猪对钙的消化和代谢
2014-01-22GonzlezVegaStein
■ J.C.González-Vega H.H.Stein
(伊利诺伊大学动物科学系,美国厄巴纳61801)
虽然在猪饲料中需要的矿物质的量很少,但无机元素是猪维持正常生长和繁殖必需的营养成分。动物体内矿物质含量约2%~5%,依品种不同而不同,而骨骼系统的矿物含量最大(Gillespie,1987)。钙(Ca)是体内含量最高的矿物质,其次是磷(P)(P;Kellems等,1998)。体内96%~99%的总Ca和60%~80%的总P是储存在骨骼组织中(Crenshaw,2001)。骨粉中含有36%~39%的Ca和17%~19%的P(Crenshaw,2001),同时在软组织如血液、体液和部分分泌物中也存在Ca和P,他们参与体内许多的生化反应(Gillespie,1987;Kel⁃lems等,1998)。
Ca和P对生理功能也发挥着重要作用,如肌肉收缩,神经冲动的传递,酶的活化,代谢反应,蛋白质的合成,维持渗透压和酸碱平衡,膜的组成和其他功能(Crenshaw,2001;Ewing等,2007)。
Ca和P属于常量矿物质,因为在日粮中的需要量水平超过100 mg/kg(Ewing等,2007)。然而,这两种矿物质的含量也应有控制,因为Ca和P之间存在互作,会影响两种矿物质的吸收(Crenshaw,2001;Ewing等,2007)。任一矿物质的过量或缺乏都会影响另一矿物质的利用。与动物蛋白(鱼粉、肉骨粉等)中的Ca含量相比,或和无机矿物质如石灰石、碳酸钙和磷酸钙等的Ca含量相比,谷物、油籽粕及许多其他植物性成分的Ca含量很低。
植物原料中的P生物利用率很低,因为这些原料中的部分P以和植酸结合的形式存在。在猪日粮中添加微生物植酸酶能释放部分和植酸结合的P,改善P的消化率并降低猪P的排放(Cromwell等,1995;Almeida等,2010;Kerr等,2010)。然而,猪日粮中的高Ca含量会降低植酸酶的功效,影响P的消化率(Lei等,1994;Lantzsch等,1995)。目前,关于调节Ca消化率和植酸酶对Ca消化率的作用机理方面的文献很少。
1 日粮中钙的来源
猪日粮中大部分Ca主要来自无机矿物质源,因为在谷物中Ca的含量很低。目前,关于有机钙和无机钙饲喂猪的消化率方面的文献资料很有限。在40 kg猪的典型玉米-豆粕型日粮中,来自玉米和豆粕的Ca约为1 g/kg日粮,而大约有5 g/kg日粮的Ca是来自石灰石和磷酸钙(National Research Council,1998)。
无机钙源的Ca含量在16.9%~38.5%(见表1),植物性饲料原料的Ca含量在0.01%~0.69%(见表2)。动物源性饲料原料的Ca含量比植物源性原料的Ca含量高,但一般低于无机钙源的Ca含量(表3)。石粉、碳酸钙和磷酸钙是猪日粮中最常见的Ca补充剂。如果以碳酸钙为标准,石灰石、文石、石膏、大理石粉和牡蛎壳中Ca的相对生物利用率在93%~102%之间,白云石质灰岩的相对生物利用率为51%~78%(见表4;Ross等,1984)。
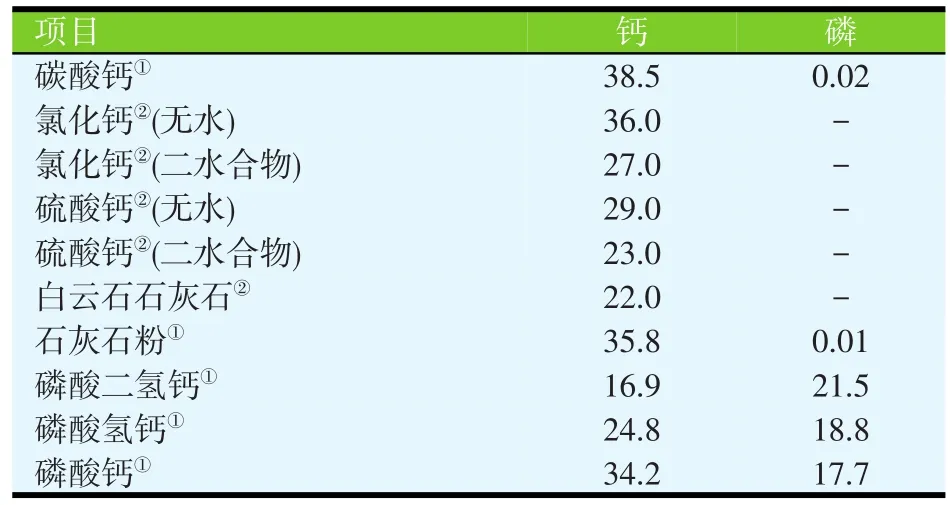
表1 无机钙源中的钙和磷的含量(%)
