杜仲IPI基因全长cDNA克隆与生物信息学分析
2014-01-19刘攀峰杜红岩
刘攀峰,吴 敏,杜红岩
(1.中国林业科学研究院 经济林研究开发中心,河南 郑州 450003;2.国家林业局杜仲工程技术研究中心,河南 郑州 450003)
萜存在于几乎所有的生命形式中,是自然界最大的一类天然产物家族,也是植物中种类最为繁多的一类天然产物[1]。萜类骨架以异戊二烯(isoprene)为基本单元,它的生物合成是由异戊 二 烯 焦 磷 酸(isopentenyl diphosphate,IPP)与其异构体二甲基丙烯基焦磷酸(dimethylallyl diphosphate,DMAPP),以及二者缩合形成中间产物相互之间的聚合反应[2]。异戊烯基焦磷酸异构酶(isopentenyl diphosphate isomerase,IPI)催化IPP的碳碳双键与DMAPP亲电丙烯基间的可逆转化,被视作整个萜类代谢网络的一个枢纽[3-4]。目前认为有2类结构迥异的非同源IPI蛋白家族[5-6],I型IPI在植物中广泛存在并且基因序列十分保守,多有2个或2个以上基因亚型存在,序列较长的I型IPI基因多定位于质体,而较短的多定位于细胞质;此外,线粒体、过氧化物酶体也发现有IPI存在。IPP可从细胞质向各种细胞器中扩散,有研究推断是由于IPP有膜通透性,也可能是存在一种特殊的转运机制。II型IPI基因序列较长,一般编码300个氨基酸以上,其活性位点几乎分布于整个氨基酸序列[4]。
目前认为IPI为关键基因调控萜类代谢途径,大肠杆菌Escherichia coli中通过外源表达IPI基因发现番茄红素与类胡萝卜素产量明显提高[7-8];山榛Corylus avellanaL.IPI基因在根、茎、叶中均有表达且以根表达量相对较高,并有试验推断CaIPI能影响植株中紫杉醇的合成[9];而有关橡胶Hevea brasiliensis(Willd.ex A.Juss.)Muell.Arg.的研究认为IPI基因的表达情况是萜类合成路径的关键步骤,并认为创伤不会影响植株IPI基因的表达水平[10]。也有试验指出,外界刺激对IPI活性也产生一定影响,在高光及高盐环境下会诱导烟草Nicotiana tabacumL.IPI mRNA的合成,但是否会影响IPI活性尚待进一步证实。
杜仲Eucommia ulmoidesOliv.是世界上极少数分布于温带、亚热带能生产优质天然橡胶的木本植物[11-12]。杜仲胶、京尼平苷和桃叶珊瑚苷等诸多杜仲萜类具有重要的产业应用价值,在橡胶工业、航空航天、国防、船舶、化工、医疗、体育等国民经济许多部门引起广泛关注[13-16]。对杜仲IPI基因结构与功能开展的研究不多,本研究试通过阐明杜仲IPI基因序列、结构特征并分析其作用机理,以期为杜仲萜类生物调控和分子育种提供基础资料。
1 材料与方法
1.1 材 料
2011年4月下旬,在中国林业科学研究院经济林研究开发中心采集 “华仲6号”杜仲幼嫩叶片,用清水冲洗表面杂质后,带回室内于液氮中-80℃保存备用。
1.2 试 剂
MightyAmp DNA Polymerase Ver.2(Takara,大连),3’-Full RACE Core Set(Takara,大连),M-MLV RTase cDNA Synthesis Kit(Takara,大连),5’-Full RACE Kit(Takara,大连),氯化锂(Amresco,美国),CTAB(Amresco,美国),琼脂糖凝胶DNA回收试剂盒(天根,北京),DH5α感受态细胞(天根,北京),SDS(上海生工,上海),DEPC(Sigma,德国),pEASY-T1 Cloning Kit(全式金,北京),PVP(Amresco,美国)。
1.3 试验方法
1.3.1 杜仲叶片RNA提取及单链cDNA的合成
通过改良CTAB-LiCl法进行杜仲叶片RNA的提取[17],并依照M-MLV RTase cDNA Synthesis Kit进行杜仲单链cDNA的合成。
1.3.2 引物设计
