桉-桤不同混合比例凋落物分解过程中土壤动物群落动态
2013-12-20李艳红杨万勤罗承德吴福忠
李艳红,杨万勤 ,罗承德,吴福忠,胡 杰
(1.四川农业大学生态林业研究所,生态林业工程重点实验室,成都 611130;2.西华师范大学西南野生动植物资源保护教育部重点实验室,南充 637009)
森林凋落物分解是地力维持的基础[1],但受到各种生物与非生物因素的综合影响[2-4]。土壤动物是凋落物分解的积极参与者[5-9],并受到凋落物种类和质量的影响,而且不同体径的土壤动物对凋落物分解的贡献也不相同[10-12]。同时,参与凋落物分解的土壤动物类群和数量也受到凋落物质量等生物与非生物因素的影响[13-14]。在凋落物分解的不同阶段,凋落物的分解程度差异为不同土壤动物提供食物来源,因而土壤动物群落结构可能发生相应变化[15-16]。不同凋落物混合提高了生境异质性,与单一凋落物相比土壤动物更丰富[17]。因此,研究不同质量的凋落物混合分解过程中,不同体径的土壤动物群落变化对于探讨混交林凋落物的分解机制有着重要的意义,也可能为人工混交林的构建提供一定的科学依据。
巨桉(Eucalyptus grandis)是南方工业原料林的主要造林树种之一,但单一的桉树人工林可能导致生物多样性降低和地力衰退等生态问题[18]。台湾桤木(Alnus formosana)是非豆科固氮的常绿速生树种,能加速低品质树种凋落物分解和养分循环[19],可能成为降低巨桉生态风险的伴生树种。伴生树种的栽植是否改变凋落物分解过程中土壤动物群落的结构和功能,加速凋落物的分解,从而影响人工林生态系统的物质循环,亟待深入研究。因此,本文研究了不同比例的巨桉与台湾桤木混合凋落物分解过程中的土壤动物群落动态,以期为人工林的可持续经营提供一定的理论依据。
1 材料与方法
1.1 研究区域与样地概况
研究区域位于四川省乐山市苏稽镇(N29°36'—29°37',E103°36'—103°37'),属中亚热带气候带,四季分明,雨量丰沛,年平均降水量多数在1000 mm以上,水热同季,年平均气温在16.5—18.0℃,大于或者等于0℃积温为6100—6500℃,大于或者等于10℃积温5269—5662℃,大于或者等于20℃积温2930—3376℃,年平均无霜期长达300 d以上。研究区域主要木本植物有多花野牡丹(Melastoma affine)、乌泡子(Rubus parkeri)、岗柃(Eurya groffii)、菝葜(Smilax china)等;主要草本植物有芒萁(Dicranopteris dichotoma)、芒(Miscanthus sinensis)、十字苔草(Carex cruciata)等。
1.2 试验方法
2009年4月,收取巨桉纯林、台湾桤木纯林的自然凋落叶,自然状态下风干,按照10∶0(TI)、7∶3(TII)5∶5(TIII)、3∶7(TIV)和0∶10(TV)等5个巨桉(Eucalyptus grandis)和台湾桤木(Alnus formosana)凋落物混合比例,将不同混合比例的凋落物10g分别放入3种不同孔径[20](6目、30目和260目)尼龙袋(20 cm×20 cm)中。其中,6目网袋允许各种土壤动物的作用;30目网袋则基本排除了大型土壤动物的影响,允许中小型土壤动物自由出入;260目(对照)网袋则尽可能排除土壤动物的作用。每种比例、每种规格的凋落物各54袋,共计810袋,4月底放置于相应比例桉-桤混交人工林下,其上覆盖少许土壤,以固定凋落物袋。当年5月至翌年4月取样。根据凋落物分解规律,前半年每半个月采集一次样品,后半年每月采集一次样品。每次采集不同孔径不同混合比例的凋落物袋各3袋,采用干漏斗法(Tullgren)和手捡法分离凋落物中的土壤动物,镜检计数分类,参照《中国土壤动物检索图鉴》[21]鉴定捕获的土壤动物。分离结束后清除叶片表面土壤颗粒和混入的杂物,65℃烘干后称量,计算质量损失率[12]。
1.3 数据处理与统计分析
土壤动物各类群数量等级划分:以个体数占总数10%以上者为优势类群,占总数1%—10%者为常见类群,占总数0.1%—1%为稀有类群,占总数0.1%以下为极稀有类群。
数据统计分析均采用SPSS16.0和Excel软件完成。经球形检验(Mauchly's Test of Sphericity),P<0.05,故采用一般线性模型(General Linear Model)的Repeated Measures对5种混合比例凋落物间土壤动物个体数、类群数进行重复测量方差分析,并用Multivariate过程实现对5种混合比例凋落物的两两比较。采用单因素方差分析(one-way ANOVA)比较3种网孔凋落物袋(260目、30目和6目)中凋落物分解速率差异及不同月份6目与30目中中小型土壤动物个体数和类群数差异。采用Pearson法检验质量损失率与土壤动物类群数和个体数的相关性,以及6目凋落物袋中大型土壤动物与中小型土壤动物的相关性。
2 结果与分析
2.1 土壤动物群落组成
在桉-桤5种不同混合比例、3种规格的810只凋落袋中共收集土壤动物75651只,隶属2门10纲20目(表1)。
其中大型土壤动物17类,占77.27%,中小型土壤动物5类,占22.73%。优势类群主要为中小型土壤动物中的弹尾目和蜱螨目,占97.24%;常见类群为大型土壤动物中的后孔寡毛目;稀有类群有蠋线纲、双尾目、啮虫目、鞘翅目(隐翅虫科)、鞘翅目幼虫、双翅目幼虫、膜翅目(蚁科)等7类;极稀有类群有蜘蛛目、裂盾目、倍足纲、地蜈蚣目、石蜈蚣目、综合纲、原尾纲、等翅目、同翅目、缨翅目、双翅目、鳞翅目幼虫等12类。5种不同混合比例的凋落物中,大型土壤动物的分布是TV最多,TI最少,混合凋落物介于二者之间,且随桤木比例的增加呈增多的趋势;而中小型土壤动物在TI中最多,而TV最少。大型土壤动物优势类群因凋落物的组成不同而有所变化,TI主要有后孔寡毛目(37.09%)、鞘翅目成虫(18.31%)、啮虫目(12.68%),TII主要为后孔寡毛目(40.17%)、双翅目幼虫(14.64%),TIII和TIV主要为后孔寡毛目,分别占58.39%和71.26%,TV主要为啮虫目(37.12%)、后孔寡毛目(33.04%)。啮虫目在TV中的数量远远高于其它四种凋落物,后孔寡毛目在混合凋落物中数量较高。中小型土壤动物的优势类群都是弹尾目和蜱螨目,在不同的凋落物中蜱螨目的个体数量在TIII和TIV凋落物中较为丰富,弹尾目的数量是TI>TII>TIII>TIV>TV,蠋线纲亦喜生活在TI凋落物中。凋落物土壤动物中,杂食性所占比例最大(50%),其次是捕食性(18.18%),植食性和腐食性各占13.64%,菌食性最少,占4.54%。杂食性动物在凋落物分解过程中自始至终存在,腐食性动物(后孔寡毛目)在6月中旬出现,7—8月最高,随后降低。
2.2 土壤动物数量变化

五种不同比例混合凋落物中的土壤动物个体数在260目凋落物袋中(P=0.033,F=4.071)差异显著,在30目(P=0.003,F=8.763)和6目(P=0.002,F=9.709)差异中极其显著。从图1可以看出,中小型土壤动物个体数量是260目>30目>6目,5—11月,中小型土壤动物个体数在30目与6目中差异显著(P<0.05)。中小型土壤动物个体数量随时间变化呈现上升、下降的动态(图1),个体数量在7—8月较高,最高可达到909只(图1)。2009年5月至8月,30目TV凋落物中的中小型土壤动物个体数量呈上升趋势(图1)。5月和7月,30目中5种不同混合比例的凋落物中中小型土壤动物个体数差异显著(P<0.05)。中小型土壤动物在TV凋落物中最少(图1)。6目中中小型土壤动物的个体数远高于大型土壤动物(图1),但二者相关性不显著(P=0.257,r=0.121)。大型土壤动物个体数在各月间波动较大(图1),7月初,TV凋落物中大型土壤动物个体数最高,为9只,与其它凋落物存在显著差异(P<0.05)。大型土壤动物的个体数量在TI中较少,低于混合凋落物。
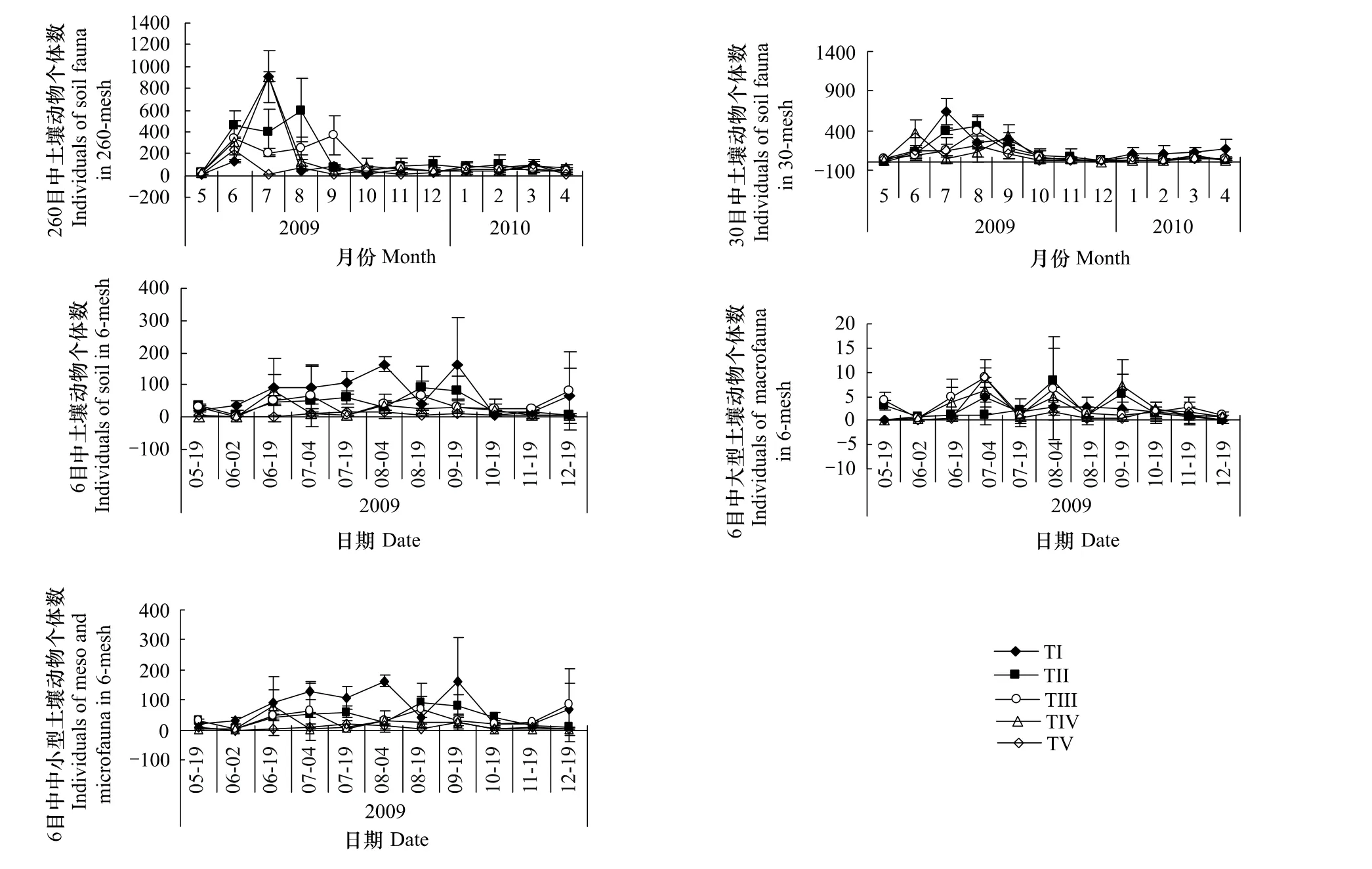
图1 不同网孔中5种不同混合比例的凋落物分解过程中土壤动物个体数量动态(平均值±标准差)Fig.1 Individuals dynamics of soil fauna in different mesh litterbags and mixing ratio litter during the decomposition of litter(mean±s.d)
2.3 土壤动物类群变化
5种不同比例混合凋落物中的土壤动物类群数在260目凋落物袋中差异不显著(P=0.380,F=1.172),在30目中差异显著(P=0.020,F=9.932),在6目中差异极其显著(P=0.000,F=13.450)。土壤动物类群数是30目>6目>260目,中小型土壤动物类群数在30目中最多,6目与30目凋落物袋中中小型土壤动物类群数仅在7月存在显著差异(P=0.017,F=6.472)。10月,中小型土壤动物类群数在TI凋落物中最高,达7类。TV中中小型土壤动物类群数较混合凋落物中少。6目中土壤动物类群数TV最低。TV中大型土壤动物类群数在8月初达到最高,为3类。6目中大型土壤动物与中小型土壤动物的类群数呈极其显著正相关(P=0,r=0.363)(图2)。
2.4 凋落物分解速率与土壤动物的关系

图2 不同网孔中5种不同混合比例的凋落物分解过程中土壤动物类群动态(平均值±标准差)Fig.2 Groups dynamics of soil fauna in different mesh litterbags and mixing ratio litter during the decomposition of litter(mean±s.d)
5—9月,260目、30目凋落物袋中5种不同混合比例的凋落物分解缓慢,10月份以后呈明显上升趋势(图3)。5种不同混合比例的凋落物在6目中的分解速率呈现TV>TIV>TIII>TII>TI(图3,c),且在6、7月分解速率差异显著(P<0.05)。TV和TIV迅速分解期在5月下旬至6月初,TIII和TII迅速分解期5月下旬至6月中旬,TI的迅速分解期分别在8月上旬,截至10月中旬,6目中凋落物基本分解完毕。6—8月,5种不同混合比例的凋落物的分解速率在6目与30目中存在显著差异(P<0.05)。
Pearson相关性检验表明,在260目凋落物袋中,质量损失率与类群数在TI(P=0.031,r=0.620)、TII(P=0.034,r=0.614)中相关性显著,在TV(P=0.003,r=0.785)中相关性极其显著。30目凋落物袋中,质量损失率与个体数在TII(P=0.001,r=0.828)中相关性极其显著,质量损失率与类群数在TV(P=0.022,r=0.649)中相关性显著。研究还发现,6目中质量损失率与大型土壤动物的个体数和类群数相关性均不显著。
3 结论和讨论
研究不同混合比例桉-桤凋落物的土壤动物群落组成及群落动态对于揭示凋落物的分解与土壤动物的关系,构建综合效应较高的人工林有着重要意义。本项研究显示,5种混合比例凋落物中的土壤动物个体数在3种网孔中(260目,30目和6目)均差异显著,类群数在30目和6目中差异显著,表明不同比例的凋落物混合可改变凋落物中土壤动物的数量和组成。中小型土壤动物弹尾目更喜生活在TI凋落物中,混合凋落物次之;TV凋落物中中小型土壤动物的个体数和类群数均较少;大型土壤动物的个体数是混合凋落物高于TI凋落物。综上表明在巨桉凋落物中混交台湾桤木虽降低了弹尾目等中小型土壤动物的数量,但在一定程度上可提高大型土壤动物的数量。凋落物分解过程中,杂食性土壤动物自始至终存在,腐食性土壤动物在凋落物分解的后期和末期最丰富,表明伴随凋落物的分解,土壤动物的组成和功能发生相应的改变。

图3 不同网孔中5种不同混合比例的凋落物的质量损失率(平均值±标准差)Fig.3 Mass loss rate of litter in 260-,30-,and 6-mesh(mean±s.d)
土壤动物个体数最高峰在7—8月份,类群数最高值在10月份。在大多数情况下,温度的升高可使微小型节肢动物密度增加[22-23],雨热同季可提高微生物的活动,为土壤动物的生存和繁衍提供了良好的微环境[24],因此高温潮湿环境中土壤动物数量丰富[25]。在凋落物分解早期,凋落物中含土壤动物所需的营养物质,土壤动物大量聚集和繁衍[26]。随着分解的持续进行,营养元素含量随凋落物失重量的增高而降低,难以分解的物质相对增多[27],土壤动物所需的养分骤减,土壤动物数量和类群呈下降趋势,同时在凋落物分解过程中,凋落物中的大型动物类群亦发生相应变化,杂食性土壤动物自始至终存在,腐食性动物分解后期和末期迅速增加,土壤动物在凋落物分解过程中的出现时序取决于其食性和凋落物的分解程度[12]。
大型土壤动物个体数和类群数在6—8月较高,这也是6目中凋落物分解最迅速的时期。6—8月,5种不同混合比例的凋落物的分解速率在6目与30目中存在显著差异(P<0.05),且6目快于30目[12]。6目允许所有土壤动物自由出入,而30目限制了大型土壤动物的活动,由此可推测,6目中活动的大型土壤动物加速了凋落物的分解。此外,TV中大型土壤动物数量高于TI凋落物,混合凋落物介于二者之间,而6目凋落物袋中,凋落物的分解速率亦是TV>混合凋落物>TI。再者,30目和6目中,TI凋落物中中小型土壤动物的数量均比TV丰富,但TI的分解速率仍慢于TV,同时,6目中的中小型土壤动物个体数远低于30目,但6目的分解速率却快于30目,由此可知,大型土壤动物对凋落物分解的贡献大于中小型土壤动物,因此,当巨桉纯林混栽台湾桤木,可提高凋落物中大型土壤动物的个体数,而大型土壤动物的活动可促进凋落物分解[16,28-29],养分的循环,这与Pozo等[19]的研究结果相一致,而Wise[5]认为大型土壤动物对新鲜凋落叶的分解作用不明显,仅加速陈旧凋落叶的分解,Garcia-Pausas等[30]则认为土壤微环境对凋落物分解的贡献远大于土壤动物的活动。在凋落物分解过程中,分解前期土壤动物贡献最大,分解后期则主要由微生物完成[31]。
研究表明,在巨桉纯林中混栽台湾桤木,可提高凋落物中大型土壤动物的个体数量,而大型土壤动物的活动可加速凋落物的分解和养分的循环。在研究中却发现6目中质量损失率与大型土壤动物的个体数和类群数相关性均不显著,原因在于部分大型土壤动物活动能力强、在凋落袋中停滞时间短,即摄食完毕则迅速离开,因而不易被采集和统计。另外,在260目仍采集到中小型土壤动物且数量较高[12]。
[1] Yang W Q,Deng R J,Zhang J.Forest litter decomposition and its responses to global climate change.Chinese Journal of Applied Ecology,2007,18(12):2889-2895.
[2] Tu M Z,Yao W H,Weng H,Li Z A.Characteristics of litter in evergreen broadleaved forest of the Dinghu mountain.Acta Pedologica Sinica,1993,30(1):34-42.
[3] Smith V C,Bradford M A.Litter quality impacts on grassland litter decomposition are differently dependent on soil fauna across time.Applied Soil Ecology,2003,24(2):197-203.
[4] Vossbrinck C R,Coleman D C,Woolley T A.Abiotic and biotic factors in litter decomposition in a semiarid grassland.Ecology,1979,60(2):265-271.
[5] Wise D H,Schaefer M.Decomposition of leaf litter in a mull beech forest:comparison between canopy and herbaceous species.Pedobiologia,1994,38:269-288.
[6] Ekschmitt K,Liu M Q,Vetter S,Fox O,Wolters V.Strategies used by soil biota to overcome soil organic matter stability-Why is dead organic matter left over in the soil?Geoderma,2005,128(1/2):167-176.
[7] Maraun M,Scheu S.Changes in microbial biomass,respiration and nutrient status of beech(Fagus sylvatica)leaf litter processed by millipedes(Glomeris marginata).Oecologia,1996,107(1):131-140.
[8] Seastedt T R.The role of microarthropods in decomposition and mineralization processes.Annual Review of Entomology,1984,29:25-46.
[9] Wang S J,Ruan H H,Wang B.Effects of soil microarthropods on plant litter decomposition across an elevation gradient in the Wuyi Mountains.Soil Biology and Biochemistry,2009,41(5):891-897.
[10] Zhong W Y,Yin X Q,Chen P.Relationship of litter decomposition and consumption with soil animals in Maoer Mountain forest.Chinese Journal of Applied Ecology,1999,10(4):511-512.
[11] Lin Y H,Zhang F D,Liu H D,Su H L.Fluctuation of soil fauna community in Baiwangshen during paper Mulberry leaf litter decomposition.Chinese Journal of Zoology,2005,40(3):60-66.
[12] Li Y H,Luo C D,Yang W Q,Hu J,Wu F Z.Decomposition of eucalyptus-alder mixed litters and dynamics of soil faunal community.Chinese Journal of Applied Ecology,2011,22(4):851-856.
[13] Lin Y H,Yang D F,Zhang F D,Wang J X,Bai X L,Wang B.Structure of soil animal community of oakery litter and fluctuation during leaf litter decomposition.Forest Research,2006,19(3):331-336.
[14] Li X F,Han S J,Guo Z L,Zheng X B,Song G Z,Li K X.Decomposition of pine needles and twigs on and under the litter layer in the natural Korean pine broadleaved forests.Journal of Beijing Forestry University,2006,28(3):8-13.
[15] Ke X,Zhao L J,Yin W Y.Succession in communities of soil animals during leaf litter decomposition in Cyclobalanopsis glauca forest.Zoological Research,1999,20(3):207-213.
[16] Irmler U.Changes in the fauna and its contribution to mass loss and N release during leaf litter decomposition in two deciduous forests.Pedobiologia,2000,44(2):105-118.
[17] Wu P F,Zhu B.Comparison of soil animal community characteristics between alder and cypress mixed plantation and cypress pure plantation.Chinese Journal of Applied and Environmental Biology,2008,14(4):488-493.
[18] Zhang J,Yang W Q.Ecosystem Researches on Eucalypt(Eucalyptus grandis)Short-Term Rotation Plantation.Chengdu:Sichuan Scientific and Technical Press,2008.4
[19] Pozo J,Basaguren A,Elósegui A,Molinero J,Fabre E,Chauvet E.Afforestation with Eucalyptus globulus and leaf litter decomposition in streams of northern Spain.Hydrobiologia,1998,373-374:101-110.
[20] Zhang X P,Zhang Y,Hou W L,Yin X Q.Decomposition of coniferous litter and the function of soil animal in the Xiao Hinggan Mountains.Scientia Geographica Sinica,2000,20(6):552-556.
[21] Yin W Y.Pictorial Keys to Soil Animals of China.Beijing:Science Press,1998.
[22] Kennedy A D.Simulated climate change:a field manipulation study of polar microarthropod community response to global warming.Ecography,1994,17(2):131-140.
[23] Harte J,Rawa A,Price V.Effects of manipulated soil microclimate on mesofaunal biomass and diversity.Soil Biology and Biochemistry,1996,28(3):313-322.
[24] Miao Y J,Yin X Q.Study on soil animals community of Pinus koraiensis broad-leaved mixed forest in Xiaoxing'an mountain.Scientia Silvae Sinicae,2005,41(2):204-209.
[25] González G,Seastedt T R.Comparison of the abundance and composition of litter fauna in tropical and subalpine forests.Pedobiologia,2000,44(5):545-555.
[26] Wang S J,Ruan H H,Wang J S,Xu Z K,Wu Y Y.Dynamic Change of soil fauna community structure in the course of litter decomposition on the Wuyi Mountains.Journal of Southwest Forestry University,2010,30(6)43-47.
[27] Shao Y Q,Zhao J,Yang J.Studies on decomposing of litters and characteristic of nutritious elements in inner Mongolia huangfuchuan watershed.Journal of Soil and Water Conservation,2004,18(3):81-84.
[28] Hättenschwiler S,Gasser P.Soil animals alter plant litter diversity effects on decomposition.Proceedings of the National Academy of Sciences of the United States of America,USA,2005,102(5):1519-1524.
[29] Bradford M A,Tordoff G M,Eggers T,Jones T H,Newington J E.Microbiota,fauna,and mesh size interactions in litter decomposition.Oikos,2002,99(2):317-323.
[30] Garcia-Pausas J,Casals P,Romanyà J.Litter decomposition and faunal activity in Mediterranean forest soils:effects of N content and the moss layer.Soil Biology and Biochemistry,2004,36(6):989-997.
[31] Li Z A,Zou B,Ding Y Z,Cao Y S.Key factors of forest litter decomposition and research progress.Chinese Journal of Ecology,2004,23(6):77-83.
参考文献:
[1] 杨万勤,邓仁菊,张健.森林凋落物分解及其对全球气候变化的响应.应用生态学报,2007,18(12):2889-2895.
[2] 屠梦照,姚文华,翁轰,李志安.鼎湖山南亚热带常绿阔叶林凋落物的特征.土壤学报,1993,30(1):34-42.
[10] 仲伟彦,殷秀琴,陈鹏.帽儿山森林落叶分解消耗与土壤动物关系的研究.应用生态学报,1999,10(4):511-512.
[11] 林英华,张夫道,刘海东,苏化龙.百望山土壤动物群落结构在枸树落叶分解中的变化.动物学杂志,2005,40(3):60-66.
[12] 李艳红,罗承德,杨万勤,胡杰,吴福忠.桉-桤混合凋落物分解及其土壤动物群落动态.应用生态学报,2011,22(4):851-856.
[13] 林英华,杨德付,张夫道,王建修,白秀兰,王兵.栎林凋落层土壤动物群落结构及其在凋落物分解中的变化.林业科学研究,2006,19(3):331-336.
[14] 李雪峰,韩士杰,郭忠玲,郑兴波,宋国正,李考学.红松阔叶林内凋落物表层与底层红松枝叶的分解动态.北京林业大学学报,2006,28(3):8-13.
[15] 柯欣,赵立军,尹文英.青冈林土壤动物群落结构在落叶分解过程中的演替变化.动物学研究,1999,20(3):207-213.
[17] 吴鹏飞,朱波.桤柏混交林与纯柏林土壤动物群落特征的比较.应用与环境生物学报,2008,14(4):488-493.
[18] 张健,杨万勤.短轮伐期巨桉人工林生态系统.成都:四川科学技术出版社,2008.4
[20] 张雪萍,张毅,侯威岭,殷秀琴.小兴安岭针叶凋落物的分解与土壤动物的作用.地理科学,2000,20(6):552-556.
[21] 尹文英.中国土壤动物检索图鉴.北京:科学出版社,1998.
[24] 苗雅杰,殷秀琴.小兴安岭红松阔叶混交林土壤动物群落研究.林业科学,2005,41(2):204-209.
[26] 王邵军,阮宏华,汪家社,徐自坤,吴焰玉.武夷山土壤动物群落结构在凋落物分解过程中的变化.西南林学院学报,2010,30(6)43-47
[27] 邵玉琴,赵吉,杨劼.内蒙古皇甫川流域凋落物分解过程中营养元素的变化特征.水土保持学报,2004,18(3):81-84.
[31] 李志安,邹碧,丁永祯,曹裕松.森林凋落物分解重要影响因子及其研究进展.生态学杂志,2004,23(6):77-83.
