核桃青皮提取物对食管癌细胞增殖的抑制作用及其分子机制的探讨
2014-01-22张仲卫鄢文强李彩霞王冬滨李志刚司传领
张仲卫,鄢文强,李彩霞,王冬滨,李志刚,司传领,孔 棣
1.天津医科大学(天津300070)
2.天津市南开医院(天津300100)
3.天津科技大学(天津300222)
食管癌是一种常见肿瘤,治疗以手术为主,但复发转移率较高,化疗效果不理想。有研究表明,核桃青皮提取物可抑制体外培养的肿瘤细胞的增殖,其机理是直接杀伤肿瘤细胞[1]。本研究观察了核桃青皮粗提取物和核桃青皮提取物没食子酸、乙酸乙酯、紫杉叶素、7-D-芹菜糖-儿茶酚对食管癌细胞增殖和CyclinD1蛋白表达的影响,从而为进一步开发治疗食管癌的中药提取物提供理论根据。
1 材料与方法
1.1 细胞 食管癌细胞株EC9706,KYSE150(天津医科大学总医院肺癌研究所提供)。
1.2 主要试剂与仪器 二甲基亚枫(上海生工)、小牛血清(GIBCO公司)、RPMI-1640培养基(Hyclone公司)、胰蛋白酶(Hyclone公司),四甲基偶氮唑盐(MTT)(美国Sigma公司)。CO2培养箱(Thermo公司),倒置显微镜(Leica公司),KHB st-360免疫检测酶标仪(上海科华)。BCA蛋白定量试剂盒(北京康为世纪生物科技有限公司),鼠抗人β-actin多克隆抗体(武汉博士德生物工程有限公司),兔抗人cyclinD1多克隆抗体(cell signaling,USA),(HRP)标记的羊抗鼠Ig-G二抗(武汉博士德生物工程有限公司)。
1.3 方法
1.3.1 药物提取 核桃青皮用95%的乙醇萃取4次,风干得到微细粉末即为核桃青皮粗提物。将核桃青皮粗提物结合其水性残留物,在真空下浓缩,然后依次分馏,得到亚组分核桃青皮提取物没食子酸、乙酸乙酯、紫杉叶素、7-D-芹菜糖-儿茶酚[2]。
1.3.2 细胞培养 食管癌细胞株EC9706和KYSE 150分别常规培养于含10%小牛血清、0.1 mg/mL青霉素和0.1 mg/mL链霉素的RPMI1640培养液中,置37℃、5%CO2、饱和湿度培养箱中培养24 h,传代后取对数生长期细胞用于实验。
1.3.3 MTT法检测核桃青皮粗提物和核桃青皮提取物对食管癌细胞生长的抑制作用 将对数生长期食管癌细胞株EC9706和KYSE150分别用0.25%胰蛋白酶消化后,以每孔5×103个/mL细胞接种于96孔板中,每孔100 uL培养24 h后,分别换含不同浓度核桃青皮粗提物、没食子酸、乙酸乙酯、紫杉叶素和7-D-芹菜糖-儿茶酚的培养液,各组分的终浓度分别为10μg/mL,20μg/mL,40μg/mL,80μg/mL,160μg/mL,每组设6个复孔,同时设空白对照组。