由杜仲转录组测序中已知的IPI unigene序列,设 计 引 物 5′-TCCTTCTCTATTCGCCTCATCC-3′/5′ATGTTCGCCCCACTTCC-3′扩 增 IPI基 因 特异片段;后结合试剂盒锚定引物序列,设计3′RACE 巢 式 扩 增的 引 物 5′-CTCCTTCAGC AACGATCAGG-3′/5′-CTGAG GATGTTCCAGT CGAT-3′,以及 5′RACE 巢式扩增引物 5′-CGTCC ACACCAAAGGGAATGTTAC-3′/5′-CCACCAAA ATACATTCGTCCTC-3′。
1.3.3 基因全长的末端扩增
基因全长末端扩增反应体系与反应条件参照Takara 3′-Full RACE Core Set Ver.2.0 与 Takara 5′-Full RACE Kit 说明操作。
1.3.4 基因片段回收与测序
基因片段回收按TIANGEN通用型DNA回收试剂盒进行,扩增片段连接按pEAZY-T克隆试剂盒进行,基因片段测序由金斯瑞公司(南京)完成。
1.3.5 生物信息学分析
序列相似性检索由NCBI(http://www.ncbi.nlm.nih.gov)Blast程序完成;蛋白质氨基酸组成、相对分子质量、理论等电点以及功能位点分析由ExPASy(http://cn.expasy.org)ProtParam 程 序 与ScanProsite程序完成;蛋白质细胞定位及二级结构预测由Predict Protein(http://www.predictprotein.org/)以 及 PSIPRED方 法(http://bioinf.cs.ucl.ac.uk/psipred)完成[18];转运肽预测由ChloroP 1.1 Server(http://www.cbs.dtu.dk/services/ChloroP/)完成[19];蛋白质同源建模由SWISS-MODEL程序(http://swissmodel.expasy.org/)完 成[20];蛋白质序列多重比对由Lasergene软件完成;基因系统进化树构建由MEGA 5软件完成[21];氨基酸序列无序性分析由在线程序FoldIndex(http://bip.weizmann.ac.il/ fl dbin/ fi ndex)完成[22]。
2 结果与分析
2.1 杜仲IPI基因全长cDNA序列特征
在逆转录cDNA模板上分别扩增出3条杜仲IPI基因特异片段,测序拼接后得到1条1 231 bp的基因序列,如图1、图2所示。与喜树Camptotheca acuminataDecne.(AF031079.1)、欧洲榛Corylus avellanaL.(EF553533.1)、番薯Ipomoea batatas(L.)Lamarck(AB499048.1)、 番 茄Lycopersicon esculentumMiller.(NM_001247924.1)、长 春 花Catharanthus roseus(L.)G.Don.(EU135981.1)、葛 根Pueraria lobataOhwi.(AY315650.1)IPI全长基因序列的相似性分别为84%、84%、83%、84%、83%、82%。通过ORF fi nder程序查到1个长921 bp的开放阅读框,与欧洲榛(ABW06959.1)、葡萄Vitis viniferaL.(XP_002277935.1)、烟 草Nicotiana tabacumL.(BAB40974.1)、番茄(NP_001234853.1)、丹参Salvia miltiorrhizaBunge(ABV08818.1)、番薯(BAI47570.1)IPI氨基酸序列的相似性分别为78%、77%、72%、92%、86%、91%,确定为杜仲IPI基因cDNA全长序列,将其命名为EuIPI。
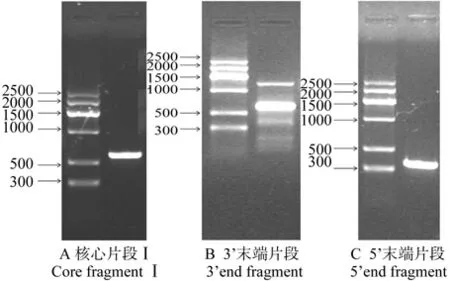
图1 EuIPI基因RT-PCR扩增的琼脂糖凝胶电泳Fig.1 Agarose gel electrophoresis of RT-PCR ampli fi cation of EuIPI

图2 EuIPI全长cDNA及推导的氨基酸序列Fig.2 Full-length sequence of EuIPI cDNA and the deduced amino acid sequences
2.2 EuIPI编码蛋白结构特征