白云石质灰岩中Ca的生物利用率较低的原因可能是由于Mg的含量较高,从而降低了Ca的吸收,因为Ca和Mg共用同一转运机制(Ross等,1984)。这些原料的颗粒大小并不会影响Ca的相对生物利用率(Ross等,1984)。

表2 常见植物性原料的钙、磷和植酸含量(%)
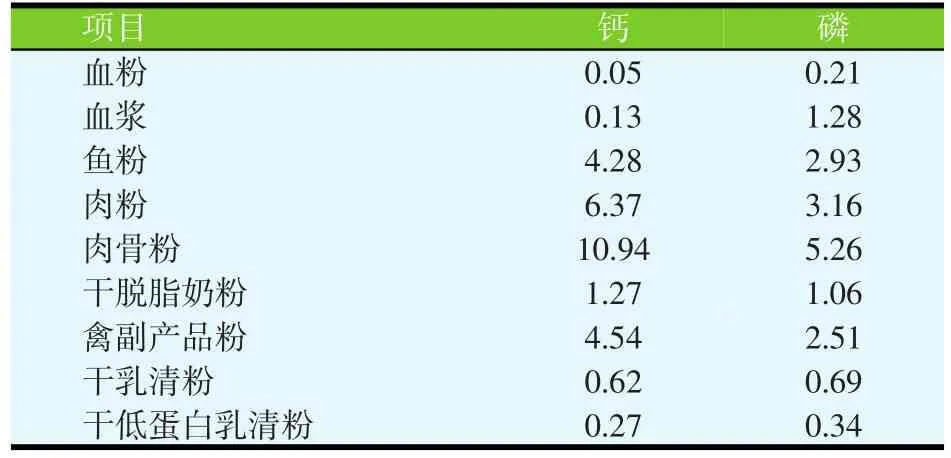
表3 动物源性饲料原料的钙和磷含量(%)
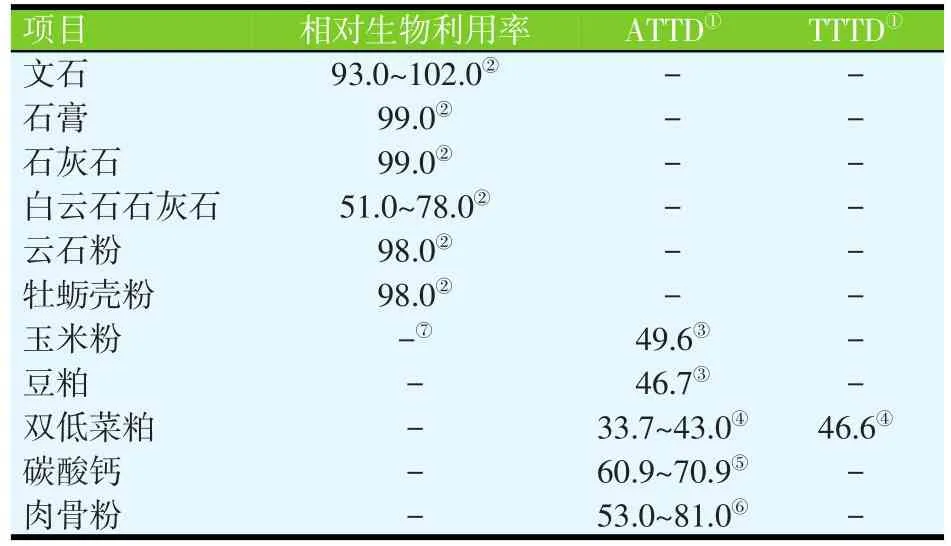
表4 钙的利用率和消化率(%)
相对生物利用率只是一个相对的衡量值,且并不反映Ca真正被动物体吸收的比例。动物体在不同生长阶段对Ca的吸收也不同(Hansard等,1961),但这不是绝对的(Baker,2011)。因此要测定营养物质的消化率,以更准确的估计营养物质饲喂给猪的利用率(Stein等,2007、2011)。测定营养物质消化率的好处是测定了营养物质被动物体吸收的比例以及被动物体排出体外的量,而不是相对生物利用率。
2 钙的消化率
营养物质的消化率是评估所有摄入养分是否被肠道全部吸收(Stein等,2007),是否所有被吸收的养分全部能够被动物利用(Ammerman,1995)。营养物质的消化率可定义为表观消化率、标准消化率和真消化率(Stein等,2007)。计算标准消化率或真消化率,需要先测定内源性损失量,计算表观消化率则不需要测定内源性损失量。有一部分内源性损失量是由于肠道内存在的干物质(DM)引起的,和日粮没有关系。这部分内源性损失被定义为基础内源性损失(Stein等,2007)。然而,另一部分的内源性损失是由于不同的日粮品种或饲料原料引起的,这部分损失则定义为特定内源性损失。
基础内源性损失和特定内源性损失两个构成了总内源性损失,并用于计算真消化率。然而,基础内源性损失往往比总内源性损失更容易测定,基础内源性损失则用于计算标准消化率(Stein等,2007;Almei⁃da等,2010)。表观消化率的缺点之一是,在测定混合日粮的消化率时,表观消化率不能叠加,而用标准消化率衡量混合型日粮的消化率时,则能叠加(Stein等,2005)。
消化率可表示为十二指肠、回肠或全肠道消化率。氨基酸(AA)的消化率一般用回肠消化率表示(Stein等,2007),磷(P)的消化率一般用回肠消化率或总肠道消化率表示,因为P在后肠道没有净吸收或排放(Bohlke等,2005)。然而,由于实际原因,很多原料通常都能测定其P的全肠道消化率及P的全肠道表观消化率(ATTD)和全肠道标准消化率(STTD)(Almeida等,2010、2012;National Research Council,2012;Rojas等,2012;Sulabo等,2013)。
由于唾液(Tryon等,1966)、细胞、胰腺和胆汁分泌引起的矿物质内源性损失会进入肠道(Vitti等,2010)。因此,用于测定猪的磷内源性损失的方法有很多种(Fernández,1995;Fan等,2001;Petersen等,2006;Bünzen等,2008)。关于牛的Ca内源性损失(Visek等,1953;Hansard等,1957;Martz等,1999)、鸡的Ca内源性损失(Cowieson等,2004;Liu等,2012)已有文献报道。猪的Ca全肠道内源性损失也已有文献报道(Besançon 等,1969;Fernández,1995;González-Vega等,2013),但关于猪的Ca基础内源性损失未见有报道。
