北京松山自然保护区森林群落物种多样性及其神经网络预测
2013-12-16苏日古嘎张金屯王永霞
苏日古嘎,张金屯 ,王永霞
(1.内蒙古师范大学生命科学与技术学院,呼和浩特 010022;2.北京师范大学生命科学学院,北京 100875;3.内蒙古交通职业技术学院道路与桥梁工程系,赤峰 024005)
物种多样性是群落结构和功能复杂性的一种度量[1]。物种多样性的空间分布格局受许多环境因子的影响,它沿环境梯度的变化规律是多样性研究的一个重要方面[2]。研究植物群落的物种多样性,有助于更好地认识群落的组成、变化和发展趋势,同时也可以反映群落及其环境的保护状况,所以物种多样性及其与生态环境关系的研究已成为生态学关注的热点[3]。北京松山自然保护区以油松林和次生落叶阔叶林为主,森林群落保存较为完整,种类丰富、结构复杂是该区群落最为突出的特点。对该区的植被动态[4-5]、油松种群结构[6-7]、生态旅游[8-9],净化服务功能的生态经济价值[10]等方面已有一些研究,但对于松山森林群落物种多样性及其预测的研究尚未见报道。
本文以松山自然保护区森林群落为对象,运用多样性指数,多层感知器网络综合分析多样性与群落类型、结构和生境之间的关系,从而加深对该保护区森林群落结构、性质的认识,以期为该保护区的生物多样性保护提供科学依据。
1 研究方法
1.1 研究区自然概况
松山自然保护区位于北京市延庆县西北部海陀山南麓,地理坐标为115°43′44″—115°50′22″(E,East),40°29′9″—40°33′35″(N,North),北依主峰海陀山,海拔2198.4 m,为北京地区第二高峰,最低海拔 627.6 m。保护区建立于1985年,1986年晋升为国家级自然保护区,属森林和野生动物类型的自然保护区,保护区总面积4671 hm2,有林地面积3655 hm2,森林覆盖率87.6%。松山处于暧温带大陆性季风气候区,平均气温8.5℃,最高温度39℃,最低温度-27.3℃,年平均日照2836.3 h,年降水量493 mm,年蒸发量1772 mm,气候垂直分带性比较明显。土壤随海拔由低向高依次有山地褐土、山地棕壤、山地草甸土。保存有华北地区唯一的大片珍贵天然油松(Pinus tabulaeformis)林,以及保存良好的胡桃楸(Juglans mandshurica)、椴(Tilia spp.)、榆(Ulmus spp.)、桦(Betula spp.)等树种构成的华北地区典型的天然次生落叶阔叶林[4,11]。
1.2 样方设置与调查
于2008年7—8月在北京松山自然保护区沿海拔梯度取样,从海拔700 m到2000 m,每隔100 m设一条样带,在每条样带上设置2—6个面积10 m×10 m的样方,共计68个森林样方。在每个样方中,样方的正中心位置用全球定位系统(GPS)测定海拔高度、用罗盘测定坡度和坡向;用Dush-cone A-1820仪器测定土壤紧实度,10 m2内测定一个土壤紧实度值,在每个10 m×10 m样方内测定10个重复;用土钎和卷尺测定土壤深度和枯枝落叶层厚度,20 m2内测定一个土壤紧实度值,在每个10 m×10 m样方内测定5个重复。同时每个样方内记录每个物种的盖度、高度,乔木要记录株数、胸径和冠幅等数据。
1.3 数据处理
1.3.1 重要值
分别计算乔木、灌木和草本植物重要值;其中灌木和草本植物有的丛生,有的单生,个体差异较大,而其盖度能更好地反映实际情况,所以只用相对高度和相对盖度来计算其重要值;计算公式为[12-15]:
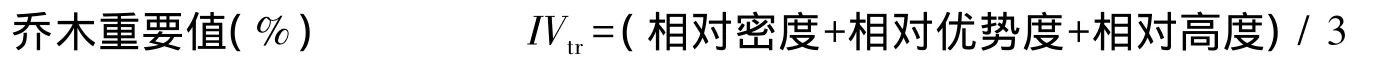
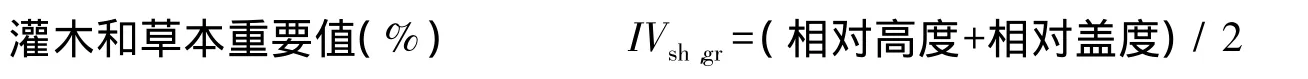
式中,IVtr为乔木重要值;IVsh,gr为灌木和草本重要值。
1.3.2 环境数据
包括6个环境因子,即海拔高度、坡度、坡向、枯枝落叶层厚度、土壤深度、土壤紧实度,涵盖了研究植被类型的全部的分布范围以及样方分布情况。土壤紧实度用土壤耐压力表示:
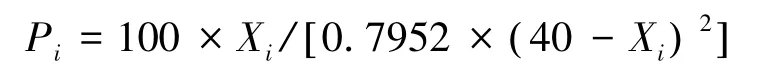
式中,Xi为紧实度仪的读数(mm);Pi为土壤耐压力(kg/m2)。
为了便于建立环境数据矩阵,将坡向数据以等级制表示;将坡向按照45°的夹角,以北为0°,顺时针方向旋转分为8个坡向等级,以数字表示各等级:1表示北坡(0°—22.5°,337.5°—360°),2表示东北坡(22.5°—67.5°),3 表示西北坡(292.5°—337.5°),4 表示东坡(67.5°—112.5°),5 表示西坡(247.5°—292.5°),6 表示东南坡(112.5°—157.5°),7 表示西南坡(202.5°—247.5°),8 表示南坡(157.5°—202.5°),数字越大,表示坡向越向阳[16-18]。
1.4 多样性指数的测定
森林群落植物分为乔木、灌木和草本等功能层,功能层反映群落结构的重要性[16]。用重要值数据分别计算3个功能层的Patrick丰富度指数、Shannon-Wiener物种多样性指数和Pielou均匀度指数。计算公式如下[12,16]:
(1)丰富度指数

式中,S为每一样方中的物种总数。
(2)物种多样性指数

式中,N为S个种的全部重要值之和;Ni为第i个种的重要值。
(3)均匀度指数

1.5 群落物种多样性的测定
采用群落分层物种多样性测度法,将群落中的乔木层、灌木层、草本层的多样性指数分别计算后,给定加权参数计算群落总体多样性指数,公式如下[19]:

式中,D为群落总体指数;D1、D2、D3分别为乔木层、灌木层、草本层的多样性指数;W1、W2、W3分别为给定乔木层、灌木层、草本层的权重系数,这里采用 0.5、0.25、0.25[19]。
1.6 多层感知器网络的原理与方法