2.2.1 EuIPI蛋白的一级结构
预测编码蛋白相对分子质量为34.65kD,理论等电点5.40;氨基酸组成以亮氨酸(12.7%)、丝氨酸(10.1%)、缬氨酸(6.9%)和丙氨酸(6.9%)含量最高,蛋白不稳定系数为42.38,属不稳定蛋白质。预测EuIPI亚细胞定位于叶绿体上,其中TargetP 1.1 Server预测分值为0.945,可靠性为Ⅱ级。多重比对结果显示,EuIPI氨基酸序列包含植物IPI蛋白保守的NTCCSHPL基序和WGEHELDYLL基序,以及催化过程中所需要的Mg2+结合位点(A154、A175)和Zn2+结合位点(A107、A119、A156、A196、A206、A208)等功能位点。5 种同源IPI序列多重比对后相同的氨基酸位点达47个,植物IPI蛋白N端比大肠杆菌多出一段80~90个氨基酸残基的蛋白序列,说明在这段区域中可能存在转运肽,推导EuIPI蛋白转运肽序列长70个氨基酸残基,去除转运肽序列后EuIPI蛋白的相对分子质量为27.25kD(见图3)。
2.2.2 EuIPI蛋白二级结构分析
在线预测EuIPI蛋白二级结构α-螺旋22.55%,β-折叠13.40%,螺环结构64.05%,属混合型蛋白质(见图4)。分析结果显示,EuIPI为多结构域蛋白,分属Nudix水解酶蛋白家族,包括金属离子结合位点和nudix 基序等功能结构(见图5)。
2.2.3 EuIPI编码蛋白的三级结构预测

图3 推导EuIPI 同源序列的多重比对Fig.3 Multiple alignment of the deduced EuIPI homologous sequences

图4 推导EuIPI蛋白二级结构预测Fig.4 Prediction of secondary structure of the deduced EuIPI protein
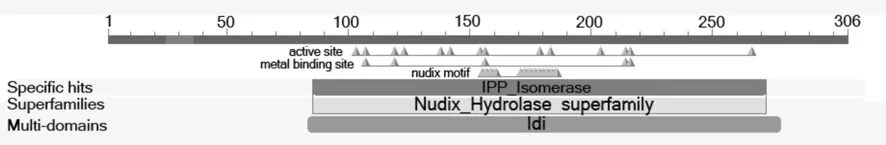
图5 推导EuIPI蛋白的保守结构域预测Fig.5 Prediction of conserved domains of the deduced EuIPI protein
以人Homo sapiens IPI蛋白(2i6k)为模板对EuIPI蛋白质同源建模,程序评测推导的EuIPI蛋白模型QMEAN4得分0.715,与模板蛋白序列相似性50.66%,从三维模型图中可以看出EuIPI空间上以单体形式存在,主要保守基序位于分子结构中部(见图6)。
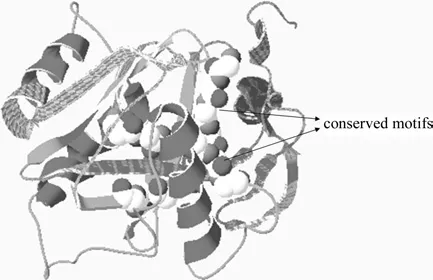
图6 推导EuIPI蛋白三级结构的预测Fig.6 Prediction of tertiary structure of the deduced EuIPI protein
2.3 EuIPI蛋白功能位点预测
程序分析EuIPI基序类型分4种,含10个潜在的功能位点,包括2个蛋白激酶C磷酸化位点(SnK,26-28;SfR,50-52);2个N端糖基化位 点(NKSR,27-30;NQSA,303-306);3个N端豆蔻酰化位点(GAarTR,43-48;GTkvTF,140-145;GVrnAA,171-176);3个酪蛋白激酶Ⅱ磷酸化位点(SgmD,73-76;SkyE,129-132;TlqE,286-289)。软件预测出18个EuIPI蛋白翻译后磷酸化修饰位点,包括11个丝氨酸磷酸化位 点(A16、A17、A26、A29、A31、A33、A36、A41、A49、A50、A73),2个苏氨酸磷酸化位点(A102、A141)以及5个酪氨酸磷酸化位点(A34、A104、A131、A159、A241)(见图 7)。
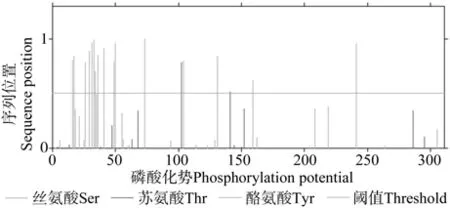
图7 推导EuIPI蛋白翻译后磷酸化位点预测Fig.7 Prediction of phosphorylation sites of deduced EuIPI protein
2.4 EuIPI蛋白的系统进化树构建
