象山港春季网采浮游植物的分布特征及其影响因素
2013-12-16江志兵朱旭宇廖一波曾江宁
江志兵,朱旭宇,高 瑜,廖一波,寿 鹿,曾江宁,* ,黄 伟
(1.国家海洋局海洋生态系统与生物地球化学重点实验室,国家海洋局第二海洋研究所,杭州 310012;2.应用海洋生物技术教育部重点实验室,宁波大学海洋学院,宁波 315211)
象山港是浙江省最重要的海水增养殖基地和渔业资源产地,也是浙江省发展海洋经济重要的天然资源,因其独特的地理区位和资源优势,近年来掀起了前所未有的海洋开发热潮,如大型火电厂运行、水产养殖、临港工业发展和滩涂围垦等。然而,由于象山港与外海的水体交换周期较长[1],港内海洋自净能力弱、环境容量小、生态脆弱,沿港高强度的建设开发对海域生态环境造成极大的压力。目前,象山港海洋开发活动引起的水动力学改变、富营养化和水温上升等现象必然造成海洋生态环境的改变,并由此引起了相关研究者的注意,包括水动力[2]、营养盐[3-5]、微生物[6-7]、浮游动物[8-10]和底栖生物[11-14]等。浮游植物作为生态系统中最主要的初级生产者,可为浮游动物、甲壳动物、软体动物和仔稚鱼等提供食物和必需的营养成分,且其分布状况与水动力、营养盐和温度等环境因子密切相关[15-17]。虽然早期也有象山港浮游植物的调查[1,18-20],但近年来报道较少[21-22],且未有多种海洋开发活动联合作用对其影响的研究。本文根据2010年春季象山港网采浮游植物调查资料,选取相关环境因子,通过典范对应分析(CCA),确定影响该海域浮游植物群落的主要因素,并与同期历史数据比较,探讨海洋开发活动对浮游植物的影响,为象山港海洋经济和环境保护政策的制订提供基础资料和科学依据。
1 研究区域与方法
1.1 研究区域和站位布设
象山港地处浙江北部沿海,是一个纵长约为62.8 km的狭长型半封闭海湾[18],总面积为563 km2,其中水域面积约占70%,滩涂面积约占30%,水动力条件较弱,港中部和港底90%水体被置换到外海的时间分别为65 d和80 d[1]。本次调查共设28个站位(图1),调查范围自港底至口门外的大目洋(121.47—122.13°E,29.42—29.75°N),测站最大水深29.0 m,平均测站水深11.2 m。根据象山港的地理位置,可将其分为支港:铁港(S1—S3)、黄墩港(S7和S8)和西沪港(S20和S21);主港:港底(S4—S6、S9和S10,邻近宁海电厂)、港中部(S11—S19,邻近乌沙山电厂)、港口(S22—S24)和港口外(S25—S28,大目洋)等7个区块。为便于分析,将铁港、黄墩港和主港港底归为全港底部,将主港中部和西沪港归为全港中部,主港港口即为全港港口,这些区块(S1—S24)均属于象山港港内。
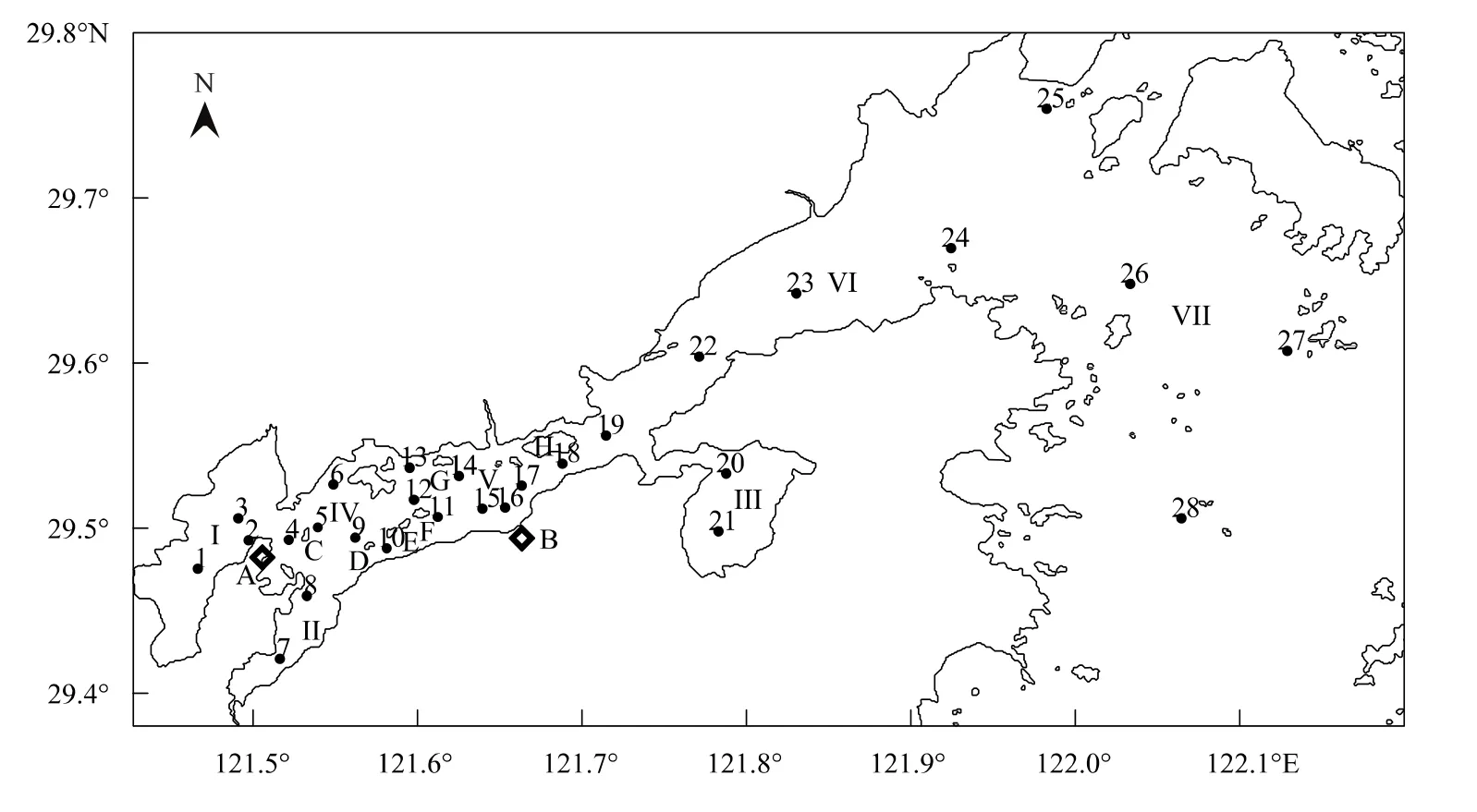
