现流程
2.2.4 实验流程
第一阶段:任务学习阶段。
被试在正式扫描开始前将先在准备室里对实验任务进行学习。主试向被试讲解任务以及程序的操作方法等, 练习用程序与正式扫描所用程序是相同的,但图片不同, 任务数量不同。练习时图5的过程会重复2次, 每次之间会有30 s的休息时间。被试需分别熟悉这两种任务和整个实验的流程, 以确保在进入正式扫描之前已完全明确任务的内容。
第二阶段:正式扫描阶段。
被试在练习结束后将进入扫描室进行正式实验。正式实验时图5的过程会重复4次, 每次之间有30 s的休息时间。整个调节过程中的数据以及最后生成的图片都将被记录在扫描室外的电脑内。全程共35.5 min。
2.3 数据的收集与采集
2.3.1 数据采集
图像数据通过西门子 3.0T磁共振扫描仪采集获得。结构图像 3D-T1 MPRAGE序列, 主要参数如下, 层数:矢状面192层, 层厚:1 mm, 层间距:0.5 mm, 重复时间2530 ms, 回波时间:2.34 ms, 视野大小:256 mm × 256 mm, 采集矩阵:256×256, 采集时间:6.03 min。fMRI采用T2*加权的平面回波EPI序列, 主要参数如下, 层数:横断面 33层, 层厚:4 mm, 层间距:0.8 mm, 采集矩阵:64×64, 重复时间:2000 ms, 回波时间:30 mm, 倾倒角:90°,视野大小:230 mm × 230 mm, 采集时间:35.5 min。
2.3.2 数据分析和统计
对fMRI 数据的分析通过SPM8 (http://www.fil.ion.ucl.ac.uk/spm/)数据处理平台完成;
(1)预处理:基本步骤按先后顺序如下:(a)层间时间校正, 为了校正大脑不同层采集时间的延迟;(b)头动校正, 用来校正扫描过程中大脑的刚体运动, 任意方向平动大于 2.5 mm或者转动大于 2.5°被定义为头动过大; (c)空间标准化, 把头动校正后的功能像归一化到 Montreal Neurological Institute(MNI)标准空间; (d)空间平滑, 采用半高宽为6 mm的各项同性平滑核。
(2)统计分析:个体分析:首先根据实验设计和预处理后的 fMRI数据进行矩阵设计, 然后根据GLM 模型对此矩阵进行参数估计, 再通过设计对比度(contrast) (design task>control task)和 t-test得到相应的统计参数图(对比度图和 t值图), 最后对个体的t值图设置阈值(如p
<0.001, cluster size=10)从而得到在该对比条件下个体的激活图。组分析:利用每个个体的对比度图进行组分析, 采用的是双尾单样本t检验, 阈值为p
<0.01且激活团块大于40个体素(经过AlphaSim校正p
<0.05, 平滑核FWHM= 6 mm, 采用全脑mask), 其中AlphaSim校正是联合单体素的概率阈值(p
值)和最小团块(体素个数),采用蒙特卡罗(Monte Carlo)模拟的方法来确定联合阈值(http://afni.nimh.nih.gov/pub/dist/doc/manual/AlphaSim.pdf)。从而得到在该对比条件下的组激活图。3 结果分析
进行全脑分析发现, 与 control task相比,design task更显著地激活了如下脑区:内侧前额叶、右侧额上回、右侧颞上回、左侧前扣带回、双侧海马、左侧楔前叶、右侧尾状核等脑区(见表1, 图6);
4 讨论
通过这两种任务的比较, 本研究希望分离出创造性思维中与即兴创作相关的脑区, 从而探索与创造力相关的神经机制。
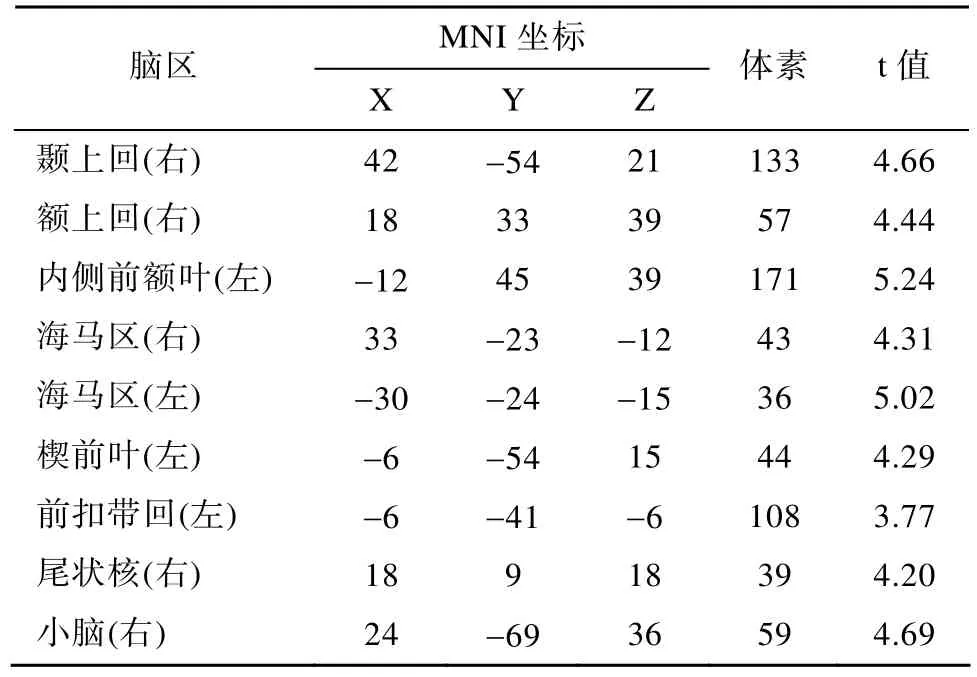
表1 Design Task>Control Task脑激活区