利用 ClustalW法对20个IPI同源蛋白进行序列比对,并用Neighbor-joining法构建系统进化树,如图8所示。发现不同来源的IPI蛋白在进化上具有较明显的界限。其中推导EuIPI蛋白与欧洲榛IPI蛋白亲缘关系最为接近,进化距离为0.061,其次为水稻Oryza latifoliaDesv.(0.075)、丹参(0.082)、玉米Zea maysL.(0.082)、毛果杨Populus trichocarpaTorr.& Gray.(0.082)和喜树(0.089)。

图8 IPI同源蛋白的系统进化树构建Fig.8 Construction of phylogenetic tree of IPI homologous protein
2.5 EuIPI蛋白氨基酸序列的无序性分析
FoldIndex无序性分析对EuIPI氨基酸序列中共预测出2个无序化区域,分别为A87~A97,A218~A240,总的无序化氨基酸数目为34个,无序化比例达11.11%,最长的1段无序化区域包含23个氨基酸残基(见图9)。
2.6 EuIPI编码蛋白的功能的初步预测
Profun 2.2 Server预测蛋白功能的结果见表1。从表1可看出,EuIPI作为酶蛋白的概率(0.770)和比数(2.690)大于非酶蛋白,并可能属于连接酶类,参与氨基酸生物合成的可能性(概率为0.282;比数为12.813)最高,而作为植物免疫反应基因本体的可能性(概率为0.147;比数为1.727)最大。

图9 EuIPI氨基酸序列的无序化分析Fig.9 Disordered analysis of the deduced EuIPI amino acid sequence
3 讨 论
诸多植物萜类合成都需要有IPI异构酶的参与,植物中IPI基因最早于1995年在花香基因模式植物仙女扇Clarkia breweri中分离出,后在拟南芥全基因组发现2个IPI编码基因,并将其分别命名为IPI1和IPI2[5,23],至今有近2 000余条IPI基因相关核酸序列在GenBank注册,包括细菌、古细菌以及真核生物等。序列分析结果说明本研究从杜仲中分离出的EuIPI与其它植物IPI基因有很高的同源性,所推导EuIPI氨基酸序列包含植物IPI蛋白保守的NTCCSHPL基序和WGEHELDYLL基序,并推测出多个潜在功能位点,分析结果预示从杜仲中分离出的EuIPI为一功能蛋白的编码基因,是高等植物IPI基因家族的新成员。
有39种IPI蛋白晶体模型在PDB数据库中注册,目前认为有2类结构迥异的非同源蛋白家族作用于IPP与DMAPP间的异构化过程。I型IPP蛋白于20世纪50年代首先被发现,存在于几乎所有真核生物及细菌中,其反应过程需要2价金属离子的参与。与II型IPI相比,I型IPI蛋白序列较短,具有保守的NXXXCXHP与EXE基序以及富甘氨酸序列,其催化机理可能依赖于通过生成3位碳正离子中间产物而实现的质子化与去质子作用,这个机制此前较少提出但得到来自生物信息学和酶学的试验证据的支持[3,24]。II型IPI发现较晚,被认为属于黄素酶家族,常见于缺氧、高盐、高温、高酸以及贫瘠等严酷条件生长的细菌及所有已知的古细菌中,特别是嗜热菌Thermophilic bacteria和超嗜热菌中极为普遍。II型IPI反应过程需要黄素蛋白、NAD(P)H以及2价金属离子的参与,NAD(P)H被认为用于还原黄素蛋白并生成IPI蛋白与还原型FMN的中间产物,但黄素蛋白在反应过程中的作用机制目前尚不清楚。IPP与DMAPP之间最为可能的转换机制为依赖于1,3位氢原子异位时的立体选择性,但其异构化过程究竟以那类IPI酶为主导至今还没有定论[3]。序列分析说明本研究EuIPI编码蛋白属于I型IPP蛋白,但其在杜仲萜类生物合成所扮演的角色尚待进一步确定。
[1] Eric O,Lin F Y.Terpene biosynthesis:modularity rules[J].Angewandte Chemie,2012,51(5):1124-1137.
[2] Nobuaki S,Hirotaka U,Takashi N,et al.Construction and analysis of EST libraries of the trans-polyisoprene producing plant,Eucommia ulmoidesOliver[J] Planta,2012,236(5):1405.
