钦州湾不同立地类型红树林的生长评价
2013-12-27何琴飞申文辉黄小荣谭一波
何琴飞,申文辉,黄小荣,刘 秀,谭一波
(1.广西林业科学研究院,广西 南宁 530002;2.国家林业局中南速生材繁育实验室,广西 南宁 530002;3.广西优良用材林资源培育重点实验室,广西 南宁 530002)
钦州湾不同立地类型红树林的生长评价
何琴飞1,2,3,申文辉1,2,3,黄小荣1,2,3,刘 秀1,2,3,谭一波1,2,3
(1.广西林业科学研究院,广西 南宁 530002;2.国家林业局中南速生材繁育实验室,广西 南宁 530002;3.广西优良用材林资源培育重点实验室,广西 南宁 530002)
在不同立地类型设定样方进行调查,对比分析了钦州湾14个立地类型红树林的生长情况,同时测定了立地土壤有机质、速效N等7个因子,进行主成分分析、聚类分析、相关分析和逐步回归分析。结果表明:(1)不同立地类型土壤因子不同,影响红树林生长的主要土壤因子是全盐量、速效N和pH值;(2)不同立地类型红树林的生长不同,种类、及群落覆盖度和更新层也存在一定差异。无瓣海桑在类型3中生长较好,秋茄在类型13中生长较好,桐花树在类型1、类型5、类型11、类型13和类型14中生长良好,白骨壤在类型11、类型13和类型14中生长好;(3)红树林生长指标与部分土壤因子之间存在显著的相关性;(4)桐花树树高与有机质,地径与土壤密度之间,白骨壤树高与土壤密度,地径与土壤密度、速效K、全盐量,以及冠幅与土壤密度之间,建立的回归方程均达到显著水平。
红树林;立地类型;土壤因子;生长评价
红树林是在热带、亚热带地区,陆地与海洋交界的海岸潮间带滩涂上生长的由木本植物组成的乔木和灌木林[1]。在消浪护岸、防治灾害、维持生物多样性和海岸带生态平衡、防治污染、净化水体、优化沿海环境和科学研究等方面发挥着重要作用,是全球海洋生物多样性保护的一个重要对象。红树林生长于常受海水浸淹、缺氧、多盐的潮滩上,生态环境和土壤基质影响着红树林植物的生态分布[2]。而沉积物及其上发育的土壤是直接影响红树植物生长发育最重要的环境因素之一[3]。Eisma等曾评价沉积物类型和土壤性质与红树林树种和生长状况之间的关系为“仍然不清楚”,只是粗略的和定性的[4]。大量学者在红树林分布[5-6]、群落特征[7]、空间结构[8-9]、宜林因子[10]、群落变化及其环境关系[11]、群落健康评价[12]、生态系统健康评价[13]等,但关于土壤因子直接对红树林生长的影响以及他们之间的相互关系的研究甚少。蒋燚等[14]根据海水盐度、潮滩位置、土壤质地将钦州湾沿海红树林宜林滩涂划分为3个立地区、9个立地组和28个立地类型(包含1个特殊杂草立地类型)。在此基础上调查了钦州湾不同立地类型红树林的生长情况,包括树高、胸径/地径、冠幅和从数等生长指标,并做对比分析和评价,同时测定立地土壤因子,找出影响红树林生长的主要因子,以便探讨红树林生长与土壤因子之间的关系,旨在为北部湾沿海滩涂适地适树造林、建设高效防护林体系提供科学依据。