P基础内源性损失是通过饲喂基础日粮测定的,在生长猪上已有报道P的基础内源性损失约为200 mg/kg干物质采食量(Petersen等,2006;Stein等,2006;Widmer等,2007;Almeida等,2010)。测定P或AA的基础内源性损失需要使用一种特定的日粮(无特定某种营养素)进行试验(Petersen等,2006;Stein等,2007),但这种方法并未用于猪的Ca基础内源性损失的估测。如果日粮营养素的摄入量和从粪便排出的营养素之间存在线性关系,那么营养素的总内源性损失则能通过y-轴截距外推法线性回归后得出,而营养素的真消化率即回归直线的斜率(Fan等,2001;Kil等,2010)。
某些饲料原料的磷STTD已有报道(Petersen等,2006;Stein等,2006;Widmer等,2007;Almeida等,2010、2012),但是关于Ca的STTD估测值则未见有报道。玉米、豆粕、双低菜粕、肉骨粉和碳酸钙的钙ATTD(Bohlke等,2005;Stein等,2011;González-Vega等,2013;Sulabo等,2013)及某些日量的钙ATTD(Kemme等,1997;Stein等,2006、2008;Malde等,2010)已有报道(见表4)。要将这些值转换成Ca的STTD或真实总肠道消化率(TTTD),则需要测定Ca基础内源性损失或总内源性损失。Ca总内源性损失最近已由González-Vega等(2013)报道,通过饲喂添加或未添加微生物植酸酶的双低菜粕日粮,采用回归程序测得,结果显示微生物植酸酶对Ca总内源性损失没有影响。不同方法测得的内源性损失值之间的差异比较已有报道(Dilger等,2006;Almeida等,2010),但关于Ca的内源性损失的报道很少。已有报道,用同位素稀释法测得的Ca总内源性损失为0.55 g/d(Fernández,1995)和1.49 g/d(Besançon等,1969),用回归程序测得的 Ca总内源性损失为 0.11~0.13 g/d(González-Vega等,2013)。
3 钙-植酸复合物
植物性原料中的大部分P是和植酸结合的,而日粮中的Ca也有1/3左右是和植酸结合形式存在的,这导致了Ca和P这两种矿物质的可利用率受限(Selle等,2009)。典型猪日粮中的植酸含量约为7~10 g/kg日粮(Selle等,2009;National Research Council,2012);而植酸显著影响了Ca和P的消化率。
日粮中过高含量的Ca对植酸酶的负作用效应(Lei等,1994;Lantzsch等,1995;Brady等,2002;Selle等,2009)可用3种机制来解释(Liu等,1998)。一种可能的机制是在小肠内形成了不可溶的Ca-植酸化合物(Wise,1983;Fisher,1992;Selle等,2009);另一可能的机制是高浓度Ca增加了肠道内容物的pH值,从而降低了植酸酶的功效(Sandberg等,1993;Selle等,2009);第三种可能的机制是,日粮中过剩的Ca可能会争夺植酸酶的活性位点,从而降低植酸酶水解植酸的功效(Wise,1983;Pointillart等,1985;Qian等,1996)。还需要更多的研究来阐明Ca、植酸和植酸酶之间的相互作用机制,以确定日粮中Ca的需要量。