[3] Zheng W,Sun F,Mark B,et al.The crystal Structure of human isopentenyl diphosphate isomerase at 1.7 Å resolution reveals its catalytic mechanism in isoprenoid biosynthesis[J].J Mol Biol,2007,366:1447-1458.
[4] Karine B,Yannick E,Alain D,et al.Isopentenyl Diphosphate Isomerase: A checkpoint to isoprenoid biosynthesis[J].Biochimi,2012,94(8):1621-1634.
[5] Michael C,Frederick MH,C Dale P,et al.Analysis of the isopentenyl diphosphate isomerase gene family fromArabidopsis thaliana[J].Plant Mol Biol,1998,36:323-328.
[6] Raphaël D,Jérǒme de R,Virginie,et al.Overexpression,physicochemical characterization,and modeling of a hyperthermophilic pyrococcus furiosus type 2 IPP isomerase[J].Proteins,2008,71(4):1699-1707.
[7] Jin S,Yu-Yang Z,Hui L,et al.A novel cytoplasmic isopentenyl diphosphate isomerase gene from tomato (Solanumlycopersicum): cloning,expression,and color complementation[J].Plant Mol Biol Rep,2010,28:473-480.
[8] 化文平,王喆之.向日葵异戊烯焦磷酸异构酶基因(ipi)的电子克隆和生物信息学分析[J].植物生理学通讯,2008,44(1):8l-86.
[9] 汪业春,唐克轩.山榛中异戊烯焦磷酸异构酶基因的克隆及分析[J].中国农业科技导报,2007,9(4):95-100.
[10] Soo KO,Hunseung K,Dong HS,et al.Molecular cloning and characterization of a functional cDNA clone encoding isopentenyl diphosphate isomerase fromHevea brasiliensis[J].J Plant Physiol,2000,157:549-557.
[11] 杜红岩,胡文臻,俞 锐.杜仲产业绿皮书:中国杜仲橡胶资源与产业发展报告[M].北京:社会科学文献出版社,2013.
[12] 杜红岩,李 钦,杜兰英,等.杜仲雄花茶营养成分的测定分析[J].中南林业科技大学学学报,2007,27(6):88-91.
[13] 刘攀峰,杜红岩,乌云塔娜,等.杜仲1-脱氧-D-木酮糖-5-磷酸还原异构酶基因cDNA全长克隆与序列分析[J].林业科学研究,2012,25(2):195-200.
[14] 杜红岩,刘昌勇,李 钦,等.杜仲叶中3 种主要活性成分含量的季节变化[J].中南林业科技大学学报,2013,31(8):6-9.
[15] 杜红岩.杜仲活性成分与药理研究的新进展[J].经济林研究,2003,21(2):58-61.
[16] 杜红岩.我国的杜仲胶资源及其开发潜力与产业发展思路[J].经济林研究,2010,28(3):1-6.
[17] 周明兵,王红珍,赵德刚.杜仲叶和树皮总RNA的快速提取法[J].山地农业生物学报,2003,22(5):430-431.
[18] Jones DT.Protein secondary structure prediction based on position-speci fi c scoring matrices [J].J Mol Biol,1999,292(2):195-202.
[19] Emanuelsson O,Nielsen H,von Heijne G.ChloroP,a neural network-based method for predicting chloroplast transit peptides and their cleavage sites[J].Protein Sci,1999,(8): 978-984.
[20] Arnold K,Bordoli L,Kopp J,et al.The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling[J].Bioinformatics,2006,22:195-201.
[21] Tamura K,Peterson D,Peterson N,et al.MEGA5: Molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Mol Biol Evol,2011,28:2731-2739.
[22] Prilusky J,Felder CE,Zeev-Ben-Mordehai T,et al.A simple tool to predict whether a given protein sequence is intrinsically unfolded[J].Bioinformatics,2005,21(16):3435-3438.
[23] Blanc V M,Pichersky E.Nucleotide sequence of a Clarkia breweri cDNA clone of IPI1,a gene encoding isopentenyl pyrophosphate isomerase[J].Plant Physiol,1995,108(2): 855-856.
[24] Raphael D,Jerome R,Virginie D,et al.Overexpression,physicochemical characterization,and modeling of a hyperthermophilic pyrococcus furiosus type2 IPP isomerase[J].Proteins,2008,71: 1699-1707.
