滇楸花部性状的表型多样性研究
2013-12-28姚淑均张守攻王军辉张乃春张明刚
姚淑均,张守攻,王军辉,张乃春,张明刚
(1.林木遗传育种国家重点实验室,中国林业科学研究院林业研究所;国家林业局林木培育重点实验室,北京100091;2.贵州省林业科学研究院,贵州 贵阳 550005;3.贵州省林业种苗站,贵州 贵阳550001)
滇楸花部性状的表型多样性研究
姚淑均1,2,张守攻1,王军辉1,张乃春3,张明刚2
(1.林木遗传育种国家重点实验室,中国林业科学研究院林业研究所;国家林业局林木培育重点实验室,北京100091;2.贵州省林业科学研究院,贵州 贵阳 550005;3.贵州省林业种苗站,贵州 贵阳550001)
在贵州滇楸Catalpa duclouxii Dode.主要分布区收集了17个群体的花,对花枝长度、花朵数量、上下花萼长度及花冠(花径)大小等7个表型性状进行系统调查和遗传多样性分析。结果表明:花的表型性状在群体间和群体内均存在丰富的变异,7个性状的差异均达到极显著水平(P<0.01),平均变异系数为25.94%,变化幅度为7.85~47.17%,平均表型分化系数为27.95%(14.92~42.15%),群体内变异(72.05% )大于群体间变异(27.95%);花性状与经纬度、海拔呈显著、极显著相关;通过聚类分析17个群体划分为4个类。
滇楸;花;形态特征;表型性状
滇楸Catalpa fargesii f. duclouxii (Dode)属梓属楸树组,落叶乔木,树高达30 m。滇楸花期3~5月,果成熟期9~11月,圆锥花序或聚伞状圆锥花序,蒴果长约70~100 cm。滇楸不仅树姿宏伟,高大挺拔,枝叶繁茂,花色艳丽,自古人们就把它作为绿化观赏树种,广泛栽植于皇宫庭院、刹寺庙宇、胜景明园之中,而且木材材质优良,纹理直,不翘不裂,耐腐 、耐潮 ,是极好的室内装饰、建筑、家具、器具、 造船、雕刻、乐器等优良用材。其分布在云南、贵州、四川、湖南、湖北、广东等省,浙江、南京、山东、陕西、河南、福建等省均有引种栽培[1]。目前有关滇楸的研究主要在分布、引种、苗木繁殖技术、胚胎学、花粉特性、种子发芽率、气孔性状、苗期遗传测定、幼苗期抗旱、生理与光合特性等[2-14],有关滇楸花部性状表型变异研究未见报道。