多层感知器(MLP)网络具有独特的学习算法——误差反向传播算法(BP算法),所以也被称之为BP网络。在生态学和农业研究中,MLP网络模型通常作为非线性函数模拟器用于预测作物产量、生物生产量、生物与环境之间的关系等[20-21],还应用于预测物种多度[22]、物种多样性[23]。它由输入层、隐含层和输出层组成(图1)[22],其中隐含层可以为一层或多层隐单元,隐单元从输入模式中提取更多有用的信息,使网络可以完成更复杂的任务。输入层神经元的个数为输入信号的维数,隐含层个数以及隐含层节点个数视具体情况而定,输出层神经元的个数为输出信号的维数,每一层的神经元个数是MLP网络一个重要的参数[23]。输入层含独立变量,每个输入层有对应的输出层,所以通过输入层能预测输出层信息。

图1 多层感知器模型Fig.1 Multilayer perceptron neural network model
在本文中,海拔高度、坡度、坡向、枯枝落叶层厚度、土壤深度,土壤紧实度等6个环境因子为输入层,分别把乔木层、灌木层、草本层的Shannon-Wiener物种多样性指数为输出层。通过多次训练MLP网络,确定隐含层个数,选择均方误差最小的网络[24]。预测乔木层和灌木层的Shannon-Wiener指数时选择输入层6、隐含层11、输出层1(6-11-1结构)的MLP网络,而预测草本层的Shannon-Wiener指数时选择输入层6、隐含层9、输出层1(6-9-1结构)的MLP网络。完成学习过程后,可以计算观察值和预测值之间的相关性,检验网络的可预测性。用敏感性分析来决定MLP网络中输入层对输出层的贡献。输入层的每个变量加减[-0.1,0.1]至[-0.5,0.5]的随机数,计算每个输入层变量在不同干扰水平下的均方误差,估量输出层的反映[24-25]。
在Matlab7.0.1平台上,NNToolbox工具箱里完成分析计算。
2 结果与分析
2.1 不同群落类型的物种多样性分析
图2显示了松山7个森林群落类型的物种丰富度、多样性和均匀度的分布变化图,反映了松山不同森林群落类型在物种组成上的差异。从图2中看出,群落Ⅱ和Ⅴ分别是大果榆+山杨混交林、油松+青杨混交林,群落结构复杂、发育较好,其物种丰富度、物种多样性和均匀度均较高,这说明混交林的群落物种多样性比其他群落的高。而群落Ⅲ和Ⅶ分别为大果榆林、华北落叶松林,群落结构简单,因而各项指数值均较低。
在一个特定的地带性气候内,植物群落垂直结构受群落所处的海拔、坡向、坡位等微环境、群落的种类组成、发育阶段、生活史对策的影响,因此群落生长型的功能层,乔木层、灌木层、草本层物种多样性变化各有其特点。从图3中看出,群落Ⅶ中没有灌木层,只有乔木层和草本层。Patrick指数在森林群落中表现为草本层>灌木层>乔木层;Shannon-Weiner指数在森林群落中表现为草本层>灌木层>乔木层,与森林群落丰富度指数排列顺序一致;Pielou指数在榆林中表现为草本层>乔木层>灌木层,而在其他森林群落中表现为灌木层>草本层>乔木层。
2.2 功能层的物种多样性分析
从松山森林群落典范对应分析(CCA)得出海拔是影响森林群落和功能层的重要因子之一[26]。因此,分析了森林群落功能层多样性在海拔梯度上的变化趋势。图4—图6分别为松山自然保护区森林群落乔木层、灌木层和草本层的Patrick指数、Shannon-Wiener指数和Pielou指数在海拔梯度上的变化趋势及多项式拟合效果。
在乔木层,丰富度、多样性和均匀度随海拔的升高逐渐降低。海拔高度和Patrick指数、Shannon-Wiener指数、Pielou指数拟合的相关系数平方(R2)分别为0.189、0.2582、0.3074,均呈极显著相关(图4)。
在灌木层,丰富度、多样性和均匀度都呈比较明显的单峰曲线变化趋势,即随海拔的升高先增加后逐渐降低。海拔高度和Patrick指数、Shannon-Wiener指数、Pielou指数拟合的R2分别为0.179、0.2363、0.142,均呈极显著相关(图5)。
在草本层,丰富度和多样性随海拔的升高都呈下降趋势,而均匀度变化不大。Patrick指数、Shannon-Wiener指数拟合的R2分别为0.1397、0.0818,均呈显著相关,而Pielou指数拟合的R2为0.0122,与海拔无显著的相关(图6)。

图2 松山自然保护区森林群落的物种多样性趋势图Fig.2 The tendency of the species diversity of the forest communities in Songshan Nature Reserve

图3 松山自然保护区功能层的物种多样性趋势图Fig.3 The tendency of the species diversity of functional groups in Songshan Nature Reserve
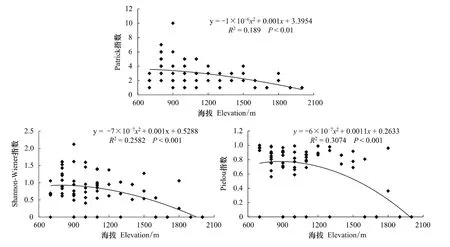
图4 森林群落乔木层丰富度指数、多样性和均匀度指数在海拔梯度上的变化趋势Fig.4 Variations of richness,diversity and eveness indices of tree layer of forest communities along elevational gradient
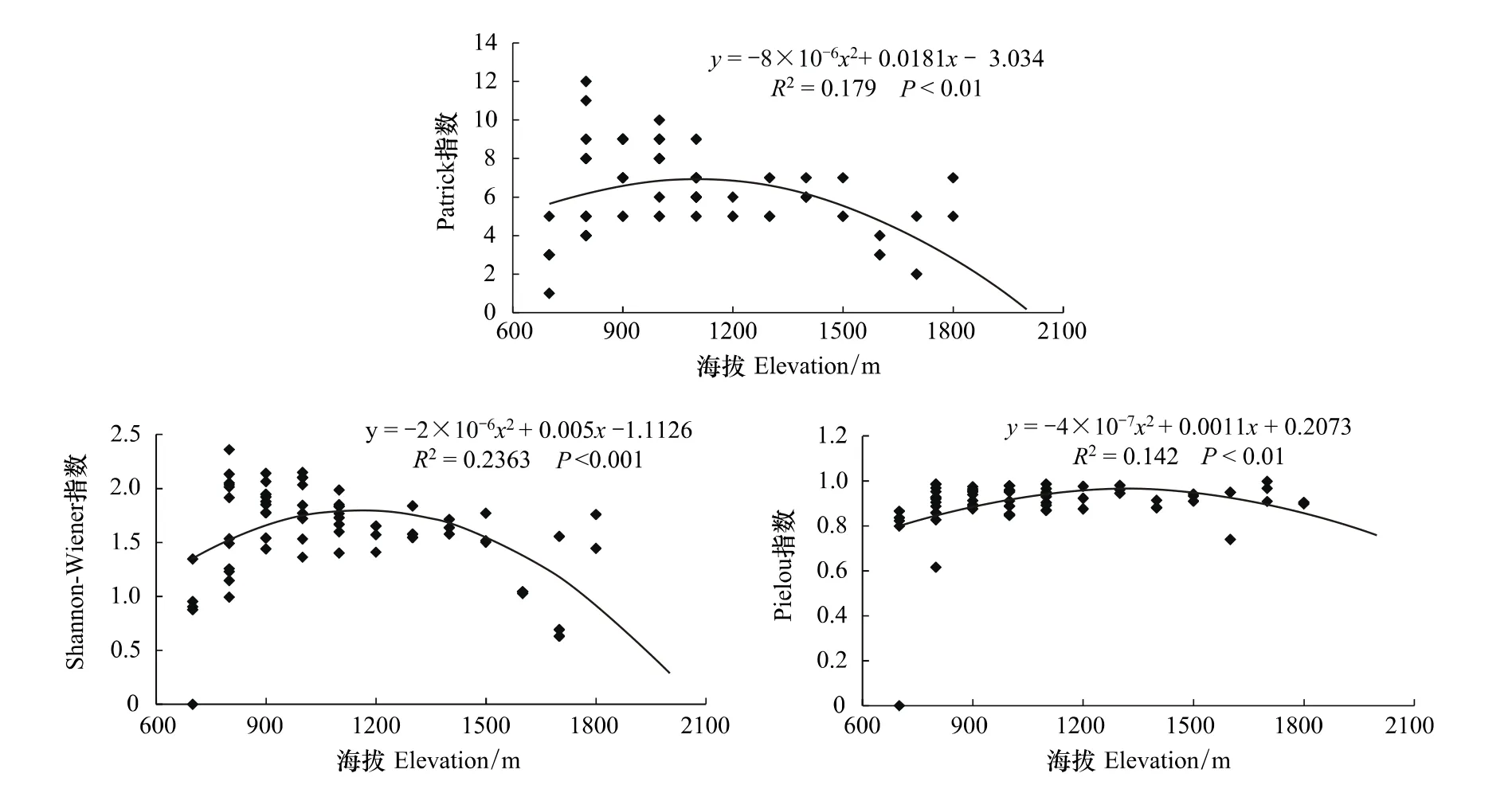
图5 森林群落灌木层丰富度指数、多样性和均匀度指数在海拔梯度上的变化趋势Fig.5 Variations of richness,diversity and eveness indices of shrub layer along elevational gradient
2.3 用MLP网络预测功能层的物种多样性
用MLP网络预测物种多样性,用海拔高度、坡度、坡向、枯枝落叶层厚度、土壤深度和土壤紧实度等6个环境因子作为输入层,Shannon-Wiener指数为输出层。在乔木层,用MLP网络训练,迭代100次,平方误差为2.75×10-29;在灌木层,迭代100次,平方误差为7.7×10-29;在草本层,迭代100次,平方误差为7.01×10-6。MLP训练网络显示通过环境因子预测物种多样性准确性高,在乔木层、灌木层、草本层,R均达到1(图7)。
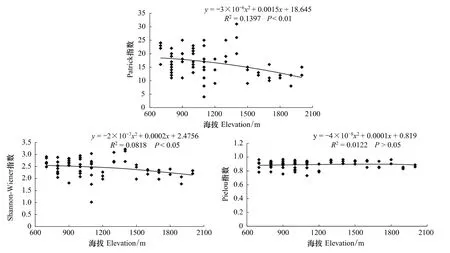
图6 森林群落草本层丰富度指数、多样性和均匀度指数在海拔梯度上的变化趋势Fig.6 Variations of richness,diversity and eveness indices of herb layer along elevational gradient

图7 在MLP训练网络中观测值和预测值相关性散点图,斜直线代表最佳预测结果Fig.7 Scatter plots of correlations between observed and predicted values by the trained MLP.The diagonal lines represent perfect prediction values
通过敏感性分析,评价每个输入变量对MLP训练网络的影响。在每个输入变量上加减[-0.1,0.1]至[-0.5,0.5]之间的随机数,计算输入变量不同干扰水平下的均方误差。在乔木层,坡向对乔木层物种多样性的影响最大,其次是土壤紧实度、海拔高度、土壤深度、坡度,枯枝落叶层厚度;在灌木层,坡向对灌木层物种多样性的影响最大,其次是枯枝落叶层厚度、土壤紧实度、海拔高度、坡度,土壤深度;在草本层,海拔高度对草本层物种多样性的影响最大,其次是土壤紧实度、坡向、土壤深度、枯枝落叶层厚度、坡度(图8)。
3 结论与讨论
松山自然保护区内的森林群落物种多样性反映了该区植被类型在物种组成上的差异。大果榆+山杨混交林、油松+青杨混交林物种丰富度、物种多样性和均匀度均较高,而大果榆林、华北落叶松林的各项指数值均较低,这与群落结构复杂程度相关,华北落叶松林没有幼树和幼苗,缺乏灌木层。Patrick指数在森林群落中表现为草本层>灌木层>乔木层,这一结果与其他森林群落的物种多样性研究结果较为一致[2,27];Shannon-Weiner指数在森林群落中表现为草本层>灌木层>乔木层;Pielou指数在榆林中表现为草本层>乔木层>灌木层,而在其他森林群落中表现为灌木层>草本层>乔木层。
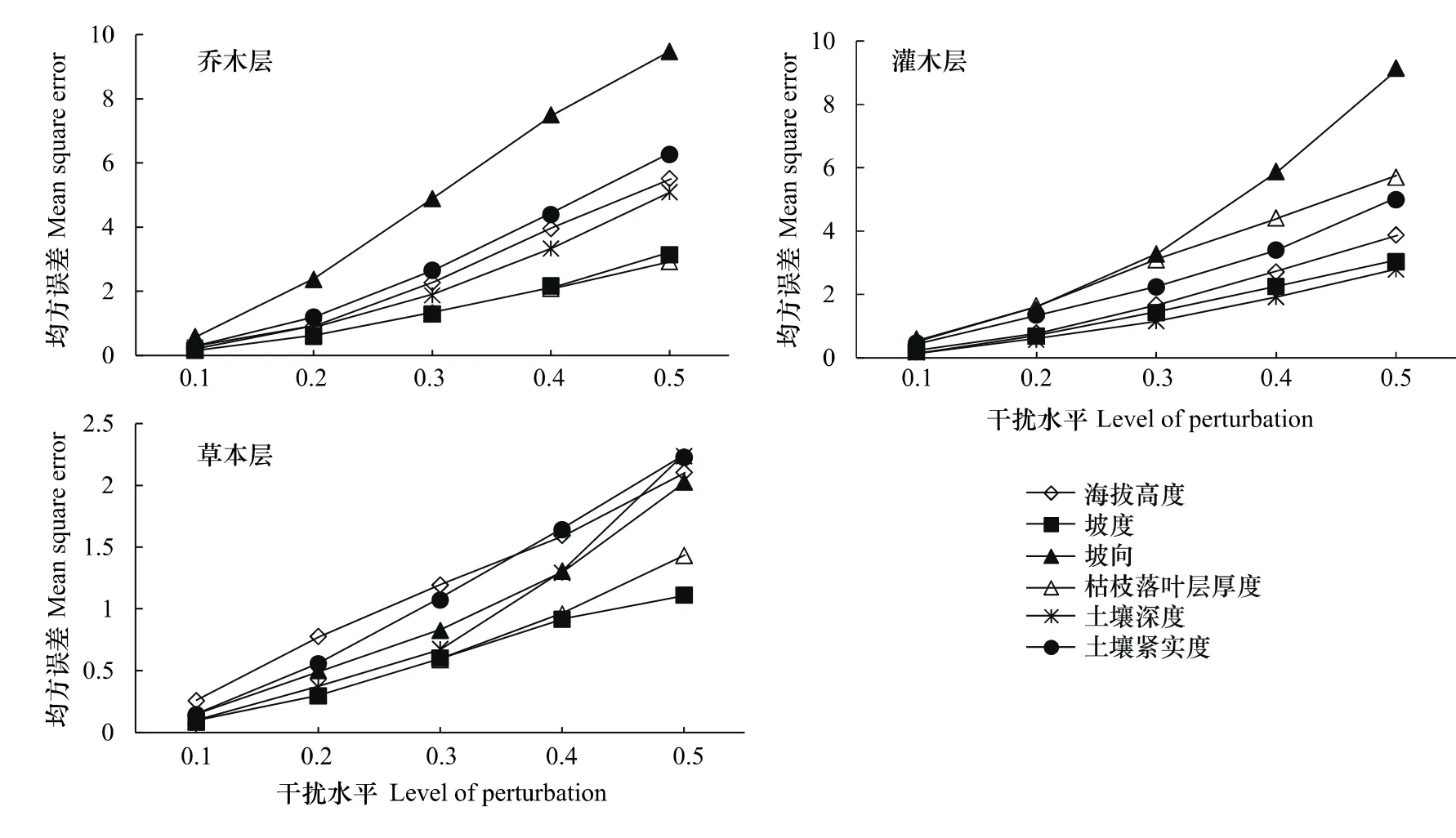
图8 MLP网络的敏感性分析,在不同干扰水平下计算的均方误差Fig.8 The sensitivity analysis of MLP,mean square error values were measured at different levels of perturbation of the input variables
山地海拔高度是影响物种多样性格局的决定性因素[19],主要反映水热梯度变化,包含了温度、湿度和光照等各种环境因素。从平地到高山山顶,气候条件差异很大,通常海拔每升高100m,气温大约要下降0.5—1℃左右,温度、风力、光照和其他气候因子及其配合方式也会产生相应的变化,这些变化都会对群落的物种多样性产生影响[28]。在乔木层,丰富度、多样性和均匀度随海拔的升高逐渐降低,说明乔木层的物种多样性受高海拔地区热量限制,从而形成高海拔地区物种多样性降低的趋势。在灌木层,丰富度、多样性和均匀度呈比较明显的单峰曲线变化趋势,即随海拔的升高先增加后逐渐降低;在草本层,丰富度和多样性随海拔的升高都呈下降趋势,这与中间海拔高度多样性最高的假设相一致[16]。而在草本层,均匀度变化不大。
用MLP网络预测功能层的物种多样性,在乔木层,坡向对乔木层物种多样性的影响最大,其次是土壤紧实度,影响力较小的是枯枝落叶层厚度;在灌木层,坡向对灌木层物种多样性的影响最大,其次是枯枝落叶层厚度,影响较小的是土壤深度;坡向是通过改变光照、温度、水分和土壤等生态因子而对乔木层和灌木层多样性产生重要影响。乔木根系分布到土壤的深层,枯枝落叶层厚度对乔木层多样性的影响力较小,而因灌木根系相对较浅,所以受土壤深度的影响较小。在草本层,海拔高度对草本层物种多样性的影响最大,其次是土壤紧实度,影响力较小的是坡度,说明草本层物种多样性受海拔水热梯度变化影响较大,而坡度对林下的草本植物影响不大。MLP网络应用于预测昆虫物种多度[29]、物种多样性[30]方面较多,通过本文的分析,MLP网络可以应用于预测森林群落多样性。人工神经网络处理复杂数据的上显示优势,有待于进一步探索和挖掘MLP网络在评估、理解、预测和管理生物多样性中的应用。
虽然松山自然保护区的森林群落分布格局主要受海拔梯度的影响,导致植被类型的垂直分布,但影响森林群落功能层物种多样性差异的因子具有明显的区别,因此,今后对该保护区内的森林群落进行管理时,既要注重群落分布格局,也要注重各功能层物种多样性受影响的环境因子,从而达到保护森林群落及其物种多样性的双重目标。
[1] Bo Y J,Zhang X,Ai H J,Liu C Y,Ji X L.Species diversity in the wind-sandy grass shoal area of Yulin region.Bulletin of Soil and Water Conservation,2008,28(4):80-85.
[2] Xu B,Zhang J T,Yang H X,Jiang H F.Species diversity of Baihua mountain forest plant community.Bulletin of Botanical Research,2007,27(1):112-118.
[3] Zhang L,Xing F,Yu L L,Xu K,Sun Z L,Lü X G.Plant species diversity of the island forest in a marsh in the Jiang plain,China.Journal of Plant Ecology,2008,32(3):582-590.
[4] Zhou R,Wang H,Ge J P,Xiong Y C,Wu J G.Vegetation dynamics and patterns of change in functional protection subzones in Songshan National Nature Reserve,Beijing.Biodiversity Science,2006,14(6):470-478.
[5] Zhou R,Ge J P,Yu B,Liu L J,Wu J G.Simulation of forest dynamics at Songshan Mountain,Beijing.Journal of Beijing Forestry University,2007,29(1):19-25.
[6] Niu L L,Yu X X,Yue Y J.Spatial patterns of different age-class individuals in Pinus tabulaeformis forest in Songshan Nature Reserve of Beijing,China.Chinese Journal of Applied Ecology,2008,19(7):1414-1418.
[7] Zhang B,Zhao Y Z,Zhang C Y,Zhao X H.Structure and spatial distribution of Pinus tabulaeformis population in the Songshan Nature Reserve,Beijing,China.Chinese Journal of Applied and Environmental Biology,2009,15(2):175-179.
[8] Chen J,Zhu D D,Liu M L.Research on the environmental education of ecotourism in Songshan Nature Reserve.Sichuan Forestry Exploration and Design,2008,(3):38-41.
[9] Chen L,Wang W.Research on the relationship between tourism development and ecological protection in Songshan Nature Reserve.Sichuan Forestry Exploration and Design,2009,(1):41-43.
[10] Wu S,Qin D H,Bi X L,Wang X C,Ge J P.Purifying service of Songshan mountain and its value.Journal of Beijing Normal University:Natural Science,2004,40(3):386-389.
[11] Niu J,Zhang Y J.Analysis on the model of ecotourism management and local community participation of Beijing Songshan Mountain Nature Reserve.Sichuan Forestry Exploration and Design,2007(2):40-43.
[12] Zhang J T.Quantitative Ecology.Beijing:Science Press,2004.
[13] He H Q,Li S C,Sun H L,Liu S C,Xiong W L.Quantitative classification and ordination of Jinping hydropower station,Sichuan Province,China.Acta Ecologica Sinica,2008,28(8):3706-3712.
[14] Dong L S,Zhang X D,Zhou J X,Song A Y.Quantitative classification and ordination of shrub species and communities in a loess landscape of western Shanxi.Acta Ecologica Sinica,2007,27(7):3072-3080.
[15] Zhang X P,Wang M B,She B,Xiao Y.Numerical classification and ordination of forest communities in Pangquangou National Nature Reserve.Acta Ecologica Sinica,2006,26(3):754-761.
[16] Zhang J T,Zhang F.Diversity and composition of plant functional groups in mountain forests of the Lishan Nature Reserve,North China.Botanical Studies,2007,48(3):339-348.
[17] Zhang W Q,Li X H,Luo Q Z,Zhang W M,Zhao J,Shan Y B.Spatial distribution of vegetation in Tianmu Mountain Nature Reserve based on RS and GIS data.Chinese Journal of Ecology,2003,22(6):21-27.
[18] Shen Z H,Fang J Y.Niche comparison of two Fagus species based on the topographic patterns of their populations.Acta Phytoecologica Sinica,2001,25(4):392-398.
[19] Zhang F,Zhang J T,Shangguan T L.Plant diversity of forest community in Zhuweigou of Lishan mountain Nature Reserve.Acta Phytoecologica Sinica,2002,26(supplement):46-51.
[20] Lek S,Guégan J F.Artificial neural networks as a tool in ecological modelling,an introduction.Ecological Modelling,1999,120(2/3):65-73.
[21] Mi X C,Ma K P,Zou Y B.Artificial neural network and its application in agricultural and ecological research.Acta Phytoecologica Sinica,2005,29(5):863-870.
[22] Acharya C,Mohanty S,Sukla L B,Misra V N.Prediction of sulphur removal with Acidithiobacillus sp.using artificial neural networks.Ecological Modelling,2006,190(1/2):223-230.
[23] Ge Z X,Sun Z Q.The Theory of the Neural Network and MATLAB R2007 Application.Beijing:Publishing House of Electronics Industry,2007.
[24] Scardi M,Harding L W Jr.Developing an empirical model of phytoplankton primary production:a neural network case study.Ecological Modelling,1999,120(2/3):213-223.
[25] Park Y S,Céréghino R,Compin A,Lek S.Applications of artificial neural networks for patterning and predicting aquatic insect species richness in running waters.Ecological Modelling,2003,160(3):265-280.
[26] Suriguga,Zhang J T,Zhang B,Cheng J J,Tian S G,Zhang Q D,Liu S J.Numerical classification and ordination of forest communities in the Songshan National Nature Reserve.Acta Ecologica Sinica,2010,30(10):2621-2629.
[27] Tang Z Y,Fang J Y,Zhang L.Patterns of woody plant species diversity along environmental gradients on Mt.Taibai,Qinling Mountains.Biodiversity Science,2004,12(1):115-122.
[28] Xie J Y,Chen L Z.Species diversity characteristics of deciduous forests in the warm temperate zone of north China.Acta Ecologica Sinica,1994,14(4):337-344.
[29] Céréghino R,Park Y S,Compin A,Lek S.Predicting the species richness of aquatic insects in streams using a limited number of environmental variables.Journal of the North American Benthological Society,2003,22(3):442-456.
[30] Lek-Ang S,Deharveng L,Lek S.Predictive models of collembolan diversity and abundance in a riparian habitat.Ecological Modelling,1999,120(2/3):247-260.
参考文献:
[1] 耀军,张雄,艾海舰,刘翠英,纪晓玲.榆林风沙草滩区物种多样性研究.水土保持通报,2008,28(4):80-85.
[2] 许彬,张金屯,杨洪晓,姜海凤.百花山植物群落物种多样性研究.植物研究,2007,27(1):112-118.
[3] 张亮,邢福,于丽丽,许坤,孙忠林,吕宪国.三江平原沼泽湿地岛状林植物多样性.植物生态学报,2008,32(3):582-590.
[4] 周睿,王辉,葛剑平,熊友才,吴记贵.松山自然保护区各功能区植被动态及变化格局.生物多样性,2006,14(6):470-478.
[5] 周睿,葛剑平,于波,刘丽娟,吴记贵.北京松山森林动态模拟研究.北京林业大学学报,2007,29(1):19-25.
[6] 牛丽丽,余新晓,岳永杰.北京松山自然保护区天然油松林不同龄级立木的空间点格局.应用生态学报,2008,19(7):1414-1418.
[7] 张赟,赵亚洲,张春雨,赵秀海.北京松山油松种群结构及空间分布格局.应用与环境生物学报,2009,15(2):175-179.
[8] 陈静,朱丹丹,刘明丽.松山自然保护区生态旅游环境教育研究.四川林勘设计,2008,(3):38-41.
[9] 陈亮,王雯.北京松山自然保护区旅游开发与生态保护关系的探讨.四川林勘设计,2009,(1):41-43.
[10] 吴姗,覃德华,毕晓丽,王秀春,葛剑平.松山自然保护区净化服务功能及生态经济价值研究.北京师范大学学报:自然科学版,2004,40(3):386-389.
[11] 牛江,张玉钧.北京松山自然保护区生态旅游管理与当地社区参与.四川林勘设计,2007(2):40-43.
[12] 张金屯.数量生态学.北京:科学出版社,2004.
[13] 何惠琴,李绍才,孙海龙,刘世昌,熊文兰.锦屏水电站植被数量分类与排序.生态学报,2008,28(8):3706-3712.
[14] 董林水,张旭东,周金星,宋爱云.晋西黄土丘陵与土石山区交错地带灌木种的数量分类与排序.生态学报,2007,27(7):3072-3080.
[15] 张先平,王孟本,佘 波,肖扬.庞泉沟国家自然保护区森林群落的数量分类和排序.生态学报,2006,26(3):754-761.
[17] 章皖秋,李先华,罗庆州,张卫民,赵军,单勇兵.基于RS、GIS的天目山自然保护区植被空间分布规律研究.生态学杂志,2003,22(6):21-27.
[18] 沈泽昊,方精云.基于种群分布地形格局的两种水青冈生态位比较研究.植物生态学报,2001,25(4):392-398.
[19] 张峰,张金屯,上官铁梁.历山自然保护区猪尾沟森林群落植物多样性研究.植物生态学报,2002,26(增刊):46-51.
[23] 葛哲学,孙志强.神经网络理论与MATLAB R2007实现.北京:电子工业出版社,2007.
[26] 苏日古嘎,张金屯,张斌,程佳佳,田世广,张钦弟,刘素军.松山自然保护区森林群落的数量分类和排序.生态学报,2010,30(10):2621-2629.
[27] 唐志尧,方精云,张玲.秦岭太白山木本植物物种多样性的梯度格局及环境解释.生物多样性,2004,12(1):115-122.
[28] 谢晋阳,陈灵芝.暖温带落叶阔叶林的物种多样性特征.生态学报,1994,14(4):337-344.
