基于MAXENT模型的贺兰山岩羊生境适宜性评价
2013-12-20刘振生滕丽微王晓勤孔芳毅
刘振生,高 惠, 滕丽微,*,苏 云,王晓勤,孔芳毅
(1.东北林业大学野生动物资源学院,哈尔滨 150040;2.国家林业局野生动物保护学重点开放实验室,哈尔滨 150040;3.内蒙古贺兰山国家级自然保护区管理局,巴彦浩特 750300)
生境为野生动物提供了生存、繁衍及种群发展所必须的资源[1],生境适宜度是指生境能支持一个特定物种的潜在能力[2],是影响野生动物生存和发展的重要因素,对物种进行生境评价是对野生动物进行有效保护和管理的第一步,并且能够为相关部门制定有效的物种保护对策提供科学依据。
岩羊(Pseudois nayaur)属偶蹄目、牛科、羊亚科、岩羊属,是青藏高原的特有种,也是青藏高原分布最广的有蹄类之一,被世界自然保护联盟(IUCN)收录为低危种[3],在我国被列为国家Ⅱ级重点保护野生动物[4]。国内主要分布于西藏、云南、四川、内蒙古、新疆、甘肃、青海、宁夏和陕西等省区[5]。目前,专家学者已对其行为生态学[6-7]、食性[8]、种群动态[9-10]和家域[11]等方面做了大量研究。刘振生等采用直接观察法对该区域岩羊冬春季的取食生境选择进行了研究[12]。刘振生等对贺兰山地区岩羊的种群数量及分布做了调查和报道,2003年岩羊种群数量约为10 000只,东坡是岩羊的主要分布区[13]。刘振生等利用距离取样法对贺兰山地区不同季节岩羊的种群密度进行了估测[14]。在岩羊的生境评价方面,尚缺乏相关的报道。
近年来,随着3S(GIS,RS,GPS)技术的发展,从大尺度上对野生动物进行生境研究成为热点[15-16]。国内外的众多学者利用机理模型、回归模型和生态位模型结合3S技术来对目标物种进行生境适宜性评价[17-19]。最大熵模型是生态位模型的一种,近年来已广泛应用于保护动物的生境分布预测。李明阳等基于WEB数据库和MAXENT模型,对白头叶猴的潜在生境进行评价研究[20]。齐增湘等利用MAXENT模型,进行秦岭山系黑熊潜在生境预测[21]。徐卫华等采用MAXENT模型,对秦岭地区的川金丝猴的生境进行评价[22]。本文应用GIS技术和MAXENT模型对贺兰山岩羊生境适宜性进行评价,分析影响岩羊生存的主要生境因子并模拟其空间分布区域,以期为贺兰山国家级自然保护区的合理规划及岩羊的保护提供科学依据。
1 材料与方法
1.1 研究地区概况
贺兰山位于银川平原与阿拉善高原之间,地理坐标为 N38°21'—39°22',E105°49'—106°42',海拔一般是2 000—3 000 m,由内蒙古贺兰山国家级自然保护区和宁夏贺兰山国家级自然保护区两部分组成,是岩羊的一个重要的集中分布区。贺兰山具有典型的大陆性气候特征,地处荒漠与半荒漠地带,主要特点是干旱、高寒。年平均气温为-0.9℃,平均年降水量在200—400 mm之间[23]。冬季长达5个月,降雪较少。贺兰山是典型的温带山地森林系统,其植被垂直分布明显:山地草原带(1400—1600 m),山地疏林草原带(1600—2000 m),山地针叶林带(1900—3000 m),亚高山灌丛和草甸带(3000—3556 m)[24]。
1.2 模型介绍
最大熵原理(MAXENT)起源于信息科学,最早由Jaynes提出,在许多学科中有广泛应用,近年来引起了生态学家们的关注[25]。MAXENT是一种基于生态位原理的模型,根据物种“出现点”的环境变量特征得出约束条件,探寻此约束条件下最大熵的可能分布,以此来预测目标物种在研究地区的生境分布[26]。2004年,Phillips等人开发了MAXENT软件,近来广泛用于物种生境适宜区的预测和评价,表现出了良好的预测能力[27]。该模型提供了自检验功能,可以自动生成ROC曲线进行模型的模拟预测自检,且在对动物生境进行评价与预测时,只需动物“出现点”的数据,并且具有较高的精度[28-30]。在岩羊“未出现点”的数据难以准确采集的实际情况下,MAXENT模型相比其他模型具有更大的优势。
1.3 数据来源及预处理
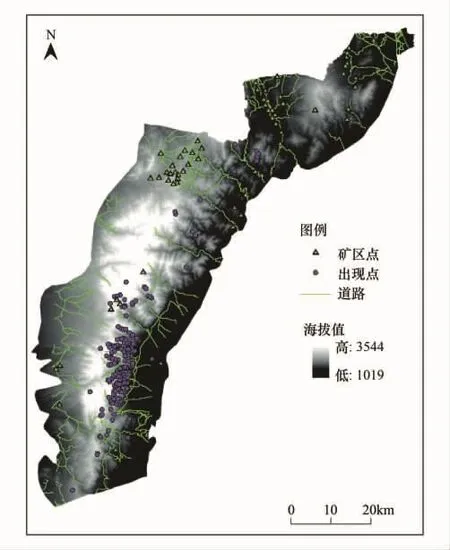