4 钙的吸收
大部分Ca是在小肠中被吸收(Partridge,1978;Liu等,2000),但有些研究报道表明Ca也能在结肠被吸收(Liu等,2000)。关于Ca在小肠内被特定吸收的部位尚无明确定义。Ca在小肠近端(前4段)的吸收比在小肠远端的吸收要高(Moore等,1955a、b)。饲喂的日粮类型也可能影响Ca在肠道内被吸收的部位(Par⁃tridge,1978)。
在Ca被吸收之前,Ca和日粮中其他化合物结合存在,需要被释放成可溶性成分或以离子形式才能被吸收。胃酸和pH值依赖性的酶参与Ca的释放过程。断奶仔猪的十二指肠和空肠的pH值比回肠的pH值低,分别为5.7、5.9和6.9(Li等,2008)。钙结合蛋白负责Ca通过肠上皮细胞的转运(Bronner,1998),在十二指肠中钙结合蛋白含量比空肠的高(Allen,1982)。因此,十二指肠和空肠中pH值环境和钙结合蛋白的存在,可解释为什么Ca在小肠近端的吸收效率更高(Allen,1982)。
Ca在小肠内的吸收机制有不饱和细胞旁路(自由扩散)和饱和跨膜运输(主动运输)两种方式(Bron⁃ner,1987)。在小肠近端(十二指肠和空肠前段;Bron⁃ner,1987)主要是饱和跨膜运输机制。这种机制受日粮中Ca的含量影响,且当摄入的Ca含量比需要量低时这种吸收机制更活跃(Bronner,1987)。沿着小肠黏膜细胞之间的通道发生细胞旁路吸收,虽然不饱和旁路吸收机制和日粮类型不相关,但如果Ca的摄入量比需要量高很多,那么通过细胞旁路吸收机制运输的Ca会更多 (Bronner,1987)。
小肠内的饱和跨膜运输机制吸收Ca是通过如下3个步骤完成(Bronner,1998;R&D Systems Inc.,2007)。首先,肠腔内的Ca通过膜通道蛋白进入肠上皮细胞内,当肠腔内的Ca浓度低时,膜通道被打开,通道数量的增加依赖于维生素D的浓度。然后钙通过钙结合蛋白calbindins转运通过肠上皮细胞,钙结合蛋白calbindins是受维生素D调控的。Ca通过钙结合蛋白calbindins转运到Ca-Na泵,Ca-Na泵则将细胞外3个Na离子和1个Ca离子,或者1个Ca-ATP酶进行交换,把钙运输到细胞外。Ca-Na泵和Ca-ATP酶分布在肠上皮细胞的基底膜上,维生素D在通道蛋白的基因表达及Ca-ATP酶的表达中起重要作用(Bronner,1998;R&D Systems Inc.,2007)。