培养24 h,48 h,72 h后,弃上清,每孔加入5 mg/mL的MTT溶液10μL,继续孵育4 h,之后弃去上清液,每孔加入150μL的DMSO,置摇床上低速振荡10 min,使结晶物充分溶解,用酶标仪测定在490 nm波长处的吸光度(A)值。按下公式计算细胞增殖抑制率:细胞增殖抑制率=(对照组A值-实验组A值)/对照组A值×100%。
1.3.4 Western Blot检测食管癌细胞中CyclinD1蛋白表达水平 以1×105个/mL浓度将EC9706和KYSE150分别接种于6孔板中,每孔2 mL,待细胞贴壁后分别换含50μg/mL核桃青皮粗提物、没食子酸、乙酸乙酯的培养液,继续培养24 h后,弃培养液,用预冷的PBS洗涤细胞3次,每孔加100μL含PMSF的RIPA裂解液,快速刮下细胞,转移至1.5 mL离心管中,置冰上裂解30 min。于4℃,12 000 r/min离心10 min,取上清备用。BCA蛋白定量后,取35μg蛋白与上样缓冲液混匀,恒压下电泳,待溴酚蓝至分离胶底部时,停止电泳。半干转膜23 min。用5%脱脂牛奶室温封闭PVDF膜2 h。一抗(β-actin 1:200或CyclinD1 1∶1000,均用1×TBST稀释)4℃结合过夜,1×TBST室温洗膜;二抗(羊抗兔二抗工作浓度为1∶5000)室温孵育1.5 h,洗膜后加ECL避光孵育5 min,化学发光影像分析仪进行显影,采图,用Quantity one图像处理软件分析结果。
1.4 统计学方法 所有结果均以(x±s)表示,采用SPSS 16.0统计软件进行数据分析,多组均数间比较用方差分析,均以P<0.05为差异显著的检验标准。
2 结果
2.1 核桃青皮提取物对EC9706及KYSE150细胞增殖的抑制率 核桃青皮粗提物和4种提取物中的没食子酸和乙酸乙酯对EC9706和KYSE150细胞均有抑制作用,当浓度增加至80μg/mL时,对KYSE150的抑制率分别为82.75%、90.51%、88.36%,对EC9706的抑制率分别为85.54%、87.21%、85.17%。而紫杉叶素、7-D-芹菜糖-儿茶酚对细胞增殖无影响。如图1、图2所示。
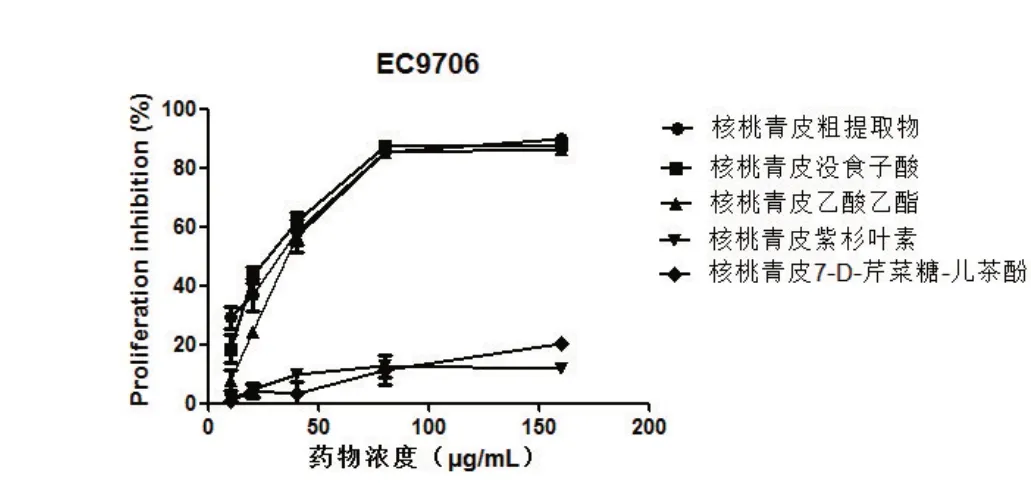
图1 不同浓度核桃青皮提取物对EC9706细胞增殖的抑制率