图1 调查站位Fig.1 Sample stations in the Xiangshan Bay
1.2 样品采集及分析
于2010年春季(4月23日—24日)小潮期间在各站用浅Ⅲ浮游生物网自底至表作垂直拖网采集浮游植物样品28个,用Hydro-bios流量计记录滤水量。样品用2%中性甲醛固定,经浓缩后用Leica DM2500显微镜观察、鉴定和计数。所有操作均按《海洋调查规范》[23]进行。同时采集表层(0.5 m)水样,测定其温度、盐度、pH值、溶解氧(DO)、溶解无机氮、活性磷酸盐、硅酸盐和悬浮物浓度,并记录水深和透明度。
1.3 数据处理
用PRIMER 5.0版软件计算浮游植物群落的Shannon-Weiner多样性指数(H′)和Pielou均匀度指数(J′)。优势度(Y)计算公式为:
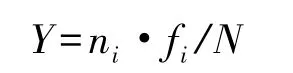
式中,N为样品的总丰度;ni为样品中第i种的丰度;fi为该种浮游植物在样品中的出现频率。若某物种的Y≥0.01,则认定为优势种。相关参数(或经log转换后)如满足正态分布(K-S检验)和方差齐次性 (Levene检验),用SPSS 13.0软件对其进行单因子方差分析(One-way ANOVA),反之则对变量进行非参数检验(K-W检验)。K-W检验若发现各组间呈显著差异,可利用SPSS软件自编程序进行多重比较Nemenyi秩和检验。用PRIMER软件对f≥10%的物种丰度进行log(x+1)转换后,建立Bray-Curtis相似性聚类分析,并采用非度量多维尺度分析(nMDS)分析浮游植物群落的空间分布。浮游植物群落区域间比较采用单因素相似性分析(ANOSIM)。用CANOCO 4.5软件进行CCA排序,用于排序的物种在各站位的f≥10%,且其丰度占总丰度的比例需≥0.2%[16,24]。
2 结果与分析
2.1 种类组成与生态类群
象山港春季网采浮游植物共有6门55属158种(含变种、变型),其中,硅藻41属135种(占85.4%),其丰度占浮游植物总丰度的99.0%;甲藻10属18种(占11.4%),其丰度占总丰度的0.8%;金藻1属2种(占1.3%);蓝藻、绿藻和隐藻各1属1种(各占0.6%)。
根据浮游植物的适盐范围,可将其分为4个生态类群:(1)淡水和半咸水类群,出现频率和丰度均较低,代表种类有桥弯藻属(Cymbella)、颗粒直链藻极狭变种(Melosira granulata var.angustissima)、羽纹藻属(Pinnularia)和单角盘星藻具孔变种(Pediastrum simplex var.duodenarium)等;(2)沿岸河口低盐类群,出现频率和丰度均较高,代表种类有柱状小环藻(Cyclotella stylorum)、克尼角毛藻(Chaetoceros