图1 贺兰山概况及样点分布图Fig.1 The overview of Helan Mountain and distribution of sample points
应用MAXENT模型对岩羊进行生境适宜性评价需要两方面数据:一是岩羊的现实地理分布点数据,二是贺兰山地区的环境变量数据。其中岩羊分布点数据来源于野外调查以及贺兰山保护区各管理站的野外监测,自2010年至2011年,根据贺兰山各个沟段的实际情况,分别在宁夏贺兰山国家级自然保护区的马莲口、苏峪口、大水沟、红果子4个管理站和内蒙古贺兰山国家级自然保护区的腰坝、哈拉乌、水磨沟、古拉本4个管理站的25条沟系,按春、夏、秋、冬4个季节以步行的方法对岩羊进行调查。这些沟系分布于不同的海拔高度,覆盖了贺兰山岩羊栖息的全部生境类型,保证了分布点数据的代表性和准确性。用GPS对所观察到的岩羊实体或岩羊利用痕迹的位置进行定点记录,在贺兰山与岩羊同域分布的有蹄类还有马麝(Moschus chrysogaster)和马鹿(Cervus elaphus),可根据体形大小和形态特征对它们进行区分,共得到了317个岩羊出现的GPS坐标点。环境变量数据包括:①地形因子数据,包括海拔、坡度和坡向,由中国科学院科学数据库30 m分辨率的DEM(数字高程图)计算提取得到;②植被因子数据,利用ERDAS 9.1遥感影像处理软件,采用监督分类方法对2007年贺兰山的TM影像进行分类处理,将其分为针叶林、阔叶林、灌丛、草甸、草地、荒漠和水体等7种类型;③其他数据,包括护林点、矿区、道路及水源因子,从贺兰山1∶50000的矢量化地图中提取获得,利用ARCGIS的空间分析求算到各因子的距离栅格图层。
以ARCGIS为平台,去除边界之外的GPS坐标点,将所有环境变量的图层统一边界,坐标系统统一为WGS-1984-UTM-Zone-48N,栅格大小统一为30×30 m,并转化成MAXENT软件所要求的ASCII格式的文件。

图2 贺兰山岩羊生境评价结果的ROC曲线验证Fig.2 ROC curve verification of blue sheep in the Helan Mountains,China
1.4 模拟方法
将岩羊现实分布点的数据和环境变量数据导入MAXENT 3.3中,随机选取75%的岩羊分布点用于建立模型,其余25%的岩羊分布点用于模型验证,选择利用Jackknife来检测变量的重要性,并对各生境因子进行敏感性分析,其它参数均为模型的默认值,结果以Logistic格式输出。模型的预测结果利用受试者工作特征曲线下的面积值进行检验,其评价标准为:曲线下的面积的值在 0.5—0.6,不及格;0.6—0.7,较差;0.7—0.8,一般;0.8—0.9,良好;0.9—1.0,优秀[31]。将模型输出结果放入 ArcGIS 中进行进一步分析,按照专家经验法[32],对模型预测的贺兰山岩羊分布图进行重新分类。
2 结果与分析
2.1 MAXENT预测结果检测
ROC曲线评价结果为:训练集与验证集的AUC值分别为0.915和0.901,表明MAXENT模型的预测结果达到优秀水平(图1)。
2.2 岩羊分布与环境因子的关系
Jackknife检验结果表明:坡度、距矿区距离、海拔、距道路距离、距水源距离是影响岩羊生境分布的主要环境因子。环境变量对MAXENT模型的贡献率说明:距矿区距离(35.5%)、坡度(27.8%)、海拔(12.6%)、距道路距离(10.2%)和距水源距离(7.3%)5个环境变量的累积贡献率达86%,对岩羊的生境选择具有重要影响,矿区和道路是对岩羊影响最大的2个人为干扰因子;针叶林、灌木、水体和荒漠等因子的贡献率为零,对岩羊的生境选择没有影响;其余因子的影响率较小,贡献值由大到小排序依次为坡向、护林点、草地、草甸和阔叶林,坡向对岩羊的生境选择具有一定影响,岩羊的主要食物为草本科植物和灰榆,灰榆主要分布于阔叶林中,因此草地、草甸和阔叶林对岩羊的选择具有影响。各生境因子的敏感性分析表明,随着坡度、距矿区的距离、距道路的距离的增大,生境适宜度逐渐增加,表明岩羊偏爱崎岖陡峭地带,躲避存在人为干扰的区域,在道路分析中,距其约100 m的范围内干扰值最大,超过100 m后,道路对岩羊的干扰逐渐减少;随着距水源距离的增加,生境适宜度逐渐减小;适宜度在1500—2300 m之间随着海拔增加而增大,超过2 300 m之后随着海拔的增加而逐渐减小。