随着现代社会对木材的需求,滇楸的资源变得越来越少。为充分挖掘滇楸资源、特别是用材与观赏相结合型的滇楸选育利用。在贵州滇楸主要分布区贵阳、兴仁、贵定、安顺、黔西等集中分布地进行关滇楸花部特性的调查。目的在于通过花的结构、表型性状的变异了解其群体的遗传多样性水平和结构,掌握其影响因素及其变化规律[15]。有利于开展种质资源多样性的保护,为滇楸的杂交育种、抗性育种和定向培育提供材料和依据,为滇楸种质资源基因库核心种质的构建提供参考。
1 材料与方法
1.1 滇楸群体的选择与材料采集
2009~2011年的3~5月多次在贵州滇楸主要分布区进行资源调查。选取植株分布较集中的17个群体(见表1)。在选定的各群体内随机选择10株生长正常、无明显病虫害、盛花期的植株,株间不同花色水平距离50 m以上,每个植株选取树冠中上部靠南部开花的枝条,每个花序选取中部两朵花进行性状指标测量。对选定群体的采样植株采用全球卫星定位系统(Global Positioning System,GPS)定位。
1.2 滇楸花性状的选择与测定方法
滇楸观测的花的主要表型性状包括:花枝长、花序长、单个花序上的小花枝数、单个花序的花朵数、每朵花的上萼长、下萼长、花径(花萼口的宽度)宽共计7个性状。测量时用直尺对当年生花枝、花序进行测量,精度到0.0 cm,用游标卡尺测量花萼和花径长,测量精度0.1 mm。单个花序小花数包括已经开败的和未开放的正常有花和小花枝数于盛花期进行计数。每个植株花枝长、花序长均进行25次重复,花萼长、花径宽均进行50次重复。
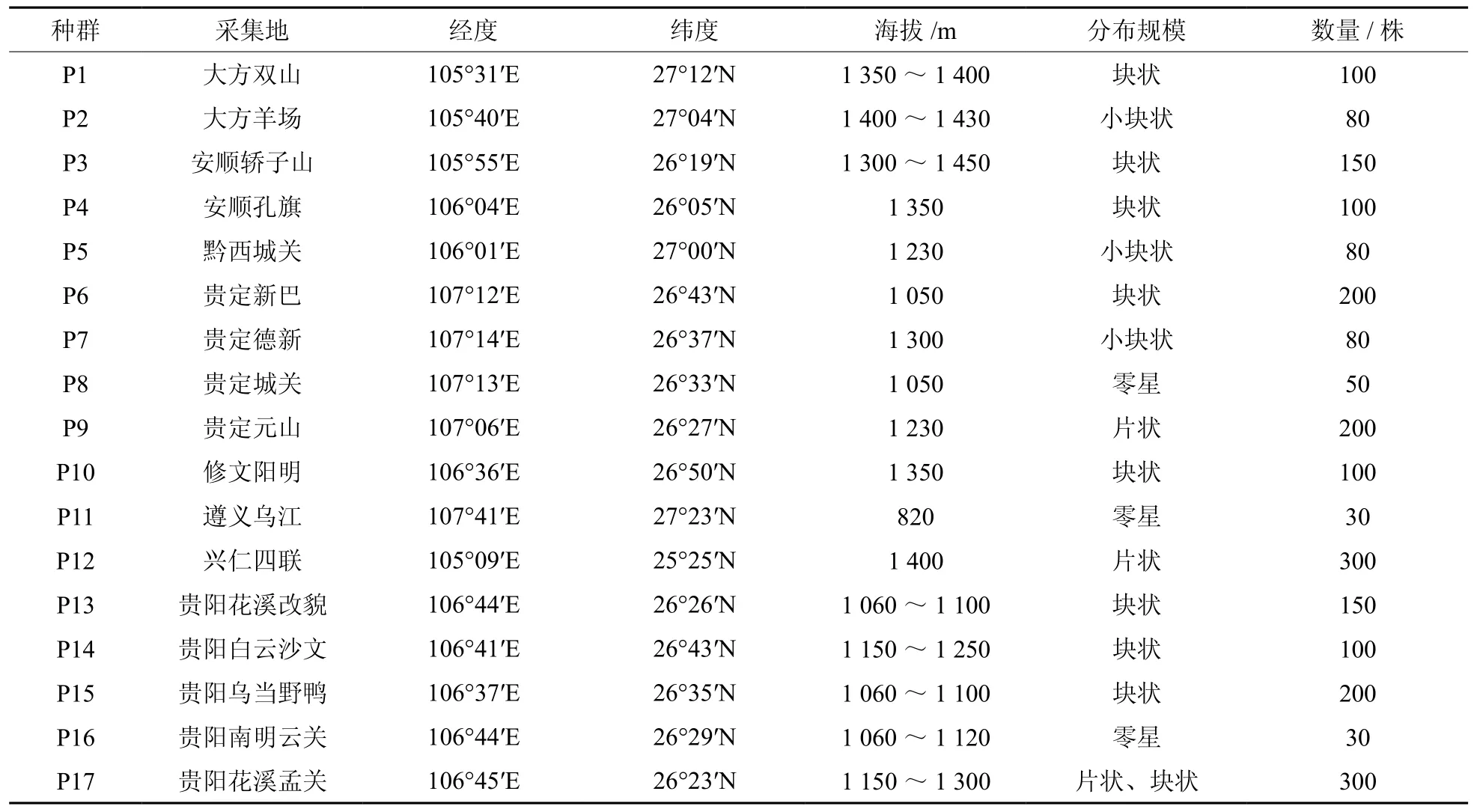
表 1 滇楸群体采样地的地理位置及基本情况Table 1 Geographical positions of sampling sites of C.duclouxii populations in Guizhou and and basic situations
1.3 统计分析方法