图6 design task与control task相比脑区激活图
研究结果显示, 与一般任务相比, 即兴发挥的创作任务更多地激活了包括内侧前额叶、颞上回、前扣带回、海马、楔前叶、尾状核等这些脑区, 提示创造性思维可能与额叶、颞叶、包括扣带回和海马等在内的边缘系统等多个脑区的活动有关, 这一结果与以往的研究报道相一致(Bhattacharya &Petche, 2005; Flaherty, 2005; Miller, 1998, 2004,2007)。本研究结果与Flaherty (2005)基于多例脑损伤病人研究报告以及部分神经影像学研究报告所提出的创造力三因素模型完全吻合。该模型认为创造力是由额叶、颞叶和边缘系统三个脑区所形成的网络彼此之间功能联结的结果。
4.1 内侧前额叶与即兴创作
本实验中使用即兴发挥的自由创作任务与明确了限定条件的任务相比, 分离出了所谓皮层中线结构(cortical midline structure, CMS)中的核心区域—— 内侧前额叶皮层(medial prefrontal cortex, 简称MPFC; BA9)的显著激活。这一结果与Limb和Braun (2008)针对音乐家所进行的爵士乐即兴创作的研究结果十分吻合。该研究使用音乐的即兴创作和过度学习两种任务, 同样分离出了内侧前额叶在即兴创作条件下的显著激活。本实验的研究结果在某种程度上支持了他们的推论, MPFC在即兴创作任务中表现为明显激活, 可能与即兴创作高度相关。
尽管与Limb和Braun (2008)的研究结果出现了如此惊人的相似, 但由于缺乏与更多成果的比较,对于 MPFC在即兴创作活动中的作用机制尚不能定论。但新近已经开始出现了关于默认模式网络(default mode network, DMN)与创造性思维关系的研究, 主要是来自日本的 Takeuchi等(2012)针对DMN与创造力之间的关系进行的研究, 研究并非基于任务模式, 而是直接测量了发散性思维能力的高低与DMN网络之间的关系, 结果显示可能存在高度相关。亦有一些其它有关创造性思维的研究证实 DMN可能在创造力中扮演重要角色(Fink,Grabner, et al., 2009; Fink, Graif, & Neubauer, 2009;Fink et al., 2010, 2012)。尽管目前对于DMN网络的功能尚无定论, 但有不少学者认为其与内省思维(introspective thought)、自我意识密切相关(Buckner,Andrews-Hanna, & Schacter, 2008; Gusnard,Akbudak, Shulman, & Raichle, 2001)。DMN与前文述及的皮层中线结构部分重合, 且 MPFC同样是DMN中的核心结构, 因此本研究认为MPFC与对内在信息的加工与表征有关, 在即兴创作活动中其作用表现为使得个体更多地倾向于对内在信息的提取与加工, 较少地依赖外界的刺激, 而较多依靠来自内部的进行自我表达的驱动, 是一种自上而下的加工过程。
4.2 颞叶与创造性思维
本研究还观察到了右侧颞叶在两种任务中激活模式的显著性差异。这与以往的研究认为颞叶与创造性思维尤其是艺术创造力有密切关系相一致(Flaherty, 2005; 沈汪兵, 刘昌, 王永娟, 2010)。
对于颞叶与创造性思维之间关系的研究, 最早始于一些神经系统疾病的病例报告。如梵高、陀思妥耶夫斯基都曾患有的“多写症” (Flaherty, 2005),Rankin等(2007)报告的语义痴呆症患者非言语创造力也受到了损害, 在艺术创造力上表现也很差,都提示颞叶可能与创造性思维有某种必然的内在联系。
