饲粮n-6/n-3多不饱和脂肪酸比例对畜禽健康和产品品质的影响
2013-12-20高巧仙宋代军
高巧仙 宋代军 靳 露
(西南大学动物科技学院,重庆 400716)
人类膳食中n-6与n-3多不饱和脂肪酸(polyunsaturated fatty acids,PUFA)摄取比例失衡会导致心脑血管疾病、癌症、炎症以及自身免疫性疾病的发生[1]。目前人类膳食中 n-6/n-3 PUFA比例极不平衡,n-6/n-3 PUFA比例已由过去的(1~2)∶1提高至现在的(20 ~30)∶1,2006 年澳大利亚健康与医疗委员会推荐膳食中n-6/n-3 PUFA的最佳比例为(2 ~4)∶1[2]。由于 n-6 与 n-3 PUFA 之间在生物体内不能相互转化,代谢过程相互竞争,因此只有在二者的数量及比例合理的前提下,才能发挥更好的生理作用。故本文主要根据当前的研究结果,综述饲粮中n-6/n-3 PUFA比例平衡的营养生理作用以及对畜禽产品品质的一些影响及其调控方法。
1 PUFA的种类和来源
人类和其他哺乳动物因自身不能合成PUFA而必须从食物中获得。PUFA是指含有2个及2个以上双键的脂肪酸,主要包括 n-9、n-6和 n-3 PUFA,其中具有特殊生理功能的是 n-6和 n-3 PUFA。
1.1 n-3 PUFA
n-3 PUFA包括 α-亚麻酸(α-linolenic acid,ALA)、二 十 碳 五 烯 酸 (eicosapentaenoic acid,EPA)、二十二碳六烯酸(docos ahexaenoic,DHA)、二十二碳五烯酸(docosapentenoic acid,DPA)等。其中,ALA主要来源于植物油,如亚麻籽油;EPA和DHA主要来源于鱼类等海洋生物[3]。
1.2 n-6 PUFA
n-6 PUFA 包括亚油酸(linoleic acid,LA)、γ -亚麻酸(γ-linolenic acid,GLA)和花生四烯酸(arachidonic acid,AA)等。LA与 GLA存在于同一植物油中,如葵花籽油、玉米油、大豆油等。
2 PUFA的营养代谢
PUFA可转化为高不饱和脂肪酸或链更长的脂肪酸,其生物代谢途径如图1所示。LA、ALA分别是n-6、n-3 PUFA的前体,n-3和n-6 PUFA在动物体内的代谢至少在以下3种酶中存在竞争:脂肪酸转化过程中的去饱和酶Δ-6和Δ-5,碳链的增长酶(延长酶),且n-3和n-6 PUFA在体内不能相互转化。
3 n-6/n-3 PUFA比例平衡的营养生理作用
3.1 n-6/n-3 PUFA比例平衡可防治心血管疾病
n-6 PUFA会引发血小板凝集和血栓的形成,而n-3 PUFA则有相反作用。血小板膜磷脂释放的PUFA经环氧合酶作用可合成血栓素-凝血恶烷A(thromboxane A,TXA),具有促进血小板凝集和收缩血管的作用;血管壁膜磷脂释放的PUFA经酶促作用合成的前列环素Ⅰ(prostacyclinⅠ,PGⅠ)具有抑制血小板凝集和舒张血管的作用,二者共同调节血小板和血管功能及血栓的形成[1]。细胞膜磷脂中释放出AA合成TXA2和PGI2,EPA生成TXA3和PGI3。EPA与AA结构相似,与AA在同一水平上竞争环氧合酶,从而竞争性抑制AA向TXA2和PGI2转化。TXA2的促血小板聚集作用强于TXA3,而PGI2与PGI3抑制血小板聚集的作用相仿,结果就降低了血小板聚集性并增加了血管舒张作用,使血栓的形成减少。因此,n-6 PUFA过量会致使血液黏度、血管痉挛度和血管收缩度升高,而增加n-3 PUFA则可缓解这些症状。但过量的 n-3 PUFA会加重出血的倾向,EPA、DHA摄取过剩则会过度抑制血栓素TXA2的形成,造成出血时间延长,使出血现象加重[1]。