应用SAS9.2 软件对滇楸花的表型性状进行巢式方差分析。表型分化系数(VST)表示种群间变异占遗传总变异的百分比,反应种群间的表型分化状况[16-19]。表型特征变异系数(CV)表示表型性状的离散程度,反映表型的变异特征。应用Excel计算各性状的变异系数。应用SPSS16.0软件对滇楸花的表型性状采用欧氏距离进行聚类分析。
2 结果与分析
2.1 滇楸群体间花的形态变异特征
滇楸群体间和群体内花表型性状的变异均方及F值和花的7个表型性状均值、多重比较见表2和表3。花枝、花序长等7个表型性状在群体间和群体内均存在极显著差异。从表中看出,滇楸花的性状特征在群体间存在极显著差异,表示花朵大小指标即花的上下萼唇长和花径宽和代表花序特征的花序上的小花枝数的变化趋势一致。以兴仁、贵定城关、德新种群变异特征最小,且与大方、黔西和贵阳其它群体间的差异较显著,以大方、黔西和贵阳种群特征值为最大。综合起来大方、黔西和贵阳种群群体性状较差,黔西南和贵定种群群体性状较优。
2.2 滇楸群体间花的形态变异特征
用变异系数表示性状值离散性特征,变异系数越大,则形状值离散程度越大 滇楸17个种群(表4)花部7个性状总的平均变异系数为21.77%、变化幅度为17.55%~32.29% ; 其中黔南贵定德新、城关、春光,黔西南兴仁和黔北的乌江、其性状变异丰富(超过23%),群体变异系数最大的兴仁、贵定和遵义,说明这些种群花表型性状的稳定性相对较低,选择潜力很大,即表示这些地区是滇楸的表型多样性分布中心,这与我们平常的调查基本一致。滇楸花性状变异系数变化幅度为7.85%~47.17%,滇楸花性状中上、下萼及花径的变异系数分别为7.85%、7.95%和10.31%,均比较小,说明花性状的稳定性高,花枝长、花序长、花序上小花的数量等变异系数均超过30%,说明其性状的稳定性相对较低,选择潜力较大,而这些性状正好是滇楸开花、结实特性的重要性状。 滇楸不同群体内的每一性状存在一定的差异,说明环境异质性导致了群体表型变异的差异。
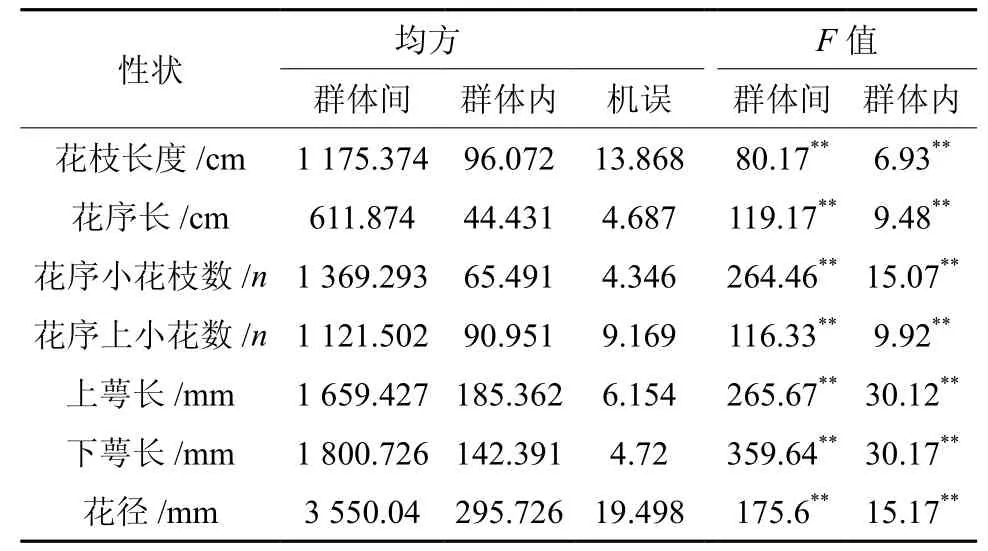
表 2 滇楸花种群间种群内表型形状的方差分析Table 2 ANOVA analysis on phenotype traits of 17 populations of C.duclouxii