后期的神经影像学研究报告中, 创造性思维的艺术创造力、顿悟、发散性思维等方面的相关研究都曾提到过颞叶的功能。Bhattacharya和 Petsche(2005)对画家和非画家进行的艺术画创作实验中发现了颞叶的激活; Fink, Graif和Neubauer (2009)针对舞蹈家进行的舞蹈创作研究中亦发现了颞叶的激活; 罗劲(Luo, 2004; Luo & Zhang, 2006; Luo &Niki, 2003; Luo, Niki, & Phillips, 2004; Mai, Luo,Wu, & Luo, 2004)等人的团队通过对谜语的研究提出了顿悟的神经机制模型, 其中包括了颞叶。Jung-Beeman等人(2004)通过对顿悟的研究发现,在问题的开始阶段和最后顿悟发生前的0.3 s, 在右侧颞叶都观察到了明显的激活。他们据此推测右侧颞叶负责在原本不相关的信息间建立联系。以发散性思维作为任务的研究中同样报告了颞叶与高发散性思维任务得分的相关(Fink, Grabner, et al.,2009; Chávez-Eakle, Graff-Guerrero, García-Reyna,Vaugier, & Cruz-Fuentes, 2007)。
因此本研究推测颞叶主要与创造性活动中在原本不相关的信息间建立联系, 不断地生成新的观点有关。以往研究证明, 颞叶中回涉及言语工作记忆, 任务切换, 想象和幻想(Bechtereva et al.,2004)。想象和幻想都是有利于大脑不断生成新观点的, 而任务切换则可能与顿悟中颞叶的作用有关。据此, 可推测颞叶提供了创造性思维过程中所必需的认知灵活性, 负责不断生成新颖性的观点并增强新颖性观点的品质。这与Flaherty (2005)所提出的创造力三因素解剖模型中对于颞叶在创造性思维中功能的表述也基本一致。
本研究中出现的颞叶在创作活动中的右脑单侧化优势, 这里套用沈汪兵等(2010)的“大脑两半球相互作用说” (Jung-Beeman, 2005)对其进行解释。“大脑两半球相互作用说”认为艺术创作力表现出更多的右侧优势可能是由于大脑右侧存在大面积的语义网络, 而左侧只存在一些零散的语义网络,同时该理论还认为右半球较左半球具有更高的互相连通性(interconnectivity), 可能也是右半球在创造力中发挥优势作用的原因之一。
4.3 边缘系统与创造性思维的关系
本研究数据结果还显示前扣带回、双侧海马、尾状核等边缘系统结构在两种任务中的差异表现,由此推测这可能与前人研究中提到的边缘系统在艺术创作中的动力驱动作用有关(Limb & Braun,2008; Flaherty, 2005)。Limb认为这其中的交互关系还有待进一步的研究, 并未做出明确解释。Flaherty(2005)则认为多巴胺通过边缘系统作用于大脑, 使大脑产生更强烈创作动机和欲望, 以维持创造性行为。据此, 本研究推测可能是在自由发挥的即兴创作任务中, 个体的创造动力驱动会表现得更强, 因此边缘系统表现出比普通任务更强的激活。
实验表明本研究开发的实验平台充分适用于功能磁共振成像实验及创造性活动神经机制的研究。使用该平台较好地克服了功能磁共振成像的实验条件限制, 该模型亦较好地模拟了创造和生成的过程, 预期以该模型为原理进行各种变式, 可以开展更多针对不同类型创造及设计活动的研究, 将针对创造性活动神经机制的研究拓展到更多的领域且能使实验任务具有更高的生态效度。
此外, 这也是首次将人类思维的神经生物学基础研究与人工智能的研究成果相结合的大胆尝试。类似创造性思维这样复杂高级的思维过程, 需要进行多学科跨领域的交叉研究, 才能取得更大的突破。当然这也同时有赖于更加先进的成像技术以及数据处理手段, 以期获得更为精准的数据结果进而逐步建立真实的人类神经网络模型。本文是进行本项目研究的第一个初步的探索性报告, 尚未提取关键认知过程相关的大脑激活信号。我们目前正在进行的第二步分析工作, 就是根据记录的被试反应如按键信息, 进一步细致分析和挖掘关键点再进行分析。无论如何, 希望这样的尝试能够是一个良好的开端, 为后面的更进一步工作走出开创性的一步。