增加猪日粮中Ca的含量(Liu等,1998)及植酸酶的添加量(Lantzsch等,1995;Almeida等,2010;Poulsen等,2010)明显能增加Ca的吸收。相对于需求量增加Ca的水平并不影响Ca的ATTD(Stein等,2011),但当Ca的水平低于需要量时Ca的浓度会影响Ca的ATTD,因为钙的内源性损失代表了更多的Ca被排出体外(González-Vega等,2013)。草酸会和钙结合成草酸钙从而降低钙的吸收(Weaver等,1987)。硫酸盐也可能降低Ca的吸收,当日粮中含有DDGS等原料时考虑这一点是非常重要的,因为这些原料中含有较高的S(Pineda等,2008)。镁能直接降低Ca的吸收,因为Ca和Mg共用相同的运输机制,这也解释了为什么在白云质灰岩中Ca的生物效价较低的原因(Ross等,1984)。同样地,肉鸡日粮中Sr含量高也会降低Ca的吸收(Corradino等,1971)。鸡料(Migicovsky等,1951)或者鼠粮(Heggeness,1959)中添加抗生素也能提高Ca的吸收。同样的,某些纤维(Weaver等,2010)、AA(Wasserman等,1956)和益生菌也能增加Ca的吸收(Scholz-Ahrens等,2007)。
5 钙的代谢
通常日粮中有30%~70%的Ca被吸收,其中约65%~95%的Ca在猪体内蓄积(Kornegay,1985;Stein等,2011)。蓄积在体内的Ca主要用于维护骨架和骨骼发育。然而,还有少部分的Ca用于血液凝固、酶的活化、肌肉的收缩及其他功能(Soares,1995;Taylor等,2009)。
钙被吸收后,随体液在体内循环。大约有50%的Ca在血清中以Ca离子形式存在,45%的Ca是和白蛋白或球蛋白结合,剩下5%的Ca以其他形式存在(Bringhurst等,2006)。循环通过骨组织并沉积在骨组织中的Ca的比例取决于日粮中Ca的水平(Bronner等,1992)。同样地,肾循环中Ca要么继续参与循环,要么排出体外,肾循环中Ca的比例也受日粮中Ca和P水平的影响(Stein等,2006、2011;Vitti等,2010)。
6 骨骼
骨骼是由有机成分和无机成分组成。骨骼约有三分之一重量是由胶原蛋白和黏多糖组成。这部分成分对于维持骨骼结构非常重要,因为这些成分赋予了骨骼弹性(Frandson等,2009)。另三分之二的骨骼成分是由无机物质,如Ca和P盐组成(Frandson等,2009)。骨骼中Ca∶P比约为2.2∶1(Crenshaw,2001)。80%的这些无机物质是磷酸钙,剩下的主要以羟基磷灰石晶体[Ca10(PO4)6(OH)2]形式存在,剩下的20%主要是碳酸钙和磷酸镁(Frandson等,2009)。无机成分赋予了骨骼的硬度(Frandson等,2009)。钙、磷在骨骼中的沉积是相互依存的,这就意味着骨骼沉积过程需要这两个元素参与(Crenshaw,2001)。
骨骼组织是不断降解和合成的,由破骨细胞、成骨细胞和骨细胞参与这一过程。成骨细胞是参与骨骼组织的合成的骨细胞,破骨细胞是参与骨质再吸收的骨细胞,骨细胞则是由成骨细胞衍生的成熟的细胞(Frandson等,2009;Veum,2010)。骨细胞参与骨组织的沉积和骨吸收,也负责参与这些过程中的信号传导(Burger等,1995)。这些细胞的活动是受甲状旁腺激素(PTH)和降钙素调节的,而甲状旁腺激素(PTH)和降钙素则分别由甲状旁腺、甲状腺C细胞分泌,且取决于血浆中Ca的浓度(Frandson等,2009;Sherwood等,2013)。当血浆中Ca的浓度低于正常水平,则分泌PTH,PTH则通过增强破骨细胞的活性并抑制成骨细胞的活性,从而增加骨骼中Ca的再吸收。甲状旁腺激素也通过刺激钙循环和抑制磷酸循环在肾脏发挥作用。1α-羟基酶也被激活,而1α-羟基酶能激活维生素D成为1,25-二羟胆钙化醇,从而增加肠道中Ca的吸收(Costanzo,2006),因为1,25-二羟胆钙化醇可能会增强肠道内钙结合蛋白的合成(Kumar,1995),激活钙通道(Bouil⁃lon等,2003)。