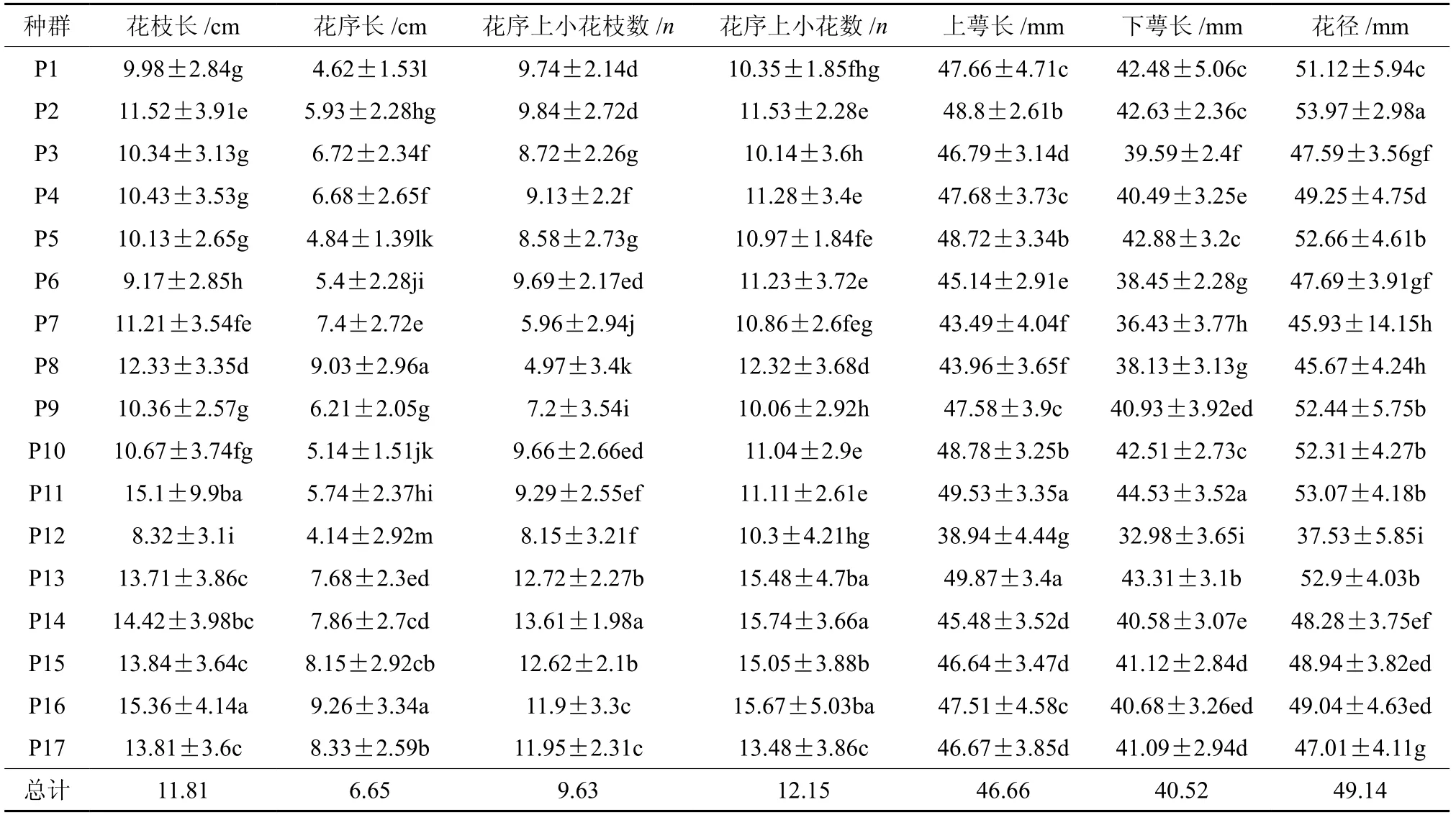
表 3 滇楸群体间花表型性状的平均值及标准差†Table 3 Mean and standard deviation of phenotypic traits of C.duclouxii populations
2.3 滇楸群体间的表型分化
根据巢氏设计计算的方差分量组成比,进一步分析出各性状间方差分量占总变异的百分比(见表5)。滇楸种群花部7个表型性状在群体间和群体内的平均方差分量百分比分别为31.31%和27.26%,差别不是很大。根据种群间方差分量占遗传总变异(种群间和种群内方差分量之和)的百分比表示种群间的分化系数。7个表型性状的分化幅度为18.91% ~ 42.15%,7个表型性状群体内的变异均大于群体间的变异(群体内表型分化系数都大于50%以上)。其中表型分化系数最大的是花的上萼长、最小的是花枝长,它表明花的上萼长性状在滇楸种群间分化最大,花序长则分化最小。7个表型性状种群间平均表型分化系数为28.26%,说明滇楸种群花表型平均变异在种群间的贡献占28.26%,种群内的贡献占71.74%,显然种群内的贡献远远大于种群间的贡献。说明贵州滇楸花部表型性状在种群间的变异程度小,在种群内表型变异程度大,种群内的表型多样性比种群间更丰富,滇楸花的表型变异主要来源于群体内的变异。
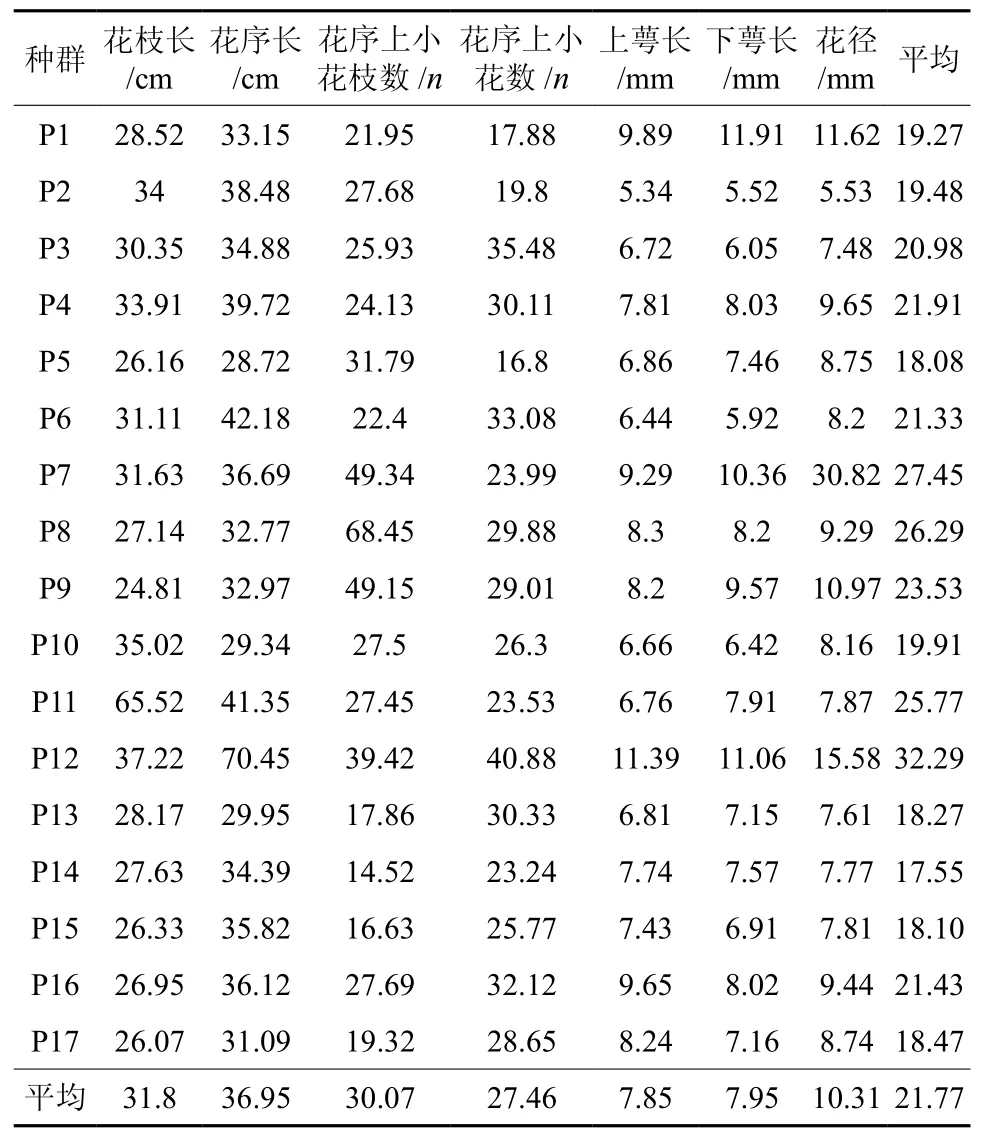
表 4 滇楸群体表型性状的变异系数Table 4 Variation coefficients of phenotypic traits of C.duclouxii populations %
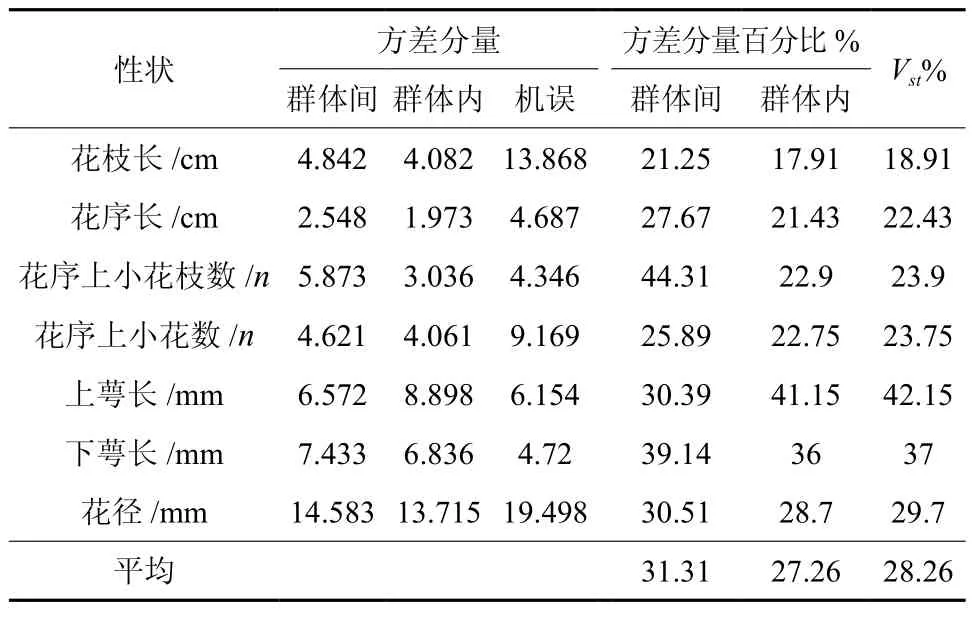
表 5 滇楸种群间花表型性状的方差分量与种群内表型分化系数Table 5 Variance components and phenotype differentiation coefficients ( VST) of morphological traits among/within populations of C.duclouxii