5 结论
本研究在方法学上做出了创新和大胆尝试。通过将人工智能辅助设计研究成果—— 类比生成模型与创造性思维神经机制研究相结合, 开发出适用于磁共振仪器使用的“三源类比人脸生成”实验平台, 对即兴创作人脸生成和条件限定人脸生成两种任务引起的脑激活进行比较, 结果显示:与限定型生成任务相比, 即兴创作任务显著激活了包括内侧前额叶、颞上回、前扣带回、海马、楔前叶、尾状核这些脑区。其结果与“创造力三因素解剖模型”相吻合。内侧前额叶可能与即兴自由创作中更多的对自我信息的表征有关, 颞叶可能与不断产生和输出新颖性的观点有关, 边缘系统则可能主要与创造性活动中的动力驱动作用有关。总的来说, 创造性思维是多个脑区同时参与的高度分布式加工的结果。
Bechtereva, N. P., Korotkov, A. D., Pakhomov, S., Roudas, M.S., Starchenko, M. G., & Medvedev, S. V. (2004). PET study of brain maintenance of verbal creative activity.International Journal of Psychophysiology,53
(1), 11–20.Bengtsson, S. L., Csíkszentmihályi, M., & Ullén, F. (2007).Cortical regions involved in the generation of musical structures during improvisation in pianists.Journal of Cognitive Neuroscience,19
(5), 830–842.Bhattacharya, J., & Petsche, H. (2005). Drawing on mind's canvas: Differences in cortical integration patterns between artists and non-artists.Human Brain Mapping,26
(1), 1–14.Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L.(2008). The brain's default network: Anatomy, function,and relevance to disease.Annals of the New York Academy of Sciences,1124
(1), 1–38.Chávez-Eakle, R. A., Graff-Guerrero, A., García-Reyna, J. C.,Vaugier, V., & Cruz-Fuentes, C. (2007). Cerebral blood flow associated with creative performance: A comparative study.Neuroimage,38
(3), 519–528.Fan, X. F. (2012).FMRI study of human creative thinking --an exploration based on the intelligent CAD system.
Unpublished master’s thesis, East China Normal University.[范晓芳. (2012).人类创造性思维的fMRI研究--基于智能计算机辅助设计系统的探索.