knipowitschi)、尖布纹藻(Gyrosigma acuminatum)和针杆藻属(Synedra)等;(3)近岸内湾性类群,是调查海域的优势类群,代表种类有琼氏圆筛藻(Coscinodiscus jonesianus)、整齐圆筛藻(Coscinodiscus concinnus)、中肋骨条藻(Skeletonema costatum)、布氏双尾藻(Ditylum brightwellii)和微小原甲藻(Prorocentrum minimum)等;(4)外海高盐类群,仅在港口和港口外的个别站位出现,代表种类有镰角藻(Ceratium falcatum)、三角角藻大西洋变种(Ceratium tripos var.atlanticum)和夜光梨甲藻(Pyrocystis noctiluca)等。
2.2 群落参数的平面分布
浮游植物丰度(N)为3.41×104—142.35×104个/m3,平均值(Mean±SD)为(35.29±33.65)×104个/m3,总体呈现港口及港口外低而港底高、支港低而主港高的分布格局,且呈较强的斑块状分布,其高值区位于横山岛至白石山岛水域及乌沙山电厂邻近水域(图2)。物种数(S)为10—50,平均值为28.9±10.3,高值区位于西沪港、港口和南沙岛邻近水域。H′为0.16—3.14,平均值为1.27±0.98,其总体分布表现为全港底部和主港中部<西沪港<港口和港口外。J′为0.09—0.84,平均值为0.82±0.37,其总体分布表现为港底和港中部<港口<港口外(图2)。
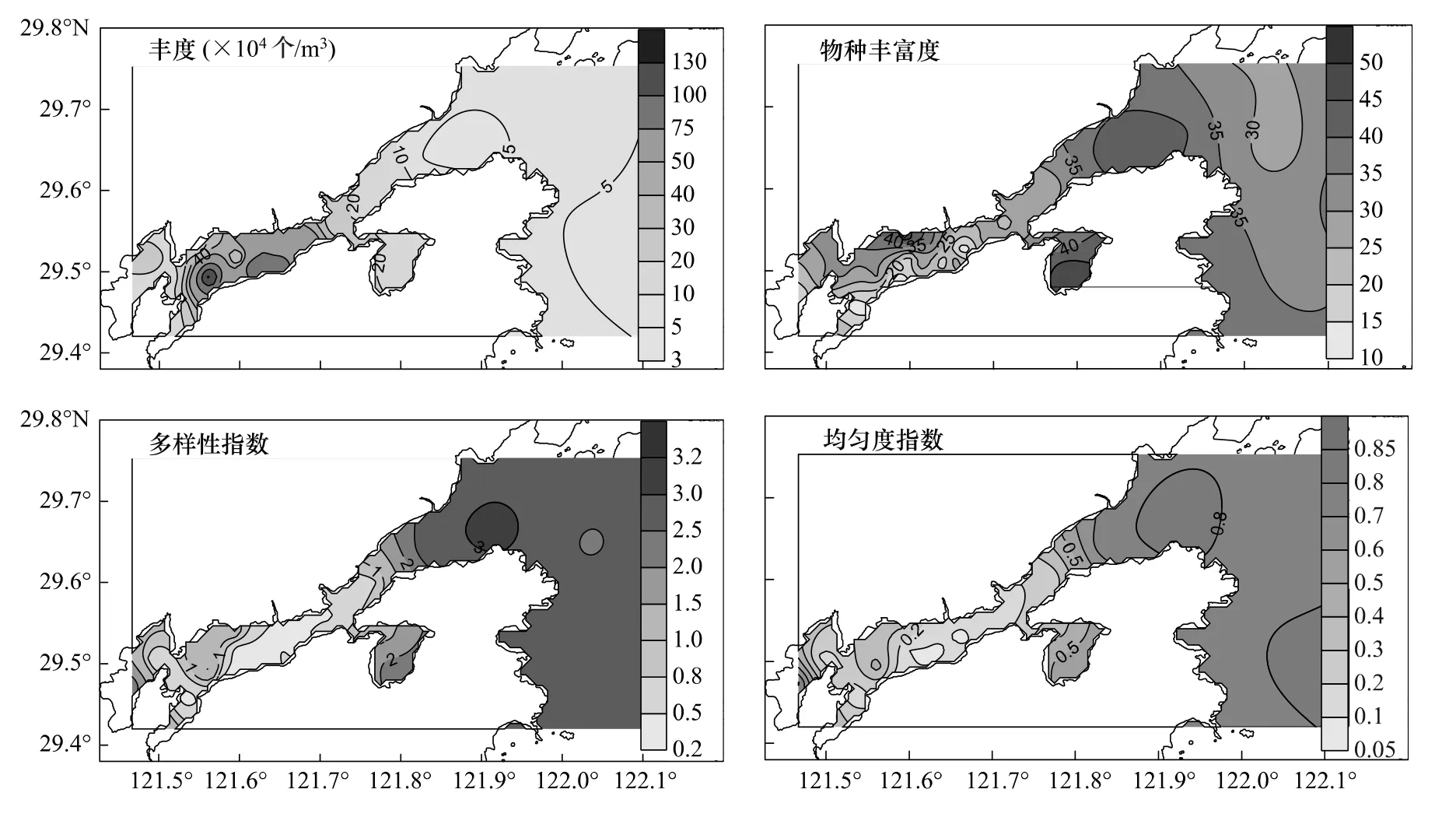
图2 浮游植物丰度、物种丰富度、多样性指数和均匀度指数的平面分布Fig.2 Horizontal distributions of phytoplankton abundance,species richness,diversity and evenness
2.3 优势种组成及其丰度分布
本次调查,共有浮游植物优势种4种,其中,琼氏圆筛藻(Y=0.808)是绝对优势种,丰度为2.41×103—863.49×103个/m3,平均值为(284.97±277.14)×103个/m3,丰度高值区位于横山岛至铜山岛海域及乌沙山电厂邻近海域(图3);盔状舟形藻(Navicula corymbosa,Y=0.025)次之,丰度为0—261.44×103个/m3,平均值为(11.93±49.16)×103个/m3,丰度高值区位于铜山岛和南沙岛海域的海带养殖区(图3);中肋骨条藻(Y=0.016)居第 3 位,丰度为 0—29.95×103个/m3,平均值为(6.92±8.33)×103个/m3,丰度高值区位于铁港港口和西沪港的网箱养殖区(图3);整齐圆筛藻(Y=0.010)居末位,丰度为0—12.19×103个/m3,平均值为(4.06±3.92)×103个/m3,丰度高值区与琼氏圆筛藻类似(图3)。