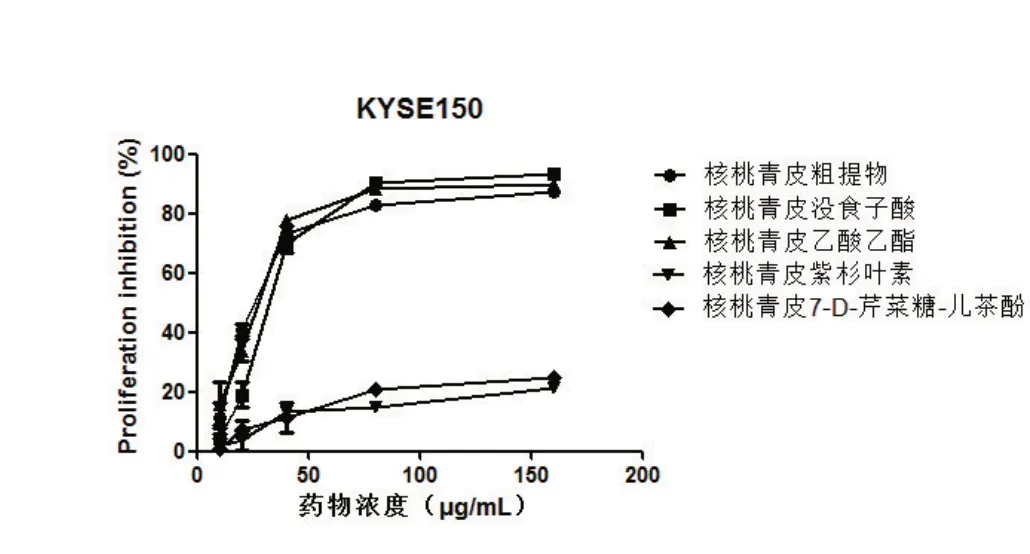
图2 不同浓度核桃青皮提取物对KYSE150细胞增殖的抑制率
2.2 核桃青皮粗提物和两种有效核桃青皮提取物对EC9706和KYSE150细胞增殖的抑制作用 核桃青皮粗提物、没食子酸和乙酸乙酯浓度为10~160μg/mL时均能抑制食管癌细胞株EC9706和KYSE150的增殖,高浓度核桃青皮提取物对食管癌细胞生长的抑制作用更强。结果见表1,表2及表3。
表1 核桃青皮粗提物对EC9706和KYSE150细胞增殖的抑制作用(±s,n=5)
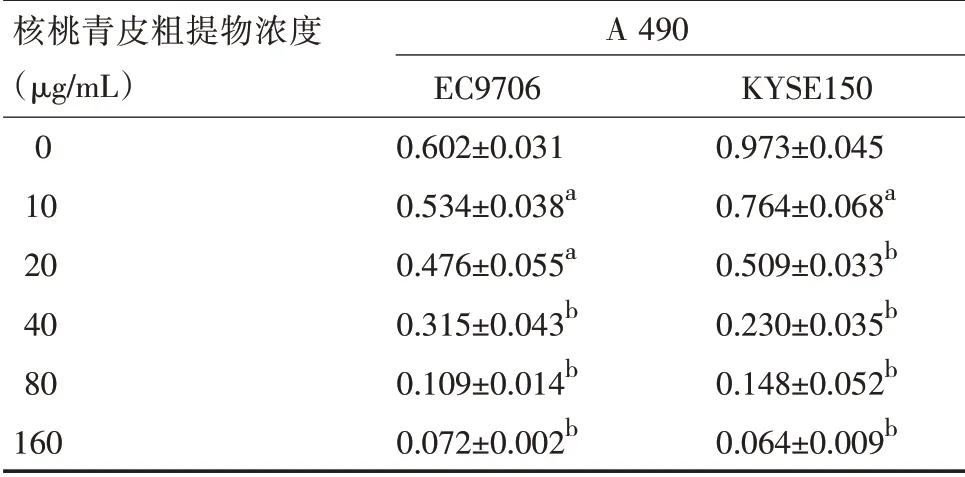
表1 核桃青皮粗提物对EC9706和KYSE150细胞增殖的抑制作用(±s,n=5)
注:与阴性对照组比较,a P<0.05,b P<0.01
核桃青皮粗提物浓度(μg/mL)0 10 20 40 80 160 A 490 EC9706 0.602±0.031 0.534±0.038a 0.476±0.055a 0.315±0.043b 0.109±0.014b 0.072±0.002b KYSE150 0.973±0.045 0.764±0.068a 0.509±0.033b 0.230±0.035b 0.148±0.052b 0.064±0.009b
表2 没食子酸对EC9706和KYSE150细胞增殖的抑制作用(±s,n=5)
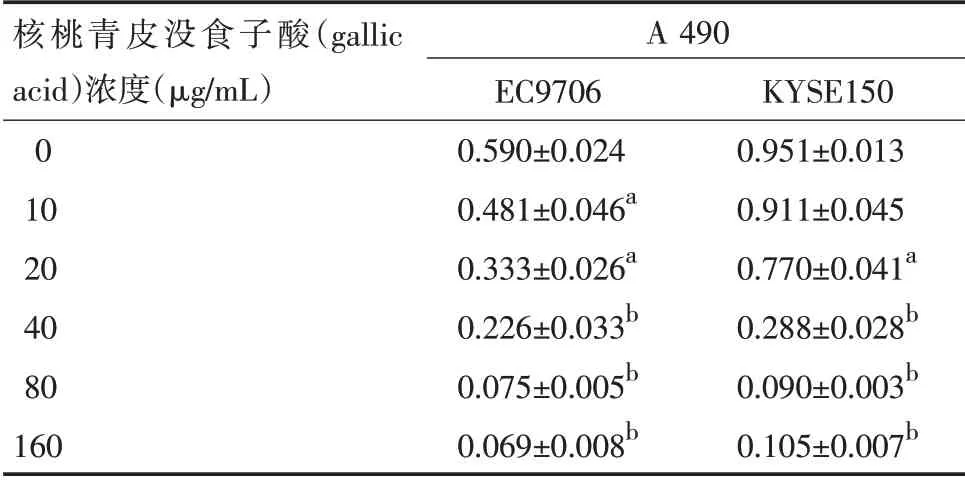
表2 没食子酸对EC9706和KYSE150细胞增殖的抑制作用(±s,n=5)
注:与阴性对照组比较,a P<0.05,b P<0.01
核桃青皮没食子酸(gallic acid)浓度(μg/mL)0 10 20 40 80 160 A 490 EC9706 0.590±0.024 0.481±0.046a 0.333±0.026a 0.226±0.033b 0.075±0.005b 0.069±0.008b KYSE150 0.951±0.013 0.911±0.045 0.770±0.041a 0.288±0.028b 0.090±0.003b 0.105±0.007b
表3 乙酸乙酯对EC9706和KYSE150细胞增殖的抑制作用(±s,n=5)
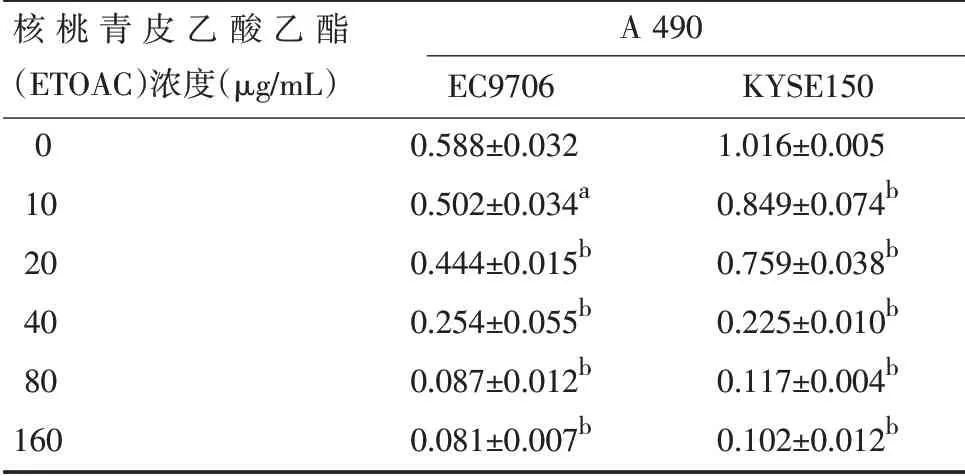
表3 乙酸乙酯对EC9706和KYSE150细胞增殖的抑制作用(±s,n=5)
注:与阴性对照组比较,a P<0.05,b P<0.01