硕士学位论文, 华东师范大大学]Fink, A., Grabner, R. H., Benedek, M., Reishofer, G.,Hauswirth, V., Fally, M.,... Neubauer, A. C. (2009). The creative brain: Investigation of brain activity during creative problem solving by means of EEG and fMRI.Human Brain Mapping,30
(3), 734–748.Fink, A., Grabner, R. H., Gebauer, D., Reishofer, G.,Koschutnig, K., & Ebner, F. (2010). Enhancing creativity by means of cognitive stimulation: Evidence from an fMRI study.Neuroimage, 52
(4), 1687–1695.Fink, A., Graif, B., & Neubauer, A. C. (2009). Brain correlates underlying creative thinking: EEG alpha activity in professional vs. novice dancers.NeuroImage,46
(3), 854–862.Fink, A., Koschutnig, K., Benedek, M., Reishofer, G.,Ischebeck, A., Weiss, E. M., & Ebner, F. (2012).Stimulating creativity via the exposure to other people's ideas.Human Brain Mapping,33
(11), 2603–2610.Flaherty, A. W. (2005). Frontotemporal and dopaminergic control of idea generation and creative drive.Journal of Comparative Neurology,493
(1), 147–153.Gilbert, S. J., Zamenopoulos, T., Alexiou, K., & Johnson, J. H.(2010). Involvement of right dorsolateral prefrontal cortex in ill-structured design cognition: An fMRI study.Brain Research, 1312
, 79–88.Goel, V., & Grafman, J. (2000). Role of the right prefrontal cortex in ill-structured planning.Cognitive Neuropsychology,17
(5),415–436.Gusnard, D. A., Akbudak, E., Shulman, G. L., & Raichle, M. E.(2001). Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function.Proceedings of the National Academy of Sciences of the United States of America,98
(7), 4259–4264.Jung-Beeman, M. (2005). Bilateral brain processes for comprehending natural language.Trends in Cognitive Sciences,9
(11), 512–518.Jung-Beeman, M., Bowden, E. M., Haberman, J., Frymiare, J.L., Arambel-Liu, S., Greenblatt, R.,... Kounios, J. (2004).Neural activity when people solve verbal problems with insight.PLoS Biology,2
(4), E97.Kowatari, Y., Lee, S. H., Yamamura, H., Nagamori, Y., Levy,P., Yamane, S., & Yamamoto, M. (2009). Neural networks involved in artistic creativity.Human Brain Mapping,30
(5),1678–1690.Limb, C. J., & Braun, A. R. (2008). Neural substrates of spontaneous musical performance: An fMRI study of jazz improvisation.PLoS One,3
(2), e1679.Luo, J. (2004). Neural correlates of insight.Acta Psychologica Sinica, 36
(2), 219–234.[罗劲. (2004). 顿悟的大脑机制.心理学报,36
(2), 219–234.]Luo, J., & Niki, K. (2003). Function of hippocampus in“insight” of problem solving.Hippocampus,13
(3),316–323.Luo, J., Niki, K., & Phillips, S. (2004). Neural correlates of the 'Aha! reaction'.NeuroReport,15
(13), 2013–2017.Luo, J., & Zhang, X. L. (2006). From the impasse to the breakthrough: The brain basis for insightful problem solving.Advances in Psychological Science, 14
(4),484–489.[罗劲, 张秀玲. (2006). 从困境到超越:顿悟的脑机制研究.心理科学进展,14
(4), 484–489.]Mai, X. Q., Luo, J., Wu, J. H., & Luo, Y. J. (2004).“Aha!”effects in a guessing riddle task: An event-related potential study.Human Brain Mapping,22
(4), 261–270.Mednick, S. A. (1962). The associative basis of the creative process.Psychological Review,69
(3), 220–232.Miller, B. L., Cummings, J., Mishkin, F., Boone, K., Prince, F.,Ponton, M., & Cotman, C. (1998). Emergence of artistic talent in frontotemporal dementia.Neurology,51
(4),978–982.Miller, B. L., & Hou, C. E. (2004). Portraits of artists:Emergence of visual creativity in dementia.Archives of Neurology,61
(6), 842–844.Pan, Y. H. (1996). The synthesis reasoning.Pattern Recognition and Artificial Intelligence,9
(3), 201–208.[潘云鹤 (1996). 综合推理的研究.模式识别与人工智能,9
(3), 201–208.]Rankin, K. P., Liu, A. A., Howard, S., Slama, H., Hou, C. E.,Shuster, K., & Miller, B. L. (2007). A case-controlled study of altered visual art production in Alzheimer's and FTLD.Cognitive and Behavioral Neurology: Official Journal of the Society for Behavioral and Cognitive Neurology, 20
(1),48–61.Shen, W. B., Liu, C., & Wang, Y. J. (2010).Neurophysiological basis of artistic creativity.Advances in Psychological Science, 18
(10), 1520–1528.[沈汪兵, 刘昌, 王永娟. (2010). 艺术创造力的脑神经生理基础.心理科学进展,18
(10), 1520–1528.]Solso, R. L. (2001). Brain activities in a skilled versus a novice artist: An fMRI study.Leonardo,34
(1), 31–34.Takeuchi, H., Taki, Y., Hashizume, H., Sassa, Y., Nagase, T.,Nouchi, R., & Kawashima, R. (2012). The association between resting functional connectivity and creativity.Cerebral Cortex,22
(12), 2921–2929.Xu, D. R., & Pan, Y. H. (1995). Generation-oriented analogy reasoning.Science in China,38
(9), 150–167.