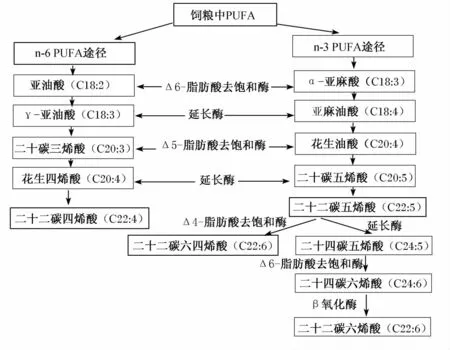
图1 PUFA的生物代谢途径Fig.1 Biological metabolic pathways of PUFA[4-5]
3.2 n-6/n-3 PUFA比例平衡影响基因表达的调控
PUFA是重要的细胞信号分子,经细胞磷脂酶作用下从膜磷脂中释放出来。PUFA不仅能够作为第二信使或替代传统的第二信使肌醇磷脂和环腺苷酸信号转导通路途径,还可以改变特定基因的转录[6]。EPA、DHA能抑制外周巨噬细胞释放的白细胞介素-1(IL-1)mRNA的转录,而AA能适度抑制血管细胞黏附分子的表达[7]。
3.3 n-6/n-3 PUFA比例平衡对脂类代谢影响
PUFA可抑制脂肪合成酶和糖酵解酶基因表达,促进脂肪活化基因的表达,从而抑制脂类合成,加速脂类分解[8]。n-3 PUFA对脂肪酸和脂肪酸合成酶的抑制作用强于n-6 PUFA。Duplus等[9]报道,AA 及其代谢物前列腺素 E(PGE),特别是前列腺素E2(PGE2)可与脂肪细胞转录因子迅速反应并激活它,活化脂肪细胞,促进其代谢,从而使机体脂肪水平下降。同时,n-3 PUFA具有降低低密度胆固醇和极低密度胆固醇,增加高密度脂蛋白的作用,由于高密度脂蛋白是一种有利于健康的胆固醇形式,它可以使胆固醇从细胞内转移到肝脏中,从而使胆固醇被肝脏代谢。
3.4 n-6/n-3 PUFA比例平衡对免疫性能的影响
PUFA组成和含量的多少会影响到免疫细胞膜表面的抗原、抗体数量、分布以及淋巴因子、抗体的分泌。在机体受到外界抗原刺激时,淋巴因子和抗体的分泌及新免疫细胞的产生等都依赖脂肪的参与,因而细胞膜磷脂中脂肪酸组成的变化将对免疫功能产生影响。饲粮n-6/n-3 PUFA比例平衡可能通过对免疫细胞的调节影响机体免疫反应。不同的n-6/n-3 PUFA比例会对免疫功能产生不同的影响,因此确定合适的n-6/n-3 PUFA比例是研究动物体免疫功能的关键。因为二者在体内由同一种酶催化代谢,在影响机体免疫机制方面发生竞争效应。Volgarev等[10]研究用体重(135.0±3.2)g的雄性鼠进行饲养试验以研究不同比例的n-6和n-3 PUFA对免疫功能和非特异性抗性的影响。n-6/n-3 PUFA的比例分别为1.1∶1.0、2.1∶1.0、6.1∶1.0 和 49.0∶1.0,结果表明与比例为49.0∶1.0的试验组相比,n-6/n-3 PUFA比例低的能增强免疫反应和非特异性抗性。对外周淋巴组织器官的分析结果表明,PUFA可以促进外周淋巴细胞的增殖,且组织中PUFA的含量随着饲粮中PUFA含量的增加而增加。Pilevar等[11]研究饲粮中不同的n-6/n-3 PUFA比例对鸡的免疫和生殖系统的影响,n-6/n-3 PUFA比例是2∶1、6∶1、10∶1,结果表明饲粮中 n-6/n-3 PUFA 比例越低,对鸡新城疫、传染性支气管炎、传染性法氏囊病的抗体量产生的越多。
4 饲粮中n-6/n-3 PUFA比例对畜禽产品脂肪酸成分的影响
动物组织脂肪酸有2种来源,即内源(动物体内生物合成)和外源(饲粮提供)。PUFA沉积几乎全部由饲粮提供。饲粮中脂肪酸种类和含量直接影响畜禽产品的脂肪酸种类和含量。