核桃青皮乙酸乙酯(ETOAC)浓度(μg/mL)0 10 20 40 80 160 A 490 EC9706 0.588±0.032 0.502±0.034a 0.444±0.015b 0.254±0.055b 0.087±0.012b 0.081±0.007b KYSE150 1.016±0.005 0.849±0.074b 0.759±0.038b 0.225±0.010b 0.117±0.004b 0.102±0.012b
2.3 食管癌细胞KYSE150和EC9706 CyclinD1蛋白表达 各组蛋白表达情况如图3,灰度值统计分析结果见图4。
3 讨论
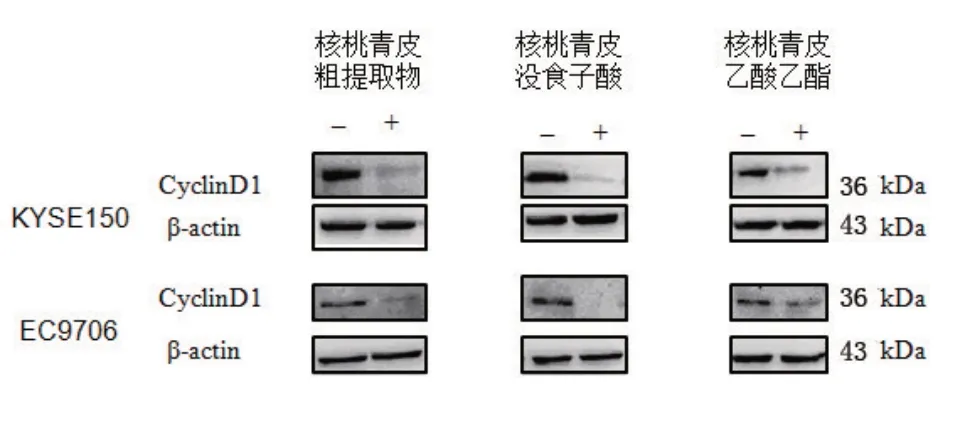
图3 KYSE150和EC9706细胞经3种药物(核桃青皮粗提物,没食子酸和乙酸乙酯)处理后CyclinD1的表达
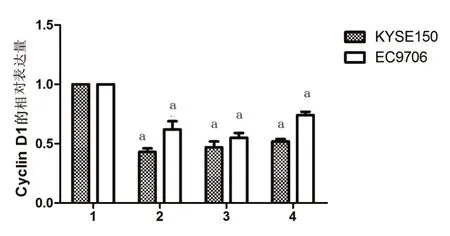
图4 CyclinD1蛋白在不同组别间表达的比较
我国食管癌发病率为19.24/10万,在癌症发病构成中排列第6位;同期食管癌死亡率为15.39/10万,在癌症死亡原因中列第4位[3]。胡桃属植物具有抗肿瘤作用[4]。核桃青皮家族核桃楸在治疗食道癌、肝癌、肠癌均有明显效果[5],青龙衣(核桃青皮)胶囊含药血清对人胃腺癌SGC-7901、肺癌A-549、乳腺癌MCF-7细胞均有不同程度的抑制作用,对人胃腺癌SGC-7901荷瘤裸鼠具有一定的体内抑瘤作用[6]。本实验观察了核桃青皮粗提物和4种核桃青皮提取物对食管癌细胞EC9706和KYSE150增殖的影响,结果发现:核桃青皮粗提物和核桃青皮提取物没食子酸、乙酸乙酯均能抑制食管癌细胞的增殖,且抑制作用随着浓度的增加而增强,表现出明显的剂量依赖性。
细胞增殖是通过细胞周期调控实现的,而细胞周期的有序运行是通过相关基因的严格监视和调控的。其中G1/S是细胞周期中最重要的调控点而CyclinD1在G1/S期起重要作用,它对G1期的调控是通过对抑癌基因蛋白pRb的调控实现的,pRb在G 1期处于低磷酸化状态(功能状态),通过与发动S期的重要分子如转录因子E2F等结合,抑制进入S期所需蛋白的相关基因的转录,使细胞停滞于S期,当外源性增殖分裂信号与相应受体结合后,通过信号传递促进CyclinD1基因表达,使CyclinD1蛋白与CDK4/6结合使之活化,以CDK4/6为催化亚基、CyclinD1为调节亚基形成激酶复合体[7],并结合pRb,在催化亚基CDK4/6作用下,对功能状态下的pRb进行磷酸化,使之失活并释放出转录因子E2F,E2F脱离pRb蛋白后发挥强大的转录调节作用,激活A-DNA聚合酶c-myc、cdc-2等的转录,使细胞越过G1/S限制点进入S期。在生理状态下[8],细胞进入S期后CyclinD1迅速分解。如果CyclinD1基因激活,则CyclinD1持续高表达,将导致G1期缩短,提前进人S期,使细胞增殖失控,最终形成肿瘤。我们利用western blot检测核桃青皮提取物对食管癌细胞CyclinD1的表达,发现核桃青皮粗提取物和核桃青皮提取物没食子酸、乙酸乙酯都能降低CyclinD1在食管癌细胞中的表达,诱导细胞周期阻滞,从而抑制细胞增殖。
近年研究发现在恶性肿瘤治疗中植物原料药比化学合成药更具优势[4],我们的研究提示核桃青皮粗提物和核桃青皮提取物没食子酸、乙酸乙酯都有可能成为临床抗食管癌新药。
[1]张野平,杨志博,景永奎,等.胡桃醌对肿瘤细胞的增殖抑制作用和抗菌作用[J].沈阳药学院学报,1993,10(4):271-274.
[2]司传领,刘忠,惠岚峰,等.核桃楸树皮提取物的化学成分及其抗氧化活性研究[J].林产工业与化学,2008,28(1):29-32.
[3]张思维,张敏,李光琳,等.2003-2007年中国食管癌发病与死亡分析[J].中国肿瘤,2012,21(4):241-247.
[4]李庆勇,王丽敏,祖元刚,等.抗肿瘤植物原料药的现状与发展趋势[J].中国药房,2009,20(3):228-230.
[5]李中原,高奎滨.青龙衣治疗食管贲门癌120例临床观察[J].中医药信息,1988,3(1):31-33.
[6]段玉敏,张志华,王伟明.青龙衣胶囊抗肿瘤活性研究[J].科技致富向导,2013,24:20-21.
[7]Holley SL,Heighway J,Hoban PR.Induced expression of hu⁃mans CCND1 alternative transcripts in mouse Cyl-1 knockout fi⁃broblasts highlights functional differences[J].Int J Cancer,2005,114(3):364-370.
[8]Motokura T,Bloom T,Kim HG,et al.A novel cyclin encoded by abcl1-linked candidate oncogene[J].Nature,1991,350(6318):462-463.