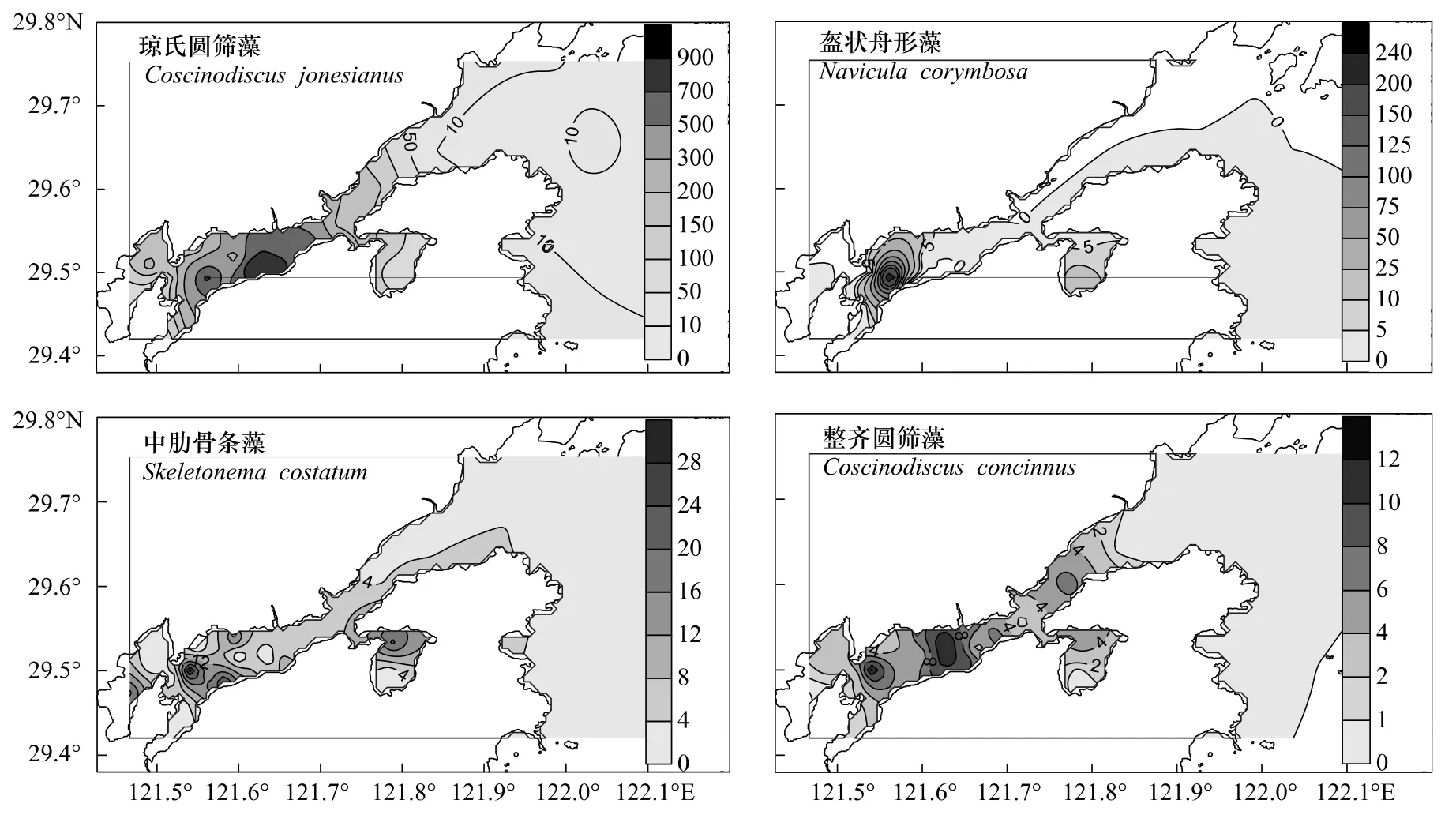
图3 浮游植物优势种丰度(×103个/m3)的平面分布Fig.3 Horizontal distributions of dominant phytoplankton species abundances(×103Cells/m3)
2.4 环境因子分布及其与群落参数的分区比较
主要环境因子与群落参数的分区比较见表1。除水深、DO和无机氮分布无显著差异外,各区其余因子间均存在显著差异(P<0.05)。
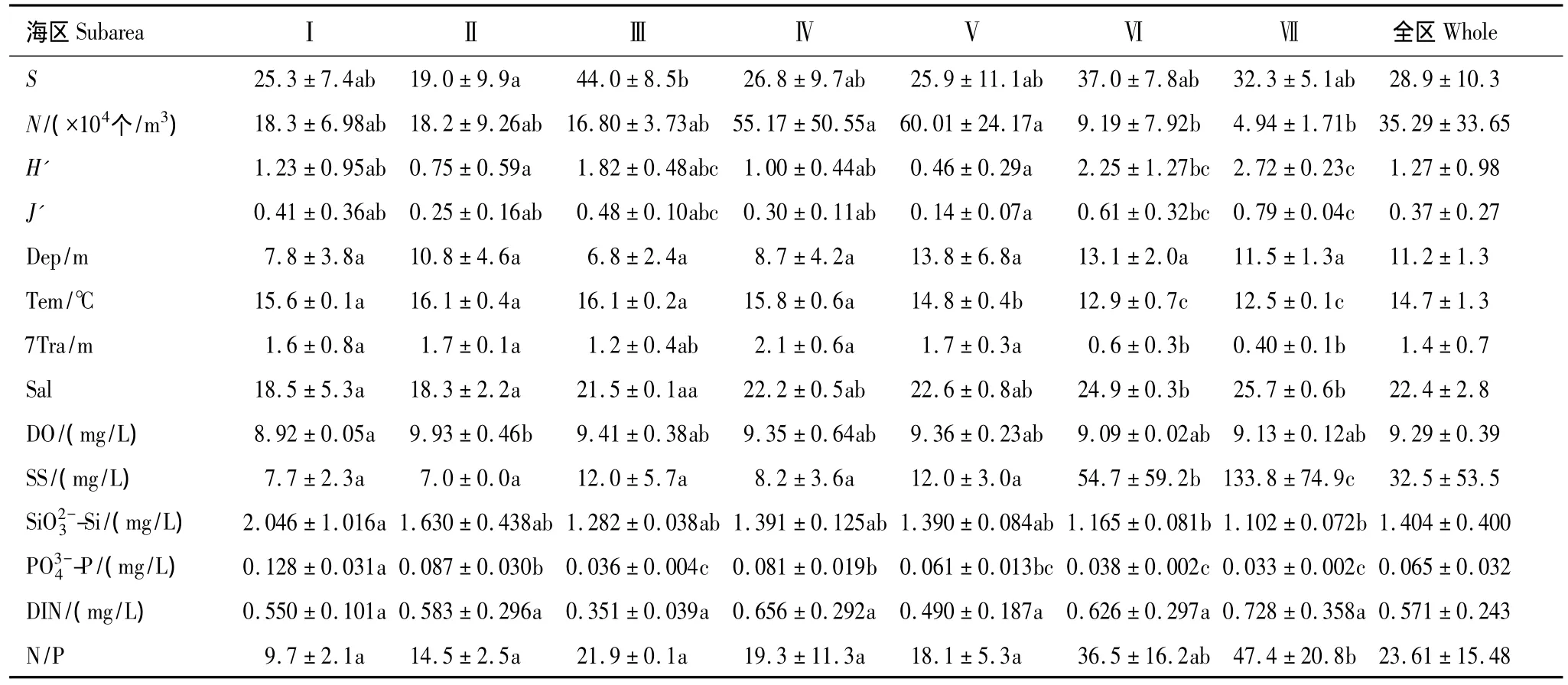
表1 象山港浮游植物群落参数和环境因子的分区比较Table 1 Phytoplankton community indexes and environmental variables(Mean±SD)in different subareas of Xiangshan Bay
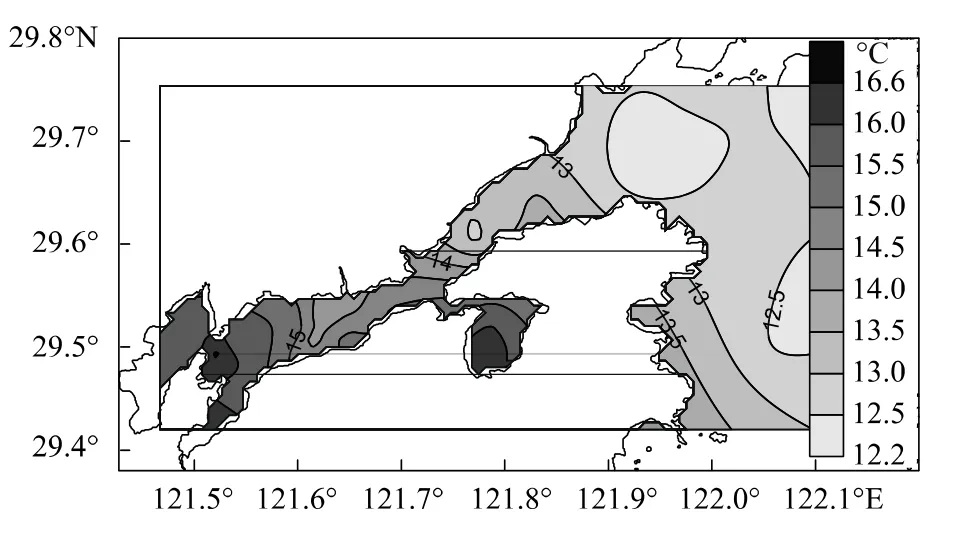
图4 表层水温的平面分布Fig.4 Horizontal distributions of surface water temperature
各站水温为12.2—16.8℃,平均值为(14.7±1.3)℃,分布明显呈港底>港中部>港口>港口外的趋势(图4),特别是宁海电厂排水口邻近海域的水温较港口外正常水温高了4℃多。从图4可知:宁海电厂温排水1℃温升包络线向东已至横山岛周围海域,0.5℃温升包络线向东已至中央山岛水域;乌沙山电厂温排水的1℃温升包络线仅局限于出水口附近的小范围内,0.5℃温升包络线向东可至西沪港口,向西可至白石山岛水域。可见主港底部、中部以及铁港和黄墩港均已纳入两座电厂的0.5℃增温范围内。
盐度为12.5—26.4,平均值为22.4±2.8,其分布则刚好与水温相反,呈港底<港中部<港口<港口外。透明度为0.3—2.5 m,平均值为(1.4±0.7)m,高值区位于横山岛至铜山岛海域及缸爿山岛邻近海域,低值区位于港口和大目洋;悬浮物浓度为2—221 mg/L,平均值为(32±53)mg/L,分布呈港底和港中部<港口<港口外。无机氮浓度为0.323—1.127 mg/L,平均值为(0.571±0.243)mg/L;磷酸盐浓度为0.031—0.157 mg/L,平均值为(0.065±0.032)mg/L;硅酸盐浓度为1.032—3.213 mg/L,平均值为(1.405±0.400)mg/L。磷酸盐和硅酸盐与无机氮的分布存在较大差异,前两者均大致呈港底向港外降低趋势,而后者浓度高值区位于主港中部、港口和港口外。氮磷比(N/P)高值区位于港口和港口外。