2.4 滇楸花部发育性状与地理因子间的相关分析
对滇楸花发育的7个性状平均值与取样地的地理环境因子纬度进行了相关分析(见图1)。结果表明,表示花朵大小指标的花上萼长、下萼长、花径的变化趋势一致,与纬度呈极显著负相关(R:0.702 、0.749、0.792),花枝长度与纬度显著相关 (R:0.652)、与海拔显著负相关(R:0.697),花序长度与海拔显著负相关(R:0.706)。总之纬度与滇楸花发育各性状值的大小都有明显的相关性。花的上萼长、下萼长、花径与纬度的函数关系分别为:y=-2.167x2+118.5x–1 572;y= -1.191x2+67.59x-914.3;y=-2.912x2+160.7x–2 165。 花 枝长、花序长与纬度、海拔的函数关系分别为:y =1.595x2-82.57x+1 072;y=-4E-05x2-0.002 8x+178.12;y=0.000 2x2-0.590 3x+416.02。
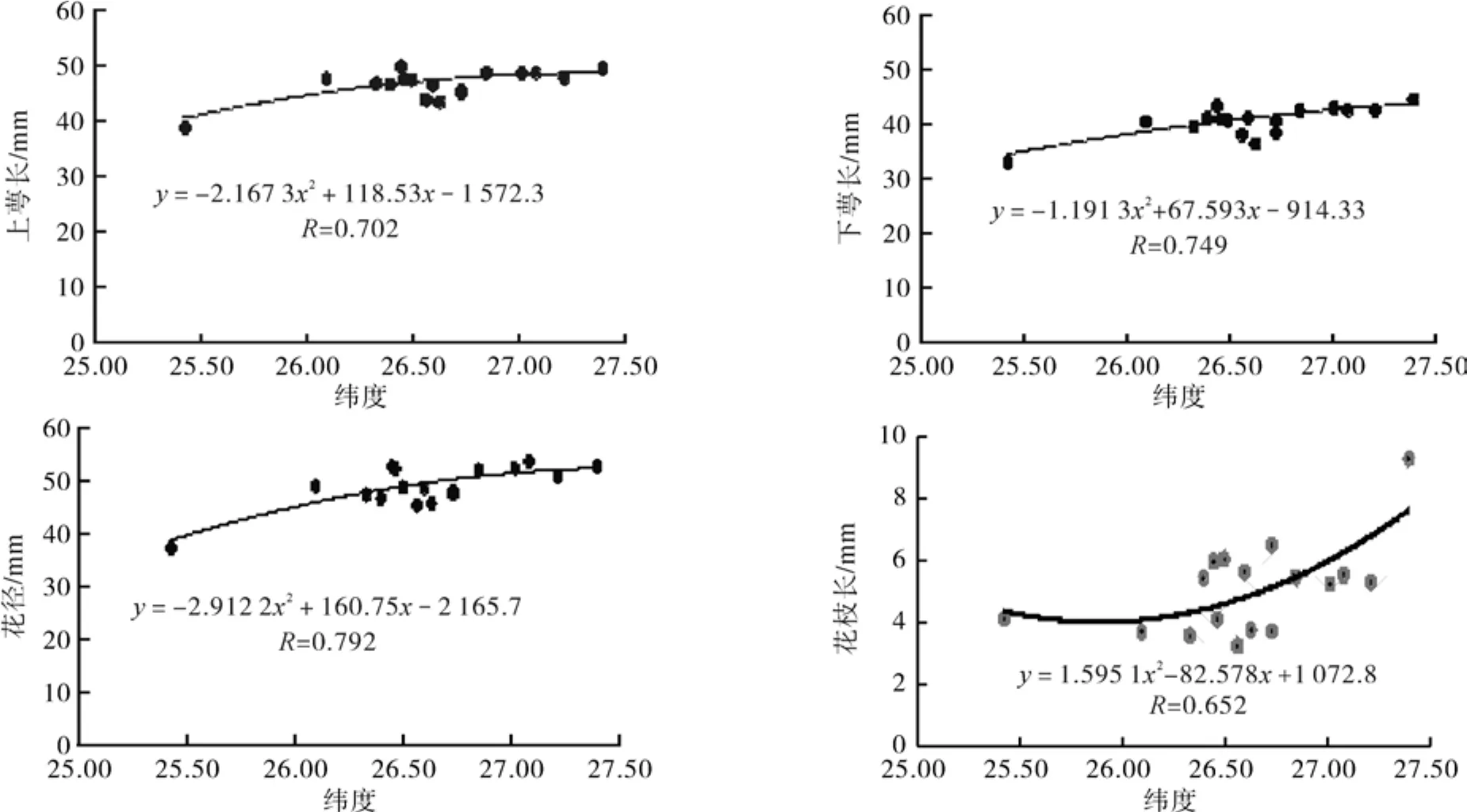
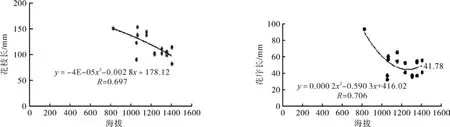
图1 花部性状与纬度、海拔关系的趋势Fig.1 Relations between fl ower characters and latitude, altitude
2.5 滇楸群体花发育性状间的相关关系
对滇楸花发育的表型性状间的相关关系进行分析(见表6),结果表明:花序着生处花枝的长度与大多数性状之间存在极显著的相关性,与花序长、小花数呈极显著相关,与小花枝数呈显著相关。其中反映花朵数量多少的性状小花朵数与花枝长、花序长、花序小花枝数量变化趋势一致,均呈极显著的相关性(R:0.789、0.733、0.754),花朵大小与花发育基础的性状间的相关关系反映了花发育过程中各部位差异化协调发育的特点。
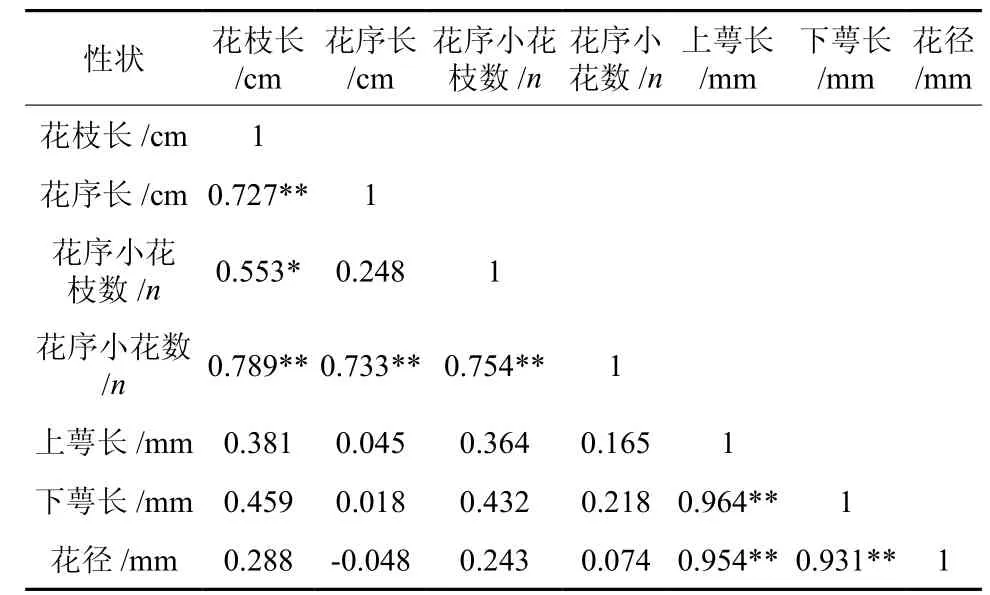