如果血浆中Ca的浓度高于正常水平,则分泌降钙素,降钙素能抑制破骨细胞的活性,减少骨组织释放Ca(Costanzo,2006)。通过肾脏排放的Ca也增加,细胞摄取的Ca也增加(Crenshaw,2001)。
7 肾钙的代谢
肾脏在调节Ca浓度方面起到重要的作用,它可以通过过滤、再吸收和排泄血液中的Ca来维持血浆中Ca的浓度(Taylor等,2009)。Ca的吸收不受日粮中Ca和P的浓度的影响,但是,如果Ca的吸收超过需要量,超量吸收的Ca则通过尿液排出(Stein等,2006、2011)。饲喂无P日粮给猪时,Ca也能在小肠被吸收,但由于没有足够的P来保留被吸收进骨骼的Ca,因此,被吸收的Ca又通过尿液被排放(Stein等,2006)。
这表明,Ca的动态平衡不是在小肠内调节的;取而代之的是肾脏,肾脏似乎是主要的调节器官。钙磷比在Ca和P随尿液排放过程起到重要作用。当钙磷比高于骨组织合成的需要量时,则通过尿液排放的Ca增加,可用的有效P较低(Helander等,1996;Stein等,2006)。当日粮中钙磷比较低时,则通过尿液排放的P增加,用于合成骨组织的Ca较低(Stein等,2011)。虽然体内本身可以调节血浆中Ca的浓度,长期过量服用Ca或长期钙不足都会导致严重的问题,如肾结石或骨骼疾病(Taylor等,2009)。
血液中并非所有的Ca都能通过肾脏再循环,因为和白蛋白结合的Ca不能被肾小球过滤(R&D Sys⁃tems Inc.,2007)。如果Ca由肾小球过滤,大约有60%的Ca是在肾小管的近端被吸收,10%~15%是在亨利氏上升环被吸收,剩下的15%~25%是在肾远曲小管和肾集合管被吸收(Kumar,1995)。
8 结论
Ca是形成骨骼和牙齿所必需的,且是体内许多其他生理功能需要的。血浆中Ca浓度主要受PTH和降钙素的调节,PTH和降钙素在小肠、骨骼和肾脏内Ca代谢起到主要作用。Ca和P在体内的沉积,在骨细胞中这两种矿物质都需要,其中任何一种浓度相对较低都会抑制骨骼的合成,并导致另一种矿物质随尿液排出。由于大部分植物源性饲料原料的Ca含量较低,猪商品日粮中的Ca大部分是由石灰石、碳酸钙和磷酸钙提供的。
作为补充钙的钙源可以是有机物也可以是无机物,对于配方日粮而言,最重要的是要考虑这种钙源的Ca消化率。为能改善动物对Ca和P的利用率,还需要有更多关于猪日粮中各种原料的Ca的消化率方面的资料。因为,大多数植物性原料都含有植酸,植酸对Ca消化率的影响方面还需要进一步证实。然而,为了确定表观、标准或真消化率是否是最准确的,还需要测定猪胃肠道的Ca内源性损失量,配合饲料中各种原料成分的消化率值的可叠加性还有待证实。为确定回肠或全肠道消化率是否是描述Ca吸收情况最精确的指标,还需要描述清楚Ca的具体吸收位点。
(参考文献87篇,刊略,需者可函索)