2.5 聚类和多维尺度分析
根据聚类(图5)和nMDS(图6)结果,可将浮游植物群落分为 S1站群落、S23—S28站群落、(S5、S6、S9、S13、S21)站群落、(S11、S14—S18、S20、S22)站群落和(S2—S4、S7、S8、S10、S12、S19)站群落,表明港底、港中部、港口至港口外与其余区域的群落组成差异较大。表2中的单因素ANOSIM也表明,不同区域(支港和主港不同区域)间浮游植物群落组成存在极显著差异(Global R=0.47,P=0.001),特别是港口和港口外群落与港内群落差异较大(P<0.05)。
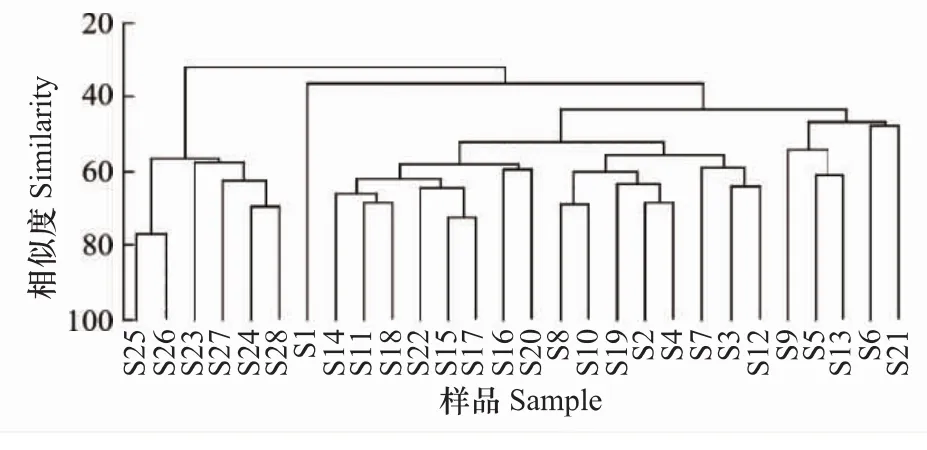
图5 各站位浮游植物样品的聚类分析Fig.5 Clusters analysis of different phytoplankton samples
2.6 典范对应分析
根据浮游植物的出现频率和丰度比例,选取16种用于CCA排序。蒙特卡罗检验表明,第1轴和全部轴均呈极显著差异(P=0.002),故排序结果是可信的。排序图中的11个环境因子可解释浮游植物群落总变量的56.8%。第1轴和第2轴的特征值分别为0.243和0.081,并各自解释了27.8%和9.3%的物种变量。第1轴和第2轴的物种-环境相关系数分别为0.960和0.802,表明这11个环境因子与16种浮游植物主要种类相关性较好。图7表明,透明度、悬浮物、温度、水深和N/P是影响浮游植物群落的主要环境因子。其中,第1轴主要受温度、透明度、悬浮物、无机氮、N/P和磷酸盐等影响,而第2轴则主要由水深、盐度、硅酸盐、DO和pH值等影响。优势种琼氏圆筛藻、整齐圆筛藻和盔状舟形藻与温度、透明度和磷酸盐呈密切正相关,而与悬浮物、N/P和无机氮呈密切负相关;中肋骨条藻与水深和盐度呈密切正相关,而与硅酸盐呈密切负相关。
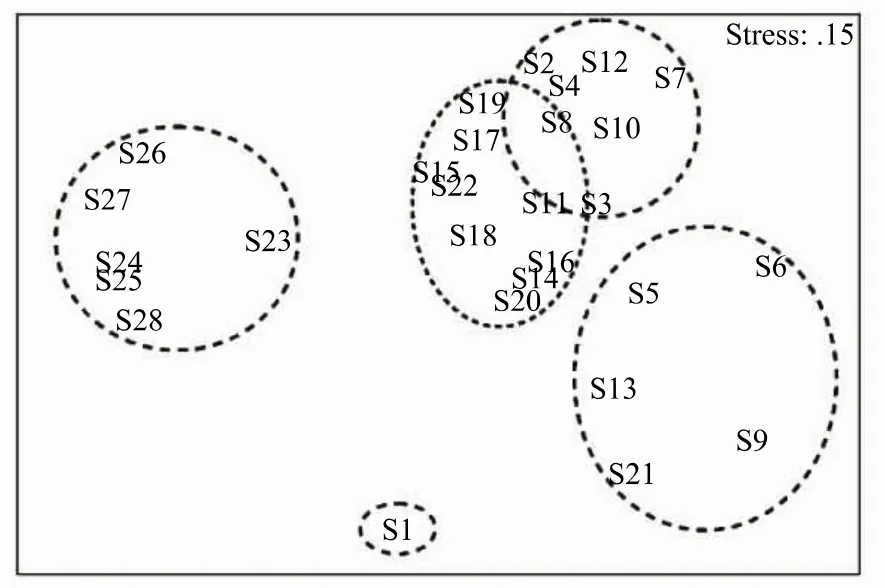
图6 各站位浮游植物样品的多维尺度分析Fig.6 Non-Metric multidimensional scaling(nMDS)of different phytoplankton samples
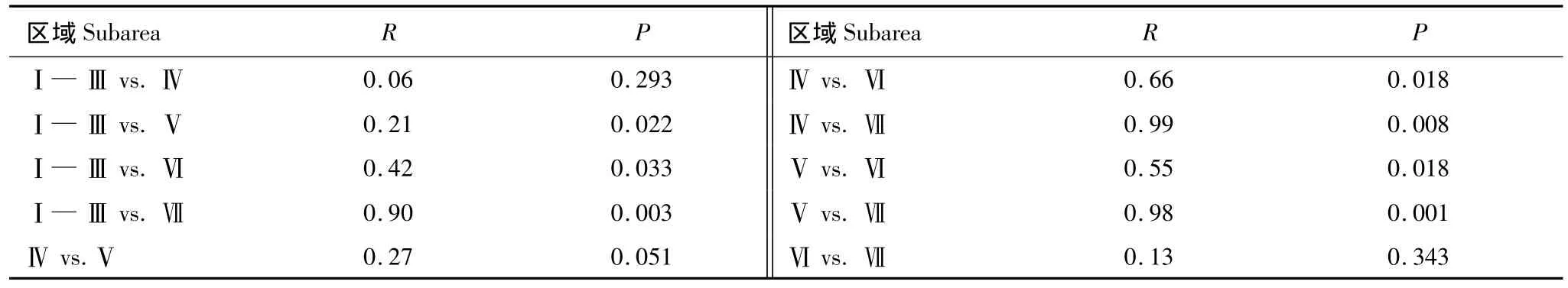
表2 不同区域浮游植物群落的单因素相似性分析Table 2 A one-way analysis of similarity for phytoplankton community at different regions
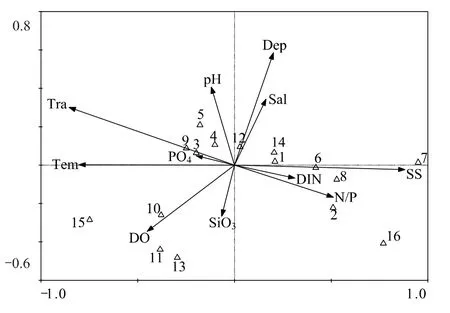