表 6 滇楸花发育性状间的相关关系†Table 6 Correlation among flower characters of C.duclouxii
2.6 滇楸群体内花表型性状的聚类分析
利用欧式平均距离及非加权算术平均法(UPGMA)对滇楸17个种群花部7表型性状进行了聚类分析(见图1)。 根据聚类结果,17个滇楸种群被分为4类。黔西北大方的双山、羊场、黔中安顺轿子山、孔旗、黔中贵阳南明区的云关、花溪区改貌、孟关、乌当区野鸭群、白云区沙文、黔西城关、修文阳明、贵定新巴和德新种群等共13个种群聚在一起为第一类。这类群体的花枝变异相对较大,花序长度都接近或大于平均平均值;贵定城关和德新两个种群聚一起聚为第二类,该类群体花序上的小花枝数量最少,花上、下萼长度及花径较小,花朵较小;分布较北的遵义乌江种群为第三类,该种群花上下萼长在所有种群中最长,花序较短;分布于南边的兴仁种群为第四类,花较小,花枝、花序长度、花上、下萼长度、花径的平均值变化趋势一致,在所有种群中最短,这与他位于西南部,春季干旱、降雨少,且花期比贵州其它地区开花早(提前一个月)有关。
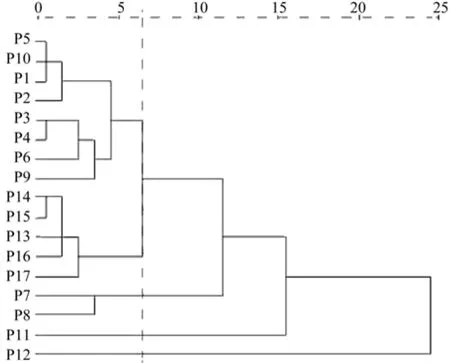
图2 滇楸群体花性状聚类Fig.2 Flower traits clustering of C.duclouxii populations
3 讨 论
滇揪属梓树属树种,该属植物大多“自花不孕”。滇楸即使能结实,其种子活力低,发发芽率很低。在大部分地区花而不实,从而限制了这一树种的发展。主要采用埋根、扦插繁殖。为了解滇楸花的繁育、结实特性,特展其花部特性的调查研究。通过贵州滇楸资源主要分布地17个种群花部的7个表型性状调查研究,表型性状方差分析显示各性状在种群间与种群内差异均达到极显著水平,表明滇楸花的表型遗传多样性高,在种群内、种群间性状均存在丰富的遗传与变异[22-26]。滇楸种群间表型多样性小于种群内,说明优良单株选择的有效性。
对滇楸种群与地理位置关系的相关分析,随着海拔的升高,花枝有越来越短的趋势,花朵的上萼、下萼长度和花径(花冠口)宽度随着纬度的升高而变得越来越大,随着纬度由南向北的移动和海拔的变化,引起环境温度和湿度、昼夜长短变化,从而影响花朵的数量和花朵的大小。
根据聚类结果,滇楸种群花的表型变异并没有依地理距离而聚类,贵定城关、德新种群与新巴、元山种群被分别分为第一类和第二类。遵义乌江种群为第三类,该种群花上下萼长在所有种群中最长,花序较短;分布于西南部兴仁种群为第四类,花较小,花序长、花上萼、下萼较短,是适应昆虫访花,且开花期气候干燥,亦有利于授粉,环境条件对滇楸的表型变异具有很大的影响,是遗传因素与环境因子共同作用的结果,在一定程度上反映了种质的遗传多样性[15]。结果表明滇楸种群间和种群内的表型性状变异特别丰富,这些变异是自身遗传因素、地理隔离造成的遗传差异和环境因素共同作用的结果,反映了群体遗传稳定性与环境复杂性的互作关系及其适应环境压力的广泛程度。
在调查中发现 ,在立地条件相对一致的小范围内滇楸花有丰富的变异 。虽然对滇楸花的变异机理没有进行深入的研究,但至少可以说明,滇楸在长期的生长和演化过程中,对不同环境形成了不同的适应机制,从而产生了从个体到种群水平的不同变异现象。
[1] 潘庆凯,康平生,郭 明.楸树[M].北京:中国林业出版社,1991,37-38.
[2] 樊汝汶,吴琼美,邹惠渝.滇楸的胚胎学研究[J].南京林产工业学院学报,1980,2(2):67-80.
[3] 余丽云.滇楸的染色体核型分析[J].西南林学院学报,1991,11(1):29-32.
[4] 贾继文,麻文俊,王军辉,等.几种梓属植物花粉形态及分类学意义[J].林业科学,2012,48(7):182-185.
[5] 贾继文,王军辉,张金凤.梓树属花粉生活力的研究[J].西北植物学报,2009, 29(5):945-950.
[6] 王改萍,彭方仁,徐 涛,等.几种不同楸树花粉萌发率的测定及花粉超低温保存方法[J].南京林业大学学报:自然科学版,2008,32(5):123-126.
[7] 贾继文,王军辉,张金凤.楸树与滇楸种间杂交的初步研究[J].林业科学研究,2010, 23(3):382-386.
[8] 杨安敏,姚淑均,许 杰,等.滇楸半同胞家系子代苗期性状的遗传变异[J].种子,2010, 29(3):89-90.
[9] 赵 坤, 吴际友, 程 勇,等.楸树无性系嫩枝扦插繁殖的研究[J].中南林业科技大学学报, 2010, 13(7):66-69.
[10] 赵秋玲,王军辉,马建伟.梓树属植物叶片的气孔特征[J].东北林业大学学报,2011, 39(8):21-24.
[11] 邸 葆,孟 昱.抗寒锻炼期间三种楸树叶片激素含量变化[J].北方园艺,2012,13:89-91.
[12] 王新建,何 威,杨淑红, 等. 干旱胁迫下4种楸树嫁接苗叶绿素含量的变化[J]. 经济林研究,2008, 26(1):20-24.
[13] 卢晓强.黔中喀斯特山地8种树种早期生长和叶片养分动态的研究[J].南京林业大学学报:自然科学版,2008,32(1):38-42.
[14] 徐利霞,杨水平,姚小华,等.石漠化地区3个树种幼苗在水分胁迫下的光合特性与抗旱性关系[J].林业科学研究, 2006,19(6): 785-790.
[15] 文亚峰, 韩文军, 吴 顺.植物遗传多样性及其影响因素[J].中南林业科技大学学报, 2010, 30(12):80-87.
[16] 李 斌,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181-188.
[17] 葛 颂,王明庥,陈岳武.用同工酶研究马尾松群体的遗传结构[J].林业科学,1998,24(4 ):399-409.
[18] KHALIL M A K. Genetics of cone morphology of black spruce in Newfoundlan[J]. Canada Sil Genet,1984,33:101-109.
[19] 郭 宁,杨树华,葛维亚,等.新疆天山山脉地区疏花蔷薇天然居群表型多样性分析[J].园艺学报,2011,38(3):495–502.
[20] GROSS C L, BARTIER F V, MULLIGAN D R. Floral srucure,breeding sysem and frui-seed in he hrea ened sub-shrub Tetratheca juncea Smith (Tre-mandraceae) [J]. Annals of Botany,2003, 92(6):771-777.
[21] ECKHART V M. The effecs of floral display on pollina or visiaion vary among populaions of Phac-elia linearis (Hydro phy llaceae)[J] . Evolutionary Ecology, 1991, 5(4): 370-384.
[22] 柳新红,李因刚,赵 勋,等.白花树天然群体表型多样性研究[J].林业科学研究,2011,24(6):694-700.
[23] 陈香波,叶文国,田 旗,等.夏蜡梅天然群体表型变异及分布特征[J].北京林业大学学报, 2010,32(2):133-140.
[24] Zeneli G, Kola H, Dida M. Phenotypic variation in native walnut populations of Northern Albania[J].Scientia Horticulturae, 2005,105: 91-100.
[25] Putenikhin V P. Phenotype analysis of Picea obovata in the southern urals population structure[J].Lesovedenie,1997,6:37-49.
[26] 顾婧婧,金则新,熊 能.濒危植物夏蜡梅花的形态变异[J].植物研究,2010, 30(4): 461-467.
Study on phenotypic diversity of fl ower characters in Catalpa fargesii f.duclouxii
YAO Shu-jun1,2, ZHANG Shou-gong1, WANG Jun-hui1, ZHANG Nai-chun3, ZHANG Ming-gang2
(1. State Key Lab. of Tree Genetics and Breeding, Research Institute of Forestry, Chinese Academy of Forestry; Key Lab. of Tree Breeding and Cultivation of State Forestry Administration, Beijing 100091, China; 2. Guizhou Academy of Forestry, Guiyang 550005,Guizhou, China; 3. Forestry Seedling Station of Guizhou Provine, Guiyang 550001, Guizhou, China)
The phenotypic variations of 17 populations of Catalpa fargesii f. duclouxii, theirs 7 phenotypic traits including fl owers branch length, upper calyx length, lower calyx length, corolla and fl ower diameter were investigated. The results show that there existed notable variations of phenotypic characters of fl owers within and among the groups, the differences of the 7 phenotypic traits reached an extremely signif i cant level (P<0.01) , the mean variation coeff i cient was 25.94%, the amplitude of variations of each characteristic of fl owers ranged from 7.85%~47.17%, the mean phenotypic differentiation coeff i cients was 27.95%(14.92~ 42.15%), the variation within seventeen populations (72. 05%) was larger than that of among populations (27.95 % ); the fl ower traits were highly signif i cantly related with the latitude and longitude, and extreme signif i cantly with the altitude. According to the UPGMA cluster analysis, the 17 populations of C. fargesii f. duclouxii were divided into four groups.
Catalpa fargesii f.duclouxii; fl owers; morphological character; phenotypic character
S718.46
A
1673-923X(2013)10-0019-06
2013-05-16
“十二五”国家科技支撑计划(2012BAD21B03);贵州省种质资源清查保护项目之一“乡土树种滇楸、香椿种质清查及优树选择”(黔林种站字2010第39号)
姚淑均(1970-),女,贵州遵义人,博士研究生,主要从事林木遗传改良方面的研究; E-mail:197ysj@163.com
张守攻(1957-),男,安徽怀远人,研究员,博士生导师,主要研究方向:中国森林资源可持续经营的理论与技术、林木育种及森林资源培育研究;E-mail:shougong.zhang@caf.ac.cn
[本文编校:文凤鸣]