1 研究方法
1.1 实验地概况
钦州湾位于北部湾顶部,广西海岸中段,在 21°33′20″~ 21°54′30″N,108°28′20″~108°45′30″E之间,属南亚热带海洋季风气候,年平均温度21~23 ℃。该湾由内湾(茅尾海)、湾颈和外湾(狭义上的钦州湾)三部分组成,中间狭窄,岛屿众多,两端开阔,呈哑铃状,东、西、北三面为陆地环绕,南面与北部湾相通,北面有钦江、茅岭江注入,是一个半封闭的天然河口湾。该湾口门宽29 km,纵深39 km,海岸线长达336 km,总面积380 km2[15]。整个钦州湾红树林岸线长约100 km,主要分布于茅尾海北部、西北部和金鼓江沿岸,在湾中部龙门群岛呈间断分布[16]。
1.2 样方设定
在不同立地类型设立10 m×10 m的样方,共调查了21个样方(同一立地类型的不同红树林群落分别调查),对样方内的植株逐株测定树高、胸径/地径、冠幅,同时调查群落类型、郁闭度与更新层情况,更新层设立1 m×1 m的样方,重复3次。
1.3 土壤取样和测定方法
土壤取样:在每个样方内采集0~30 cm的土层,混合取样,重复3次,分别装袋。
测定方法:测定有机质、速效N、速效P、速效K、全盐量、pH值、密度等7个指标。有机质用重铬酸钾氧化-外加热法,总N用凯氏蒸馏法,速效P用0.05 mol/L HCl—0.025 mol/L H2SO4浸提法,速效K用1 mol/L乙酸铵浸提—火焰光度法,全盐量用电导法,pH值用酸度计法,密度和土壤饱和持水量用烘干法。
1.4 数据处理
采用SPPS18.0软件进行单因素方差分析(One-Way ANOVA)、多重比较(Duncan)、主成分分析、样品聚类分析、相关分析(Two-tailed双尾检验)和逐步回归分析。
2 结果与分析
2.1 立地基本情况
调查的立地具体情况见表1。海水盐度:盐度>25为高盐、盐度10~25为中盐,分别用代号Ⅰ、Ⅱ表示;潮滩位置:外滩(高潮滩)、中滩(中潮滩)、内滩(低潮滩),分别用A、B、C表示;土壤质地:泥质、沙泥质、沙质,分别用1、2、3表示,含杂草用T表示。不同立地类型红树林群落存在差异,在钦州湾康熙岭、犀牛脚和茅尾海保护区(仙岛湖公园)的调查中发现,中盐区以桐花树Aegiceras corniculatum和无瓣海桑Sonneratia apertala为主,而高盐区以白骨壤Avicennia marina、桐花树和秋茄Kandelia candel为主,杂草立地出现在中盐区。不同类型的红树林覆盖度不同,其中,沙质土的类型2和类型10的覆盖度均低于0.50,沙泥质土的类型8和类型12也较低,均为0.55。大部分立地类型均有更新层,除了类型11和类型1中无瓣海桑-桐花树群落,更新层以桐花树、白骨壤和秋茄为主,其数量、苗高和地径在不同立地类型中亦不同。
可见,立地类型对红树林群落类型、覆盖度、幼苗更新层均有一定影响。