图7 主要浮游植物种类与环境因子间的CCA排序图Fig.7 CCA ordination of main phytoplankton species with environmental variables
3 讨论
3.1 浮游植物群落结构分布特征及其与水团的关系
象山港主要受长江和钱塘江冲淡水为主的沿岸流、台湾暖流和沿港径流影响[18],既有径流入海、冲淡水带来的淡水种类和半咸水种类(桥弯藻、颗粒直链藻极狭变种、羽纹藻和单角盘星藻具孔变种等),又有土著的和沿岸流带来的河口种类(柱状小环藻、克尼角毛藻、尖布纹藻和针杆藻等)与近岸低盐种类(琼氏圆筛藻、整齐圆筛藻、中肋骨条藻、布氏双尾藻和微小原甲藻等)[20-21],此外还有台湾暖流入侵所携带的外海高盐种类(镰角藻、三角角藻大西洋变种和夜光梨甲藻等),故其浮游植物种类组成较复杂,物种丰富度也较高(28.9种)。本次调查共有浮游植物158种,其中:硅藻135种(占总种类数的85.4%,占总丰度的99.0%),甲藻18种(占总种类数的11.4%,占总丰度的0.8%),其余门类5种(占总种类数的3.2%,占总丰度的0.2%)。可见,象山港春季浮游植物群落是以硅藻主导的群落。这与以往的调查结果相符[18-22,25]。琼氏圆筛藻为调查海域的绝对优势种,其细胞数占总丰度的80.8%,且在各个区域均有分布,其余优势种(盔状舟形藻、中肋骨条藻和整齐圆筛藻)仅占6.5%。由于象山港浮游植物优势种组成较单一,导致多样性指数和均匀度指数(图2)均较低,分别为1.27和0.82。这说明春季象山港浮游植物群落组成结构较简单。
3.2 浮游植物群落与环境因子之间的关系
根据CCA结果,透明度、悬浮物、温度、N/P和硅酸盐是影响象山港春季网采浮游植物的主要环境因子(图7)。影响透明度与悬浮物的主要因素就是水动力状况,由于象山港为狭长型海湾,且调查时间刚好处于小潮期间,港内潮流较缓,无法搅起底泥,使得象山港水体表现为港内较清(悬浮物浓度低,透明度高),而港口和港口外水体流速较大,水体较浑浊(悬浮物浓度高,透明度低)。浮游植物的生长与光照密切相关[26],因此,在透明度较高的(普遍>1 m)条件下,光照充足是港内浮游植物旺发的主要原因之一。另外港内温度较高也是促进浮游植物生长的主要因素。由图2和图4可知,浮游植物丰度高值区(横山岛至白石山岛水域及乌沙山电厂邻近水域)刚好对应两座电厂温排水引起的>0.5℃增温水域,这与Zeng等[27]对该处海域温排水的数值模拟结果相符。可见,春季电厂温排水的适度温升可促进浮游植物的生长。廖一波等[28]对象山港浮游植物的温升实验也证实了这一点。营养盐也是影响浮游植物群落最基本的因素。营养盐丰富和N/P适合(接近Redfield比值16)促进了浮游植物的生长。本次调查的丰度高值区(横山岛至白石山岛水域及乌沙山电厂邻近水域)营养盐浓度较高,而N/P又接近16(表1),故其浮游植物丰度也较高。事实上,象山港的浮游植物丰度高值区是上述几个环境因子共同作用的结果,因为该海域整体呈现高透明度、低悬浮物、高营养盐、适合的N/P和适度的温升。