4.1 反刍动物
富含n-3、n-6 PUFA饲粮的来源和处理方法影响畜禽产品中的n-6/n-3 PUFA比例。研究表明,饲粮中添加未经处理的亚麻籽或亚麻油可使牛奶中LA含量在一定范围内升高;而添加挤压亚麻油还可增加牛奶ALA的含量,降低n-6/n-3 PUFA比例,从而提高牛奶和奶酪的乳脂品质[12-13]。饲粮中添加挤压或压碎的亚麻籽具有同等效果,即提高牛肌肉中ALA的含量[14]。有学者研究比较饲粮中添加不同来源植物油对牛肉不同部位脂肪酸成分的影响,结果表明,添加亚麻籽组牛肌肉中LA含量显著高于其他各组,且n-6/n-3 PUFA比例是最低[15]。Wachira等[16]比较饲粮中添加未经处理的亚麻籽、鱼油、未经处理的亚麻籽和鱼油的混合物及对照组对羊肉n-3 PUFA含量影响,结果表明,添加亚麻籽组的羊肉ALA含量高于其他3组,未经处理的亚麻籽组、鱼油组、未经处理的亚麻籽和鱼油的混合物组羊肉n-3 PUFA含量高于对照组;鱼油组羊肉DHA含量稍高于对照组,结合先前的试验说明了反刍动物ALA转化为长链n-3 PUFA的限制性。
4.2 单胃动物
猪肉和鸡肉脂肪中的脂肪酸组成在一定程度上反映了其所摄入脂肪的量。富含n-3 PUFA饲粮的脂肪酸含量不仅会直接影响猪器官和组织中脂肪酸含量,还可以间接通过脂肪生成酶的表达影响肉中脂肪酸含量[17]。猪、鸡饲粮中常添加亚麻籽油而不是亚麻籽种子,Mourot等[24]报道了3种不同饲粮(分别添加牛油、油菜籽油、亚麻籽油)对猪肉和鸡肉脂肪酸含量的影响。这3种饲粮中,添加亚麻籽油的饲粮提高了猪、鸡肉中n-3 PUFA含量,降低了n-6/n-3 PUFA比例。由于亚麻籽油富含ALA而不是其他的长链n-3 PUFA,且鸡肝脏中ALA转化为长链n-3 PUFA的效率低,因此与鱼油和富含n-3 PUFA油相比,饲粮中亚麻籽油会减少鸡肉中EPA和DHA的沉积[18]。
鸡蛋和兔肉中的脂肪酸含量也可通过饲粮来改善。Shapira等[19]报道,蛋鸡饲喂添加挤压亚麻籽的饲粮可提高蛋黄中n-3 PUFA含量,与对照组相比,强化蛋中总n-3 PUFA、ALA和DHA的含量分别增加了 3.8、6.4、2.4 倍;n-6/n-3 PUFA 比例降低了3.6倍。Kouba等[20]研究表明,饲喂亚麻籽的兔肉中n-3 PUFA含量(451 mg/g)高于对照组(153 mg/g)。
4.3 熟肉及肉制品
烹饪能影响肉脂肪酸含量和种类,但不影响脂肪酸的分布,且与饲粮无关。Guillevic等[21]研究表明,烹饪后的熟肉中n-3 PUFA含量显著提高;添加4%挤压的亚麻籽组经烹饪后n-6/n-3 PUFA比例由2.66升高至2.82,而对照组的n-6/n-3 PUFA 比例由14.31降至12.51;另外,不论制成什么产品,只要饲粮中添加亚麻籽都会提高产品中n-3 PUFA含量,极显著降低n-6/n-3 PUFA比例。Kouba等[20]研究表明,兔子熟肉中的脂肪酸含量高于生肉,而n-6 PUFA含量增加却不如n-3 PUFA含量,这可能是因为烹饪损失及n-3 PUFA是结构性的脂质,其结构在烹饪过程中不易改变;n-6/n-3 PUFA比例由生肉的2.42极显著降低至熟肉的 2.35。
4.4 富含n-6、n-3 PUFA畜禽产品易氧化
长链的PUFA分子内含有很多双键,易被氧化成饱和脂肪酸。富含PUFA的肉易氧化,肉保存、煮熟的时间和温度不同会强化氧化。脂肪酸氧化导致肉的营养价值损失、感官特性降低并形成潜在的有毒物质,进而影响肉品质和降低货架期[22]。饲粮中添加α-生育酚(维生素E)可提高鸡肉的抗氧化性,α-生育酚抗氧化性伴随肉的感官特性受饲粮中其添加量、饲粮中油脂的不饱和性和氧化性、动物生理状态及肉中净氧化平衡的影响[23]。维生素E沉积受饲粮中PUFA含量多少的影响,饲粮中PUFA含量越高,鸡肉中维生素E沉积得越少。因此,饲粮中添加维生素E应与饲粮中PUFA水平相适应。低PUFA含量(15 g/kg)的饲粮中添加60 mg/kg维生素E能防止肉脂质的氧化性,而高PUFA含量(30 g/kg)的饲粮需添加 200 mg/kg 的维生素 E[24]。Gobert等[25]研究表明,产奶中期的奶牛饲喂富含n-3 PUFA饲粮,添加抗氧化剂维生素E与植物多酚混合物可提高血浆脂质的抗氧化性,从而降低富含n-3 PUFA乳制品脂质的氧化。
5 改善畜禽产品n-6/n-3 PUFA比例的方法
5.1 通过基因表达改善畜禽产品n-6/n-3 PUFA比例
哺乳动物体内缺乏n-3脂肪酸脱氢酶(去饱和酶),不能使n-6 PUFA转化为n-3 PUFA,而转基因哺乳动物体外细胞和体内组织能够有效表达低等生物的去饱和酶基因,将过多的n-6 PUFA或者饱和脂肪酸催化为相应的n-3 PUFA,从而使n-6/n-3 PUFA比例平衡,改变哺乳动物本身无法自主合成的必需脂肪酸。研究发现将线虫(C.elegans)n-3 PUFA脱氢酶序列载入到转基因小鼠体内,使得小鼠奶脂质中 n-3 PUFA含量显著增加[26]。Kao等[27]在山羊哺乳期诱导并分离出乳腺β-酪蛋白启动子,通过原核显微镜注射到含有线虫n-3 PUFA的脱氢酶(去饱和酶)转基因老鼠体内,发现可使该小鼠产生的奶中n-3 PUFA增加,n-6 PUFA降低,n-6/n-3 PUFA比例降低。且新生小鼠食用该转基因雌鼠的奶后,脑中DHA水平增加,这一转基因模型证明了可从内源性途径提高奶及奶制品中的n-3 PUFA含量。虽然研究者通过转基因工程和PUFA代谢途径相结合研究取得了进展,但脂肪酸去饱和酶在转基因过程中是否会突变,是否对转基因老鼠健康有危害,如 n-6/n-3 PUFA比例影响类固醇激素、十二烷类如前列腺素的合成,前列腺素作为重要的细胞信号分子是否与繁殖性能有关,这些问题及机制都有待进一步研究。
5.2 通过饲粮改善畜禽产品n-6/n-3 PUFA比例
饲粮n-3 PUFA的来源(鱼油、植物油、种子)对不饱和脂肪酸/饱和脂肪酸(P/S)的比值无影响。P/S主要受遗传的影响[28]。由于反刍动物瘤胃微生物的氢化作用,可使不饱和脂肪酸氢化成饱和脂肪酸,进而影响反刍动物肉制品的P/S,但单胃动物无此功能。饲粮因素主要影响畜产品中的n-6/n-3 PUFA比例。富含n-3 PUFA的饲粮在增加肉中n-3 PUFA总含量的同时降低了n-6 PUFA的沉积,且n-6/n-3 PUFA比例也随之降低。
5.2.1 添加鱼油、植物油降低畜禽产品n-6/n-3 PUFA比例
鱼油、植物油如亚麻籽油等富含n-3 PUFA,在饲粮中添加富含n-3 PUFA饲料可提高畜禽产品的 n-3 PUFA含量,降低 n-6/n-3 PUFA比例。Kouba等[29]研究发现,在猪饲粮中添加6%的亚麻籽,并在第20、60和100天对猪肉中的脂肪酸进行分析,n-6/n-3 PUFA 比例从 7.64、7.34 和8.71分 别 下 降 到 3.90、3.00 和 3.11。Collins等[30]报道,在饲粮中添加了5%亚麻油的羊肉中,n-6/n-3 PUFA 比例从 4.61下降为 2.44。Betti等[31]研究表明,鸡饲粮中添加17%、10%的亚麻籽分别在11.3、26.2 d使得每100 g鸡胸肉中含有300 mg n-3 PUFA;由于饲粮中添加亚麻籽,肉中n-3 PUFA含量显著增加,胸肌肉和腿肌肉的n-6/n-3 PUFA比例显著降低。Oliveira等[32]在产蛋鸡饲粮中添加3.4%的亚麻油,与对照组相比,产蛋鸡初期产蛋的蛋黄中n-6/n-3 PUFA比例由12.99降为2.01,产蛋鸡后期产蛋的蛋黄中 n-6/n-3 PUFA 比例由 14.39 降为 2.17。Zachut等[33]研究表明,添加亚麻籽组的血浆脂肪酸、脂肪组织、牛奶脂肪酸n-3 PUFA含量显著高于对照组,n-6/n-3 PUFA比例显著低于对照组,但是饲粮供给n-3 PUFA不能提高乳脂的ALA含量。这可能是因为脂肪酸经过瘤胃时,被瘤胃微生物氢化不饱和脂肪酸产生的脂肪酸异构体抑制乳脂中ALA生成。Farrel[34]在母鸡的日常饲粮中加入富含n-3 PUFA的鱼油,结果也显示鸡蛋中的n-6/n-3 PUFA比例从25.75下降至1.25。反刍动物由于瘤胃中的微生物群对饲粮中的n-3 PUFA进行广泛的生物降解作用,所以在反刍动物饲粮中不直接添加不饱和脂肪酸,而是添加过瘤胃保护性的不饱和脂肪酸或是富含n-3 PUFA鱼油外,再加入大豆粉、糖蜜等混合物一起作为反刍动物的饲粮,以保护n-3 PUFA免受降解作用。Kitessa等[35]报道,与对照组相比,食用添加保护性鱼油的奶牛可维持正常产奶量,且牛奶脂肪中n-3 PUFA含量可从8.49 g/kg 提高到 36.29 g/kg。Theurer等[36]研究表明,饲粮中添加多不饱和脂肪酸钙盐为泌乳期奶牛提供更多的必需脂肪酸,添加多不饱和脂肪酸钙盐组的牛奶脂肪的LA含量由3.96%增加至4.61%。通过添加亚麻籽可增加猪肉中n-3 PUFA含量,但是当ALA含量超过3%的中性脂肪或磷脂时,会对猪肉风味和货架保质期产生的负效应;加入鱼油量超过3%则会影响鸡蛋风味[37-38]。
5.2.2 通过饲草降低畜禽产品 n-6/n-3 PUFA比例
新鲜牧草中含有丰富的ALA,反刍动物采食牧草可使肉中ALA含量升高的同时降低n-6/n-3 PUFA比例,这是由于以谷物饲粮为基础的饲粮中LA是主要的脂肪酸,而ALA是牧草中的主要脂肪酸。LA、ALA分别是 n-6、n-3 PUFA的前体。Ponte等[39]研究限饲家禽采食以谷物为基础的饲粮自由放牧采食牧草对家禽生长性能和肉质的影响,结果表明采食牧草极显著的影响鸡肉中脂肪酸成分,尤其是胸肌肌肉中n-3 PUFA(ALA、EPA、DHA和 DPA),并且降低鸡肉中胆固醇含量。Razminowicz等[40]研究表明,采食牧草的肉牛与以精料喂养的肉牛相比,前者含有高水平的n-3 PUFA,同时 n-6/n-3 PUFA比例低于2∶1。Nuernberga等[41]研究不同的饲粮对牛背最长肌的多不饱和脂肪酸影响,试验牛分别饲喂牧草、精料,结果表明饲喂牧草的牛背最长肌n-3 PUFA含量显著增加,含量由 0.46%升高到 2.22%;n-6/n-3 PUFA比例由8.3降到2.0。牧草不同的收割时间、枯萎的牧草收割都会对其脂肪酸含量造成影响,进而影响反刍动物肉中的脂肪酸含量。
6 小结
合理的n-6/n-3 PUFA比例对机体健康有重要意义。通过改变饲粮可以有效地降低畜禽产品中的n-6/n-3 PUFA比例,提高畜禽产品品质,满足人类的健康需求。通过基因表达也可改善畜禽产品中的n-6/n-3 PUFA比例,但此技术尚在发展阶段,且还存在一些问题有待进一步研究。富含n-6、n-3 PUFA畜禽产品易氧化,添加抗氧化剂如维生素E可以防止其氧化,提高畜禽产品的货架期和保存期。但以下几方面的问题还有待进一步研究:1)多不饱和脂肪酸在畜禽产品中富集的规律;2)各种多不饱和脂肪酸源在不同畜禽饲粮类型中的最适添加量;3)畜禽产品中适宜的n-6/n-3 PUFA比例。
[1] SIMOPOULOSA P.Omega-6/omega-3 essential fatty acid ratio and chronic diseases[J].Food Reviews International,2004,20:77-90.
[2] NHMRC.Nutrient reference values for australia and new zealand,including recommended dietary intakes[R].[S.l.]:[s.n.],2006:37.
[3] GARG M L,LEITCH J,BLAKE R J,et a1.Long chain n-3 polyunsaturated fatty acid incorporation into human atrium following fish oil supplementation[J].Lipids,2006,41:1127-1132.
[4] BRENNER R R.Factors influencing fatty acid chain elongation and desaturation[M]//VERGROESEN A J.CRAWFORD M.The role of fats in human nutrition.New York:Academic Press,1989:45-80.
[5] SPRECHER H,LUTHRIA D L,MOHAMMED B S,et al.Reevaluation of the pathways for the biosynthesis of polyunsaturated fatty acids[J].Joural of Lipid,1995,36:2471-2477.
[6] KORVER D R,WAKENELL P,KLASING C.Dietary fish oil alters or lofrin,a 5-lipoxygenase inhibitor,decrease the growth-suppressing effects of coccidiosis in broiler chicks[J].Poultry Science,1997b,76:1355-1363.
[7] SIMOPOULOS A P.The role of fatty acids in gene expression:health implications[J].Annals of Nutrition and Metabalic,1996,40:303-311.
[8] MATER M K,THELEN A P,PAN D A,et al.Sterol response element-binding protein 1c(SREBP1c)is involved in the polyunsaturated fatty acid suppression of hepatic S14 gene transcription[J].Joural of Biological Chemitry,1999,274(46):32725-32732.
[9] DUPLUS E,FOREST C.Is there a single mechanism for fatty acid regulation of gene transcription[J].Biochemical Pharmcology,2002,64(5):893-901.
[10] VOLGAREV M N,LEVACHEV M M,TRUSHCHINA E N,et al.The modulation of the function of the immunocompetent system and of nonspecific body resistance in rats with different ratios of essential fatty acids in the diet[J].Europe PubMed Central,1993,116(12):607-609.
[11] PILEVAR M,ARSHAMI J,GOLIAN A,et al.Effects of dietary n-6∶n-3 ratio on immune and reproductive systems of pullet chicks[J].Poultry Science,2011,90:1758-1766.
[12] KOUBA M,MOUROT J.A review of nutritional effects on fat composition of animal products with special emphasis on n-3 polyunsaturated fatty acids[J].Biochimie,2011,93:13-17.
[13] GOMEZ-CORTEZ P,BACH A,LUNA P,et al.Effects of extruded linseed supplementation on n-3 fatty acids and conjugated linoleic acid in milk and cheese from ewes[J].Journal of Dairy Science,2009,92:4122-4134.
[14] RAES K,HAAK L,BALCAEN A,et al.Effect of linseed feeding at similar linoleic acid levels on the fatty acid composition of doublemuscled Belgian Blue young bulls[J].Meat Science,2004b,66:307-315.
[15] BAS P,SAUVANT D.Variations de la composition des dépõts lipidiques chez les bovins[J].Productions Animales,2001,14:311-322.
[16] WACHIRA A M,SINCLAIR L A,WILKINSON R G,et al.Effects of dietary fat source and breed on the carcass composition,n-3 polyunsaturated fatty acid and conjugated linoleic acid content of sheep meat and adipose tissue[J].British Journal of Nutrition,2002,88:697-709.
[17] MISSOTTEN J,DE SMET S,RAES K,et al.Effect of supplementation of the maternal diet with fish oil or linseed oil on fatty acid composition and expression of D5-and D6-desaturase in tissues of piglets[J].Animal,2009,3(8):1196-1204.
[18] MOUROT J,HERMIER D.Lipids in monogastric animal meat[J].Reproduction Nutrition Develpoment,2001,41:109-118.
[19] SHAPIRA N,WEILL P,LOEWENBACH R.Egg fortification with n-3 polyunsaturated fatty acids(PUFA):nutritional benefits versus high n-6 PUFA western diets,and consumer acceptance[J].The Israel Medical Association,2008,10:262-265.
[20] KOUBA M,BENATMANE F,BLOCHET J E,et al.Effect of a linseed diet on lipid oxidation,fatty acid composition of muscle,perirenal fat,and raw and cooked rabbit meat[J].Meat Science,2008,80:829-834.
[21] GUILLEVIC M,KOUBA M,MOUROT J.Effect of a linseed diet on lipid composition,lipid peroxidation and consumer evaluation of French fresh cooked pork meats[J].Meat Science,2009,81(4):612-618.
[22] CORTINAS L,BARROETA A,VILLAVERDE C,et al.Influence of the dietary polyunsaturation level on chicken meat quality:lipid oxidation[J].Poultry Science,2005,84:48-55.
[23] BOU R,CODONY R,TRES A,et al.Ditery strategies to improve nutritional value,oxidative stability and sensory properties of poultry products[J].Food Science and Nutrition,2009,49:800-822.
[24] BARROETA A C.Nutritive value of poultry meat:relationship between vitamin E and PUFA[J].World’s Poultry Science Association,2007,63:277-284.
[25] GOBERT M,MARTIN B,FERLAY A.Plant polyphenols associated with vitamin E can reduce plasma lipoperoxidation in dairy cows given n-3 polyunsaturated fatty acids[J].Journal of Dairy Science,2009,92:6095-6104.
[26] KANG JX,WANG J,WU L,et al.Transgenic mice:fat-1 mice convert n-6 to n-3 fatty acids[J].Nature,2004,427(6974):504.
[27] KAO B T,LEWIS K A,PETERS E J,et al.Endogenous production and elevated levels of long-chain n-3 fatty acids in the milk of transgenic mice[J].Journal of Dairy Science,2006,89:3195-3201.
[28] SMET D S,RAES K,DEMEYER D.Meat fatty acid composition as affected by fatness and genetic factors:a review[J].Animal Research,2004,53:81-98.
[29] KOUBA M,ENSER M,WHITTINGTON F M,et al.Effect of a high-linolenic acid diet on lipogenic enzyme activities,fatty acid composition,and meat qual-ity in the growing pig[J].Journal of Animal Science,2003,81(8):1967-1979.
[30] COLLINS C L,DAVIS J J,SINCLAIR A J,et al.Dietary flaxseed improves the fatty acid composition of lamb tissues[J].Asia Pacific Journal of Clinic Nutrition,2004,13(Suppl.):S52.
[31] BETTI M,PEREZ T I,ZUIDHOF M J,et al.Omega-3-enriched broiler meat:3.Fatty acid distribution between triacylglycerol and phospholipid classes[J].Poultry Science,2009,88:1740-1754.
[32] OLIVEIRA D D,BAIAO N C,CANCADO S V,et al.Effects of lipid sources in the diet of laying hens on the fatty acid profiles of egg yolks[J].Poultry Science,2010,89:2484-2490.
[33] ZACHUT M,ARIELI A,LEHRER H,et al.Effects of increased supplementation of n-3 fatty acids to transition dairy cows on performance and fatty acid profile in plasma,adipose tissue,and milk fat[J].Journal of Dairy Science,2010,93:5877-5889.
[34] FARREL D J.Enrichment of hen eggs with n-3 longchain fatty acids and evaluation of enriched eggs in humans[J].The American Journal of Clinical Nutrition,1998,68(3):538-544.
[35] KITESSA S M,GULATI S K,SIMOS G C,et al.Supplementation of grazing dairy cows with rumenprotected tuna oil enriches milk fat with n-3 fatty acids without affecting milk production or sensory characteristics[J].British Journal of Nutrition,2004,91(2):271-278.
[36] THEURER M L,BLOCK E,SANCHEZ W K,et al.Calcium salts of polyunsaturated fatty acids deliver more essential fatty acids to the lactating dairy cow[J].Journal of Dairy Science,2009,92(5):2051-2056.
[37] MARSHALL A C,KUBENA K S,HINTON K R,et al.n-3 fatty acid enriched table eggs:a survey of consumer acceptability[J].Poultry Science,1994a,73:1334-1340.
[38] SIMOPOULOS A P.The omega-6/omega-3 ratio:the scientific evidence and the need to return the omega-3 fatty acids into eggs and other foods[J].Nature’s Perfect Functional Food for Health Promotion,2006,20:195-218.
[39] PONTE P I P,PRATES J A M,CRESPO J P,et al.Restricting the intake of a cereal-based feed in freerange-pastured poultry:effects on performance and meat quality[J].Poultry Science,2008,87:2032-2042.
[40] RICHARDSON R I,WOOD J D,SCOLLAN N D,et al.Quality of meat from biodiverse grassland[C]//HOPKINS J J,DUNCAN A J,MCCRACKEN D I,et al.High value grassland:providing biodiversity,a clean environment and premium products.Staffordshire UK:Keele University,2007:107-116.
[41] NUERNBERGA K,DANNENBERGERA D,NUERNBERGA G,et al.Effect of a grass-based and a concentrate feeding system on meat quality characteristics and fatty acid composition of longissimus muscle in different cattle breeds[J].Livestock Production Science,2005,94:137-147.