2.2 不同立地类型的土壤因子
不同立地类型土壤因子明显不同,养分含量不同。土壤有机质、速效N、速效P、速效K、全盐量、pH值、密度经单因素方差分析都有显著差异,结果见表2。有机质、速效P和全盐量含量均是类型13最高,都与其他类型存在显著差异,有机质含量最低的是类型12,只与类型9和类型10之间不存在显著差异;而速效P含量最低的是类型7;全盐量含量最低的是类型6,高盐区的5个类型比中盐区的类型(除类型1和类型4)的全盐量含量高。速效N含量最高的是类型4,其次是类型13、类型11和类型5,这4个类型之间都不存在显著的差异,含量最低的是类型9。速效K含量波动很大,最高的是类型14,高达798.2 mg/kg,其次是类型11、类型13和类型4,他们之间也不存在显著差异,类型6的含量最低,仅6.0 mg/kg。pH值类型9、类型10、类型12和类型14均在7.0以上,而其他类型的土壤均呈酸性。密度类型6最大,与其他类型均存在显著差异。类型13最小,只与类型4和类型11没有显著差异。† 不同字母表示处理组间差异显著(P<0.05);下同。
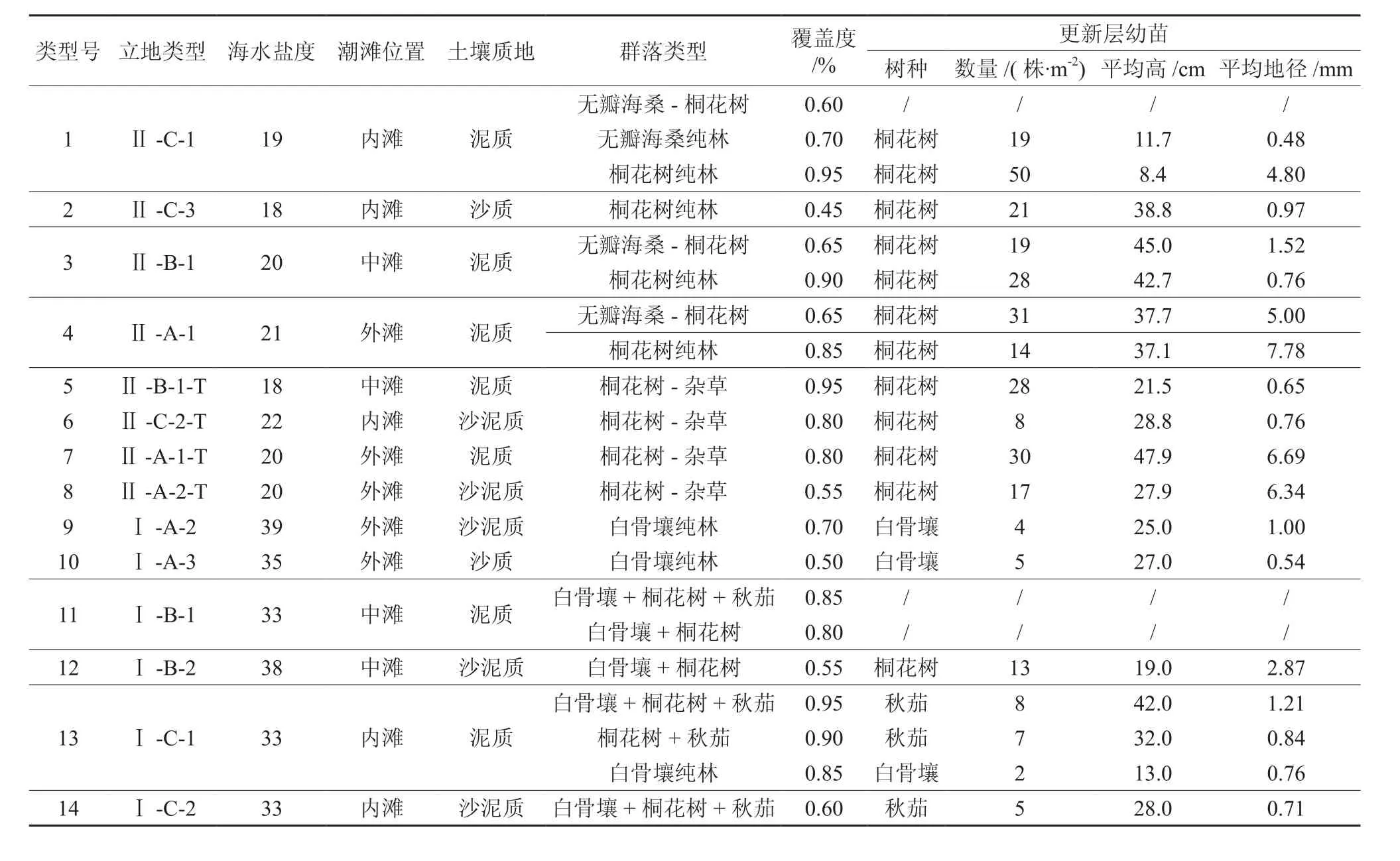
表1 立地基本情况Table 1 General situation of sites

表2 不同立地类型土壤因子†Table 2 Soil factors of different site-types
对8个土壤因子进行主成分分析,以累积贡献率大于80%的原则选择主成分,提取出对解释原有变量贡献最大的两个主成分,结果见表3。第一个主成分的特征值是4.445,方差贡献率为55.563%,第二个主成分的特征值是2.048,贡献率25.601%,累计贡献率达到81.164%(>80%),故前2个主成分是影响红树林生长状况的主要指标。

表3 土壤因子主成分分析Table 3 Analysis of principal components of soil factors
由主成分的特征向量(表4)可知,第一主成分中全盐量的特征向量值为0.920,速效N的特征向量值为0.876,第二主成分中pH值的特征向量值为0.926。故影响红树林生长的主要土壤因子为全盐量、速效N和pH值。

表4 土壤因子主成分的特征向量Table 4 Component matrix of soil factors
2.3 不同立地类型红树林的生长指标
在调查的14个立地类型中,主要是无瓣海桑、秋茄、桐花树和白骨壤,这4个树种在不同立地类型中的生长情况见表5。无瓣海桑在3个类型之间树高差异并不明显,而类型1的胸径和冠幅均明显小于类型3和类型4。秋茄在类型11、类型13和类型14这3个类型中地径不存在显著差异,但类型14的树高显著小于类型11和类型13,冠幅则是类型11明显小于类型13和类型14。桐花树的分布范围较广,共有12个立地类型,经方差分析可知,不同立地类型对桐花树的树高、地径、冠幅和从数均有明显影响,其中类型1的树高最高,为2.83 m,且与其他类型存在显著差异,其次较高的是类型13和类型11,均在2.0 m以上,而类型12的树高最小,仅为0.57 m;类型1的地径也最大,除与类型5、类型11和类型13没有显著差异外,与其他类型均存在显著差异;类型14的冠幅最大,与其他类型都存在显著差异,类型7的冠幅最小;类型8的从数最多,类型7的从数最少。白骨壤分布的5个立地类型中,树高大小为类型13>类型11>类型14>类型9>类型12>类型10,类型13与其他类型均存在显著差异;类型13的地径最大,但与类型11和类型14不存在显著差异,与类型9、类型10和类型12存在显著差异;类型14的冠幅最大,但与类型11和类型13也不存在显著差异,类型12的冠幅最小,只有0.84 m。
2.4 红树林生长立地类型的聚类分析
无瓣海桑在调查的3个类型中以类型3生长最好,秋茄在调查的3个类型中以类型13生长较好。对桐花树和白骨壤生长的立地类型进行聚类分析,结果如图1和图2所示,桐花的12个生长立地类型按4类分,把类型7单独分为一类;生长最差,类型4和类型12为一类,生长较差;类型2、类型3、类型6和类型8为一类;生长一般,类型1、类型5、类型11、类型13和类型14为一类,生长良好。白骨壤可分为2类,类型9、类型10和类型12为一类,生长较差;类型11、类型13和类型14为一类,生长好。

表5 不同立地类型红树林生长指标Table 5 Mangrove growth indexes of different site-types
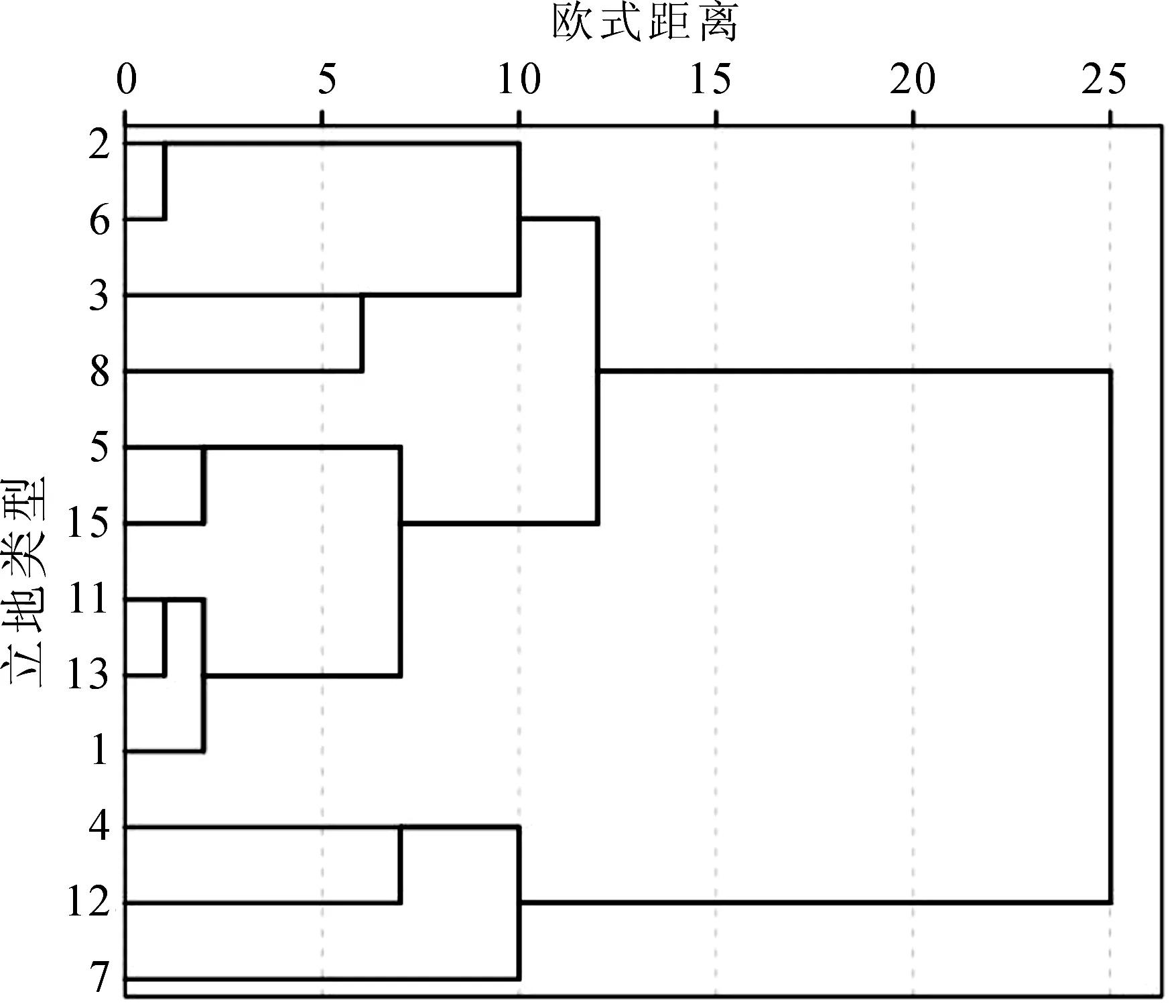
图1 桐花树生长立地类型聚类分析Fig.1 Dendrogram of cluster analysis about types of A. corniculatum
2.5 红树林生长指标与土壤因子的相关性
桐花树生长指标与土壤因子的相关性见表6。桐花树树高与有机质和速效N存在显著的正相关性,与密度存在显著的负相关性;地径与密度存在显著的负相关性,与饱和持水量存在显著的正相关性;而冠幅和从数均与土壤因子之间均不存在显著的相关性。

图2 白骨壤生长立地类型聚类分析Fig.2 Dendrogram of cluster analysis about types of A. marina
白骨壤生长指标与土壤因子的相关性见表7。白骨壤树高与速效N和全盐量存在极显著的正相关性,与有机质和饱和持水量存在显著的正相关性,与pH值存在显著的负相关性,与密度存在极显著的负相关性;地径与速效N存在极显著的相关性,与速效K和全盐量存在显著的正相关性,与密度存在极显著的负相关性;冠幅与密度存在显著的负相关性。

表6 桐花树生长指标与土壤因子的相关性†Table 6 Correlation between growth indexes and soil factors for A. corniculatum

表7 白骨壤生长指标与土壤因子的相关性Table 7 Correlation between growth indexes and soil factors for A. marina
2.6 红树林生长指标与土壤因子的逐步回归分析
逐步回归方程能建立最优回归方程,使多元线性回归方程显著,各个偏回归系数也显著。
桐花树树高(y1)与有机质(x1)红树林生长指标与土壤因子建立的回归方程如下:y1=0.577+0.036x1,R=0.648,P=0.023 < 0.05;
桐花树地径(y2)与土壤密度(x2):y2=9.341-5.209x2,R=0.600,P=0.039 < 0.05;
白骨壤树高(y3)与土壤密度(x2):y3=8.239-6.295x2,R=0.975,P=0.001 < 0.01;
白骨壤地径(y4)与土壤密度(x2)、速效K(x3)和 全 盐 量(x4):y=19.295-14.859x2+0.004x3-0.057x4,R=1.000,P=0.000 < 0.01;
白骨壤冠幅(y5)与土壤密度(x2):y5=6.077-4.112x2,R=0.867,P=0.025 < 0.05。
3 讨论与结论
(1)不同立地类型土壤因子不同,土壤有机质、速效N、速效K、全盐量、pH值和密度均有显著性差异,土壤全盐量、速效N和pH值是影响红树林生长的主要因子。红树林通过旺盛的生物累积和循环、强烈的生物积盐和严重的酸化作用等,使其生长的基质土镶的理化性状受到较大影响[17]。红树林发达的根系,能有效的网罗有机碎屑,固定和沉积污染物,使土壤N、P、K等含量增多[18]。
(2)不同立地类型红树林的生长不同,种类、及群落覆盖度和更新层也存在一定差异。无瓣海桑在类型3中生长较好;秋茄在类型13中生长较好;桐花树在类型1、类型5、类型11、类型13和类型14中生长良好,在类型7、类型4和类型12中生长较差;白骨壤在类型11、类型13和类型14中生长好,在类型9、类型10和类型12中生长较差。在几乎所有的植物群落中,养分供应是一个影响群落结构的重要的驱动变量[19],红树林也是如此。红树植物大多数表现出对养分供应变化非常敏感[20-21]。养分供应的增加,大多导致更高的生长率[22]。白骨壤属的树种具有速生和承受土壤高盐度的能力,因此,白骨壤在高盐度和高营养水平时生长会占优势[19]。但对增加养分的反应,依环境条件而异,并且取决于物种的特性[23]。总体而言,红树林生长较好的立地类型为土壤养分较高的内、中滩泥质或沙泥质土。早在我国20世纪60年代琼北红树林海岸研究就已发现红树林可以生长在各种底质的海岸上,以淤泥质潮滩最普遍和生长最好[24]。
(3)红树林生长指标与部分土壤因子之间存在显著的相关性。红树林与土壤之间的作用就是相互的,土壤条件影响着红树林的类型、分布和生长,反过来,不同红树林也对土壤产生不同影响[17]。
(4)桐花树的树高与有机质、地径与土壤密度之间的回归方程均达到显著,但复相关系数R不是太高,白骨壤树高与土壤密度、地径与土壤密度、速效K、全盐量,以及冠幅与土壤密度之间的回归方程均达到显著,且复相关系数都很高。本文建立的红树林生长指标与土壤因子之间的回归方程在一定程度上反映了其相互关系,但调查点仅限于钦州湾立地,立地类型有限,存在一定的局限性。
致谢:感谢钦州市林科所蔡林、钟文基和樊东涵在野外调查工作中给予帮助。
[1] 范航清.红树林海岸环保卫士[M].南宁:广西科学技术出版社 ,2000:7-8.
[2] 李信贤,温远光,温肇穆.广西海滩红树林主要建群种的生态分布和造林布局[J].广西农学院学报,1991,10(4):82-89.
[3] Snedaker S C,Snedaker J G eds.郑德璋,郑松发,廖宝文,译校.红树林生态系统研究方法[M].广州:广东科技出版社,1994:79-134.
[4] Eisma D.Intertidal Deposits, River Mouths, Tidal Flats and Coastal Lagoon[M].Boca Raton: CRC Press,1998:415-432.
[5] 石 莉.中国红树林的分布状况、生长环境及其环境适应性[J].海洋信息 ,2002,(4):14-18.
[6] 李春干.广西红树林资源的分布特点和林分结构特征[J].南京林业大学学报:自然科学版,2003,27(5):15-19.
[7] 范航清,黎广钊.海堤对广西沿海红树林的数量、群落特征和恢复的影响[J].应用生态学报,1997,(3):240-244.
[8] 李际平,张宏伟,李建军,等.不同潮带红树林空间结构比较研究[J].中南林业科技大学学报,2010,30(1):8-11.
[9] Ellisw L, Bell S S. Canopy gaps formed by mangrove trimming:an experimental test of impact on litter fall and standing litter stock in Southwest Florida (USA)[J]. Journal of Experimental Marine Biology and Ecology, 2004,311:201-222.
[10] 刘 亮,范航清.红树林宜林因子研究[J].湿地科学与管理,2010, 6(2):57-60.
[11] 王丽荣,李 贞,蒲杨婕,等.近50年海南岛红树林群落的变化及其与环境关系分析——以东寨港、三亚河和青梅港红树林自然保护区为例[J].热带地理,2010,30(2):115-120.
[12] 王丽荣,李 贞,蒲杨婕,等.海南东寨港、三亚河和青梅港红树林群落健康评价[J].热带海洋学报,2011,30(2):81-86.
[13] 王树功,郑耀辉,彭逸生,等.珠江口淇澳岛红树林湿地生态系统健康评价[J].应用生态学报,2010,21(2):391-398.
[14] 蒋 燚,何琴飞,刘 秀,等.钦州湾沿海宜林滩涂立地类型划分[J].林业实用技术,2011,113(5):21-23.
[15] 李树华,黎广钊.中国海湾志第十二分册(广西海湾) [M].北京:海洋出版社,1993:144-216.
[16] 邓朝亮,刘敬合,黎广钊,等.钦州湾海岸地貌类型及其开发利用自然条件评价[J].广西科学院学报,2004,20(3):174-178.
[17] 蓝福生,莫权辉,陈 平,等.广西海滩红树林于土壤的关系[J].广西植物 ,1994,14(4):54-59.
[18] 陈桂珠.研究保护和开发利用红树林生态系统[J].生态科学,1991,(11):116-119.
[19] Lovelock C E, Feller I C. Photosynthetic performance and resource utilization of two mangrove species coexisting in hypersaline scrub forest[J]. Oecologia, 2003,134:455-462.
[20] Yates E J,Ashwath N, Midmore D J. Responses to nitrogen,phosphorus, potassium and sodium chloride by three mangrove species in pot culture. Trees Struct. Funct.,2002,16:120-125.
[21] Naidoo G. Factors contributing to dwarfing in the mangrove Avicennia marina[J]. Ann. Bot., 2006,97:1095-1101.
[22] Lovelock C E, Feller I C, McKee K L, et al. The effect of nutrient enrichment on growth, photosynthesis and hydraulic conductance of dwarf mangroves in Panama[J]. Funct. Ecol.,2004,(18):25-33.
[23] Kraoss K W, Lovelock C E, Mckee K L, et al. Environmental drivers in mangrove establishment and early development: a review[J]. Aquatic Botany, 2008,89:105-127.
[24] 张乔民,隋淑珍,张叶春,等.红树林宜林海洋环境指标研究[J].生态学报 ,2001,21(9):1427-1437.
Evaluation of mangrove growth in different site types in Qinzhou Bay
HE Qin-fei1,2,3, SHEN Wen-hui1,2,3, HUANG Xiao-rong1,2,3, LIU Xiu1,2,3, TAN Yi-bo1,2,3
(1.Guangxi Forestry Research Institute, Nanning 53002, Guangxi, China; 2.Key Lab. of Central South Fast-growing Timber Cultivation under Forestry Ministry of China, Nanning 53002, Guangxi, China; 3.Guangxi Key Laboratory of Superior Timber Trees Resource and Cultivation, Nanning 53002, Guangxi, China)
Mangroves on different sites in Qinzhou Bay in Guangxi were investigated for understanding their growth in relation to sitetypes and soil factors (organic matter, available N, etc.) through principal component analysis, cluster analysis, correlation analysis and linear regression (stepwise), involving 14 site-types and 7 soil factors. The results show that (1) Soil factors varied dependent on the site-types, while salt content, available N and pH were the main soil factors that affected on the growth of mangraves; (2) Different sitetypes had specif i c predominant species, specif i c coverage and regeneration layers, with Sonneratia apertala well developed on sites of type 3, Kandelia candel on sites of type 13, Aegiceras corniculatum on type 1, 5, 11, 13 and 14 and Avicennia marina on type 11, 13 and 14 ; (3) The indexes of growth were signif i cantly correlated to part of the soil factors; (4) The regression equation between the height and the organic matter of A. corniculatum reached signif i cant level; as well as that between the basal diameter and the soil density of A.corniculatum; between the height and the soil density of A. marina; between the basal diameter and the soil density, the available K, the salt content of A. marina; and between the crown diameter and density of A. marina.
mangrove;site type;soil factor;growth evaluation
S796
A
1673-923X(2013)03-0057-07
2012-10-14
广西科技厅业务专项(林科201104号),国家科技支撑计划项目专题(2009BADB2B04-02)
何琴飞(1982-),女,湖南娄底人,工程师,硕士研究生,主要从事植物生态研究;E-mail:dragonf l y.hqf@126.com
[本文编校:吴 毅]