空间异质性是近年来生态学研究的一个极为重要的理论问题[29],同时也是水域生态学家研究不同尺度的生态系统功能和过程中最感兴趣的问题之一[30]。空间异质性是指生态学变量在空间上的不均匀性和复杂性,一般可理解为空间的缀块性和环境梯度的总和[31]。浮游植物的空间异质性由水柱理化特性(如局部水动力、营养盐、温度、盐度及光照透明度等)和生态学过程造成的[30,32-33],现已被认为是水生生态系统存在的一个普遍特征[30],它对海洋生态系统稳定性、多样性、动力学、生产力和水质状况等有极重要的影响。本文尝试将象山港分为支港(包括铁港、黄墩港和西沪港)、主港(港底、港中部和港口)和港口外等7个区块,除水深、DO和无机氮分布无显著差异外,不同区块其余环境因子和浮游植物群落参数均存在显著差异(表1)。且根据聚类分析(图5)、nMDS(图6)和ANOSIM(表2)结果,港底部、港中部、港口至港口外的群落组成差异较大。这说明受径流、海水交换和海洋开发活动(如水产养殖和电厂温排水排放)等影响,象山港的理化参数(透明度、温度、盐度、营养盐浓度和结构等)表现了明显的斑块和梯度分布(表1),这为浮游植物的生长提供了不同的生境,客观造成了其分布的空间异质性(图2和图3)。
3.3 海洋开发活动对浮游植物的影响
2010年象山港春季浮游植物总种类数高于以往,且甲藻种类数也较以往有所增加(表3),这可能与本次调查较以往更密集、范围更广有关。本次调查的网采浮游植物丰度为35.29×104个/m3,该值仅低于1987年的167.80×104个/m3,而高于其余年份的调查数据,且自1988年以来象山港浮游植物数量呈增加趋势(表3),这与营养盐浓度的持续升高有关[1,3,7,18,25]。该海域春季无机氮和磷酸盐浓度自 20 世纪 80 年代[18](分别为0.3066 mg/L和0.0195 mg/L)以来持续升高,到本世纪初[1]时分别达0.388 mg/L和0.021 mg/L,而到本次调查时,已分别增至0.571 mg/L和0.065 mg/L。据Nobre等[34]估算,象山港每年约有4 015 t无机氮和730 t磷来自陆源,同时网箱养殖每年约排放2 354 t无机氮和1 039 t磷,另外虾塘养殖每年还产生42 t无机氮和14t磷。而与此同时,曾相明等[2]研究表明,从1963年到2010年,滩涂围垦已造成象山港全潮平均纳潮量减少12.6%,30 d、60 d和90 d的平均水交换率分别减少7.7%、6.8%和6.1%。因此,象山港大面积的滩涂围垦使原有的水动力环境(潮汐、流场和余流场等)发生改变,纳潮量和水交换率降低,环境容量变小,影响了污染物的自净和向外扩散能力,造成港内工农业污水和养殖废水排放的大量营养盐长期滞留港内,污染(如富营养化和热污染等)加剧。