2.3 贺兰山岩羊生境适宜性分布
利用MAXENT模型对贺兰山区域岩羊的生境适宜性进行评价,将模型输出的ASCII文件导入到ArcGIS 10中,转换成浮点型栅格数据,按照专家经验法将生境适宜图重新分为3个适宜等级:0.00—0.12为不适宜等级,0.12—0.40为较适宜等级,0.40—1.00为最适宜等级,最终得到贺兰山地区岩羊的生境适宜性分布图(图2)。
从图中可以看出,贺兰山岩羊的适宜生境主要分布于贺兰山东坡(宁夏贺兰山国家级自然保护区)西南部及西坡(内蒙古贺兰山国家级自然保护区)的山地疏林草原带。其中最适宜生境的面积为262.3 km2;较适宜生境面积为744.6 km2,总的适宜性生境面积占研究区域面积的28%;不适宜生境面积为2649.1 km2。
3 讨论
生境评价模型目前可以分为3种类型:机理模型、回归模型和生态位模型。机理模型并不需要物种分布点数据,而是根据生境因子对物种分布的影响建立相应的评价准则,进而模拟得到物种的适宜性生境,但因其没有考虑生境的可达性,且在对因子划分等级及确定权重上存在主观性,因此具有一定局限性。相比回归模型,生态位模型只需动物“出现点”的数据而不需要“未出现点”的数据,根据在贺兰山野外调查的实际经验,发现在某个地点未发现岩羊的踪迹并不代表岩羊未曾在此出现过,因此所记录的“未出现点”的数据并非十分准确,相比较而言,利用生态位模型可信度更高。本次研究利用MAXENT模型对贺兰山岩羊生境适宜性进行评价,所取得的结果较为理想。
岩羊“出现点”数据的收集方法是对发现岩羊实体或者岩羊利用痕迹的位置用GPS进行定点记录,但是由于贺兰山山势起伏较大,多陡坡和断崖等地形,影响了一些出现点的可达性,部分出现点数据不能精确记录,如岩羊行走的险峻山崖,无法进行准确定点。实际的野外调查中,记录者在能够到达的最接近目标地位置进行定点记录,以对评估结果造成的影响降到最小。此外,道路数据是从贺兰山1∶50000的矢量化地图中提取获得,道路对岩羊影响力的大小与其车流量和宽度有关,而每条道路的建路时间以及车流量等具体信息并不明确,在数据提取过程中,忽略了山间小路这些干扰性小的数据信息,以减少误差。对于采集数据点达到精确以及提高道路信息准确度的方法仍需进一步的研究改进,以期取得更准确的评价结果。
已有研究表明岩羊偏爱靠近裸岩及悬崖的生境[33],选择山势陡峭的山地疏林草原带,避免选择山地针叶林带[34]。模型分析结果表明,自然生境因子中贡献率最大的为坡度和海拔。随着坡度的增大,地势陡峭度增加,生境适宜性逐步提高。海拔高度决定了贺兰山植被的分布,在1 500—2 300 m海拔内,主要为山地疏林草原带,生境适宜性逐步增加,其后随着海拔增高,进入山地针叶林带,生境适宜性逐渐降低,这些分析结果与之前的研究相符。模型预测结果显示贺兰山东坡(宁夏贺兰山国家级自然保护区)的岩羊适宜分布区的面积要远大于西坡(内蒙古贺兰山国家级保护区),这主要是因为东坡山势陡峻狭短,沟谷深切,地面较破碎,崎岖度较大,且有大面积的山地疏林草原带,而西坡的坡面缓长,山地针叶林带较多[13],因此相比较西坡,东坡具有更广阔的岩羊适宜生境。
贺兰山是岩羊的一个重要集中分布区域,也是岩羊分布的最北缘[35]。自20世纪80年代保护区建立以来,保护和管理力度逐年加强,采取了禁止和限制家畜放牧等一系列保护措施[36],使贺兰山植被得以恢复,环境承载力得到提高。从本研究结果来看,目前最重要的2个人为干扰因素是矿区和道路。针对矿业开采对岩羊造成的影响,保护区应采取相应措施,严格控制岩羊生境区内矿区的范围和数量,使其对岩羊的干扰降到最低。同时应该加强贺兰山地区道路的规划和管理,新建道路的选线应尽量避开岩羊频繁出没的地段。此外,在对贺兰山开发建设的同时,应加强对周围居民的宣传教育,使人们充分认识到岩羊作为贺兰山的优势种具有重要的意义和价值,提高居民的野生动物保护意识,以期对岩羊种群的稳步发展有一定的积极作用。

图3 贺兰山岩羊生境适宜性分布Fig.3 The distribution of habitat suitability of blue sheep in Helan Mountains,China
[1]Wang J L,Chen Y.Applications of 3Stechnology in wildlife habitat researches.Geography and Geo-Information Science,2004,20(6):44-47.
[2]Wang X L.Landscape Dynamics and Suitability Assessment of Przewalski's Gazelle Habitat[D].Beijing:Chinese Academy of Forestry,2004.
[3]Baillie J,Groombridge B.IUCN red list of threatened animals.Gland,Switzerland:World Conservation Union,1996.
[4]Liu Z S.Ecology of Blue Sheep(Pseudois nayaur)in Helan Mountains,China[D].Shanghai:East China Normal University,2006.
[5]Sheng H L.Handbook of Fur-bearing Animal.Shanghai:Shanghai Lexicographical Publishing House,1994:218-219.
[6]Oli M K,Rogers M E.Seasonal Patten in group size and population composition of blue sheep in Manang,Nepal.Journal of Wildlife Management,1996,60(4):797-801.
[7]Liu G K,Zhou C Q,Yang Z S,Long S,Pan L,Wang W K,You Y Q,Zeng G W,Geng SS.Diurnal activity rhythm and time budgets of the dwarf blue sheep(Pseudois schaeferi)in Zhubalong Nature Reserve.Acta Ecologica Sinica,2011,31(4):972-981.
[8]Chang Y,Liu Z S,Zhang M M,Li Z G,Hu T H.Summer diets of sympatric blue sheep(Pseudois nayaur)and red deer(Cervus elaphus alxaicus)in the Helan Mountains,China.Acta Ecologica Sinica,2010,30(6):1486-1493.
[9]Wilson P.Ecology and habitat utilization of blue sheep Pseudois nayaur in Nepal.Biological Conservation,1981,21(1):55-74.
[10]Wang X M,Liu Z X,Xu H F,Li M,Li Y G.The blue sheep population ecology and its conservation in Helan Mountain,China.Biodiversity Science,1998,6(1):1-5.
[11]Harris R B,Miller D J.Overlap in summer habitats and diets of Tibetan Plateau ungulates.Mammalia,1995,59(2):197-212.
[12]Liu Z S,Wang X M,Li ZG,Cui D Y,Li X Q.Comparison of seasonal feeding habitats by blue sheep(Pseudois nayaur)during winter and spring in Helan Mountain,China.Zoological Research,2005,26(6):580-589.
[13]Liu Z S,Wang X M,Li Z G,Zhai H,Hu T H.Distribution and abundance of blue sheep in Helan Mountains,China.Chinese Journal of Zoology,2007,42(3):1-8.
[14]Liu Z S,Wang X M,Teng L W,Cui D Y,Li X Q.Estimating seasonal density of blue sheep(Pseudois nayaur)in the Helan Mountain region using distance sampling methods,Ecological Research,2008,23:393-400.
[15]Xu W H,Ouyang Z Y,Li Y,Liu J G.Habitat assessment on giant panda's in Qinling Mountain Range based on RS and GIS.Remote Sensing Technology and Application,2006,21(3):239-242.
[16]Ward D F.Modelling the potential geographic distribution of invasive ant species in New Zealand.Biological Invasions,2007,9(6):723-735.
[17]Steven JP,Miroslav D,Robert E S.A Maximum Entropy Approach to Species Distribution Modelling.Banff,Alberta:Proceedings of the Twenty-First International Conference on Machine Learning,2004:655-662.
[18]Li H M,Han H X,Xue D Y.Prediction of potential geographic distribution areas for the pine bark scale,Matsucoccus matsumurae(Kuwana)(Homoptera:Margarodidae)in China using GARP modelling system.Acta Entomologica Sinica.2005,48(1):95-100.
[19]Brotons L,Thuiller W,Miguel B A,Alexandre H H.Presence-absence versus presence-only modelling methods for predicting bird habitat suitability.Ecography,2004,27(4):437-448.
[20]Li M Y,Xi Q,Xu H G.Evaluation of potential habitat for white-headed langur based on WEB dataset and Maxent model.Forest Resources Management,2010,(4):31-36.
[21]Qi Z X,Xu W H,Xing X Y,Ouyang Z Y,Zheng H,Gan D X.Assessment of potential habitat for Ursusthibetanus in the Qingling Mountains,Biodiversity Science,2011,19(3):343-352.
[22]Xu WH,Luo C.Application of MAXENT model in Rhinopithecusroxllanae habitat assessment in Qinling Mountain.Forest Engineering,2010,26(2):1-4.
[23]Liu Z S,Li X Q,Wang X M,Li Z G,Hu T H,Zhai H.Seasonal variations in group types and sizes of blue sheep(Pseudoisnayaur)in the Helan M ountains,China.Acta Ecologica Sinica,2009,29(6):2782-2788.
[24]Cui D Y,Liu Z S,Wang X M,Zhai H,Hu T H,Li Z G.Winter food-habits of red deer(Cervus elaphus alxaicus)in Helan Mountains,China.Zoological Research,2007,28(4):383-388.
[25]Xing D L,Hao Z Q.The principle of maximum entropy and its applications in ecology.Biodiversity Science,2011,19(3):295-302.
[26]Phillips SJ,Anderson R P,Schapire R E.Maximum entropy modelling of species geographic distributions.Ecological Modelling,2006,190(3/4):231-259.
[27]Luo C,Xu W H,Zhou ZX,Ouyang ZY,Zhang L.Habitat prediction for forest musk deer(Moschusberezovskii)in Qinling mountain range based on niche model.Acta Ecologica Sinica,2011,31(5):1211-1229.
[28]Stachura-Skierczynska K,Tumiel T,Skierczyński M.Habitat prediction model for three-toed woodpecker and its implication for the conservation of biologically valuable forests.Forest Ecology and Management,2009,258(5):697-703.
[29]Haegeman B,Etienne R S.Entropy maximization and the spatial distribution of species.The American Naturalist,2010,175(4):74-90.
[30]Li M Y,Ju Y W,Kumar S,Stohlgren T J.Modelling potential habitat for alien species of Dreissena polymorpha in the continental USA.Acta Ecologica Sinica,2008,28(9):4253-4258.
[31]Swets J A.Measuring the accuracy of diagnostic systems.Science,1988,240:85-93.
[32]Xi Q.Research on Potential Distribution of the White-headed Langur Based on GISTechnology[D].NanJing:Nanjing Forestry University,2010.
[33]Oli M K.Seasonal patterns in habitat use of blue sheep Pseudoisnayaur(Artiodactyla,Bovidae)in Nepal.Mammalia,1996,60(2):187-193.
[34]Liu Z S,Cao L R,Wang X M,Li T,Li Z G.Winter bed-site selection by blue sheep(Pseudois nayaur)in Helan Mountains,Ningxia,China.Acta Theriologica Sinica,2005,25(1):1-8.
[35]Wang X M,Schaller GB.Status of large mammals in Inner Mongolia,China.Journal of East China Normal University:Special Issue of Mammals.1996,6(1):94-104.
[36]Wang X M,Liu ZS,Li X Q,Li Z G.Comparison of age structures of male blue sheep(Pseudois nayaur)on Helan Mountain in two periods,1995 and 2004.Zoological Research,2005,26(5):467-472.
参考文献:
[1]王金亮,陈姚.3S技术在野生动物生境研究中的应用.地理与地理信息科学,2004,20(6):44-47.
[2]王秀磊.普氏原羚生境的景观动态与适宜性评价研究[D].北京:中国林业科学研究院,2004.
[4]刘振生.岩羊 (Pseudois nayaur)生态学研究[D].上海:华东师范大学,2006.
[5]盛和林.毛皮动物手册.上海:上海辞书出版社,1994:218-219.
[7]刘国库,周材权,杨志松,龙帅,潘立,王维奎,游于群,曾国伟,耿山山.竹巴笼矮岩羊昼间行为节律和时间分配.生态学报,2011,31(4):972-981.
[8]昶野,刘振生,张明明,李志刚,胡天华.贺兰山同域分布岩羊和马鹿的夏季食性.生态学报,2010,30(6):1486-1493.
[10]王小明,刘志霄,徐宏发,李明,李元广.贺兰山岩羊种群生态及保护.生物多样性,1998,6(1):1-5.
[12]刘振生,王小明,李志刚,崔多英,李新庆.贺兰山岩羊冬春季取食生境的比较.动物学研究,2005,26(6):580-589.
[13]刘振生,王小明,李志刚,翟昊,胡天华.贺兰山岩羊的数量与分布.动物学杂志,2007,42(3):1-8.
[15]徐卫华,欧阳志云,李宇,刘建国.基于遥感和GIS的秦岭山系大熊猫生境评价.遥感技术与应用,2006,21(3):239-242.
[18]李红梅,韩红香,薛大勇.利用GARP生态位模型预测日本松干蚧在中国的地理分布.昆虫学报,2005,48(1):95-100.
[20]李明阳,席庆,徐海根.基于WEB数据库和Maxent模型的白头叶猴潜在生境评价研究.林业资源管理,2010,(4):31-36.
[21]齐增湘,徐卫华,熊兴耀,欧阳志云,郑华,甘德欣.基于MAXENT模型的秦岭山系黑熊潜在生境评价.生物多样性,2011,19(3):343-352.
[22]徐卫华,罗翀.MAXENT模型在秦岭川金丝猴生境评价中的应用.森林工程,2010,26(2):1-4.
[23]刘振生,李新庆,王小明,李志刚,胡天华,翟昊.贺兰山岩羊 (Pseudois nayaur)集群特征的季节变化.生态学报,2009,29(6):2782-2788.
[24]崔多英,刘振生,王小明,翟昊,胡天华,李志刚.贺兰山马鹿冬季食性分析.动物学研究,2007,28(4):383-388.
[25]刑丁亮,郝占庆.最大熵原理及其在生态学研究中的应用.生物多样性,2011,19(3):295-302.
[27]罗翀,徐卫华,周志翔,欧阳志云,张路.基于生态位模型的秦岭山系林麝生境预测.生态学报,2011,31(5):1211-1229.
[30]李明阳,居云为,Kumar S,Stohlgren T J.美国大陆外来入侵物种斑马纹贻贝 (Dreissena polymorp ha)潜在生境预测模型.生态学报,2008,28(9):4253-4258.
[32]席庆.基于GIS技术的白头叶猴潜在分布研究[D].南京:南京林业大学,2010.
[34]刘振生,曹丽荣,王小明,李涛,李志刚.贺兰山岩羊冬季对卧息地的选择.兽类学报,2005,25(1):1-8.
[36]王小明,刘振生,李新庆,李志刚.贺兰山雄性岩羊两个时期生命表的比较.动物学研究,2005,26(5):467-472.