表3 不同年份象山港春季网采浮游植物的群落参数Table 3 Community parameters of spring net-phytoplankton at different years in Xiangshan Bay
此外,电厂温排水的海水温升也有一定关系。因为在海水适度温升的条件下,富营养化程度加剧[5],温排水的适度温升会促进浮游植物的生长[22,28],从而提高了象山港主港的浮游植物丰度,最终导致本次调查浮游植物丰度的空间分布呈现港口及港口外低而港底高、支港低而主港高(图2和表1)的趋势。且这种空间分布格局与以往调查结果(港口低而港底高、支港高而主港低)并不一致[1,18,20-21,25]。
本次调查琼氏圆筛藻为绝对优势种,且高值区位于宁海电厂至乌沙山电厂的邻近海域(图2)。由于该藻属暖水性种,细胞分裂与温度和光照密切相关[35],在温排水的适度温升和水体透明度较高的条件下,生长繁殖较快。盔状舟形藻的丰度高值区刚好对应于铜山岛和南沙岛的海带养殖区(图2),这是因为该藻属附生性藻类,大量附着于海带叶状体,导致该藻在这两个养殖区及其邻近海域有较高的丰度[22]。同样,中肋骨条藻的丰度高值区对应于铁港港口和西沪港的网箱养殖区(图2)。Morozova和Orlova[36]对日本海Vostok湾的研究也表明,该藻的种群密度与海水养殖面积呈正相关关系。对比历史数据(表3)可知,2010年春季优势种组成与近几年(2007—2009年)的调查结果类似,但与2006年之前的调查结果相差较大。这种改变可能与电厂温排水引起的海水温升有关。海水温升使耐热能力较强的浮游植物种类(如琼氏圆筛藻)成为优势种,而对温升较敏感的种群逐渐丧失优势地位甚至消亡。如自2005年底宁海电厂开始运行后,象山港浮游植物优势种组成就发生了明显改变(表3),同样在我国大亚湾,核电厂温排水造成的海水温升与当地优势种的演替及藻华的发生也有密切联系[37]。此外,象山港浮游植物群落结构的改变与营养盐浓度的升高以及营养盐结构的改变也密切相关[1,3,7,18,25]。如 Zhou 等[38]研究认为,近 50 年来营养盐浓度升高和结构(N/P 和 Si/N)失衡是长江口浮游植物丰度升高、优势种组成改变及赤潮频发的主要原因。因此,象山港浮游植物群落结构的改变与当地的各种海洋开发活动有关。
致谢:感谢刘小涯、姚龙奎、姚梅、莫俊超、刘俊峰和金思韵等协助采样并提供相关理化参数;感谢冉莉华副研究员对本文写作的帮助。
[1] Ning X R,Hu X G.Investigation and Assessment of Aquaculture Capacity of Cage Fish and Cultural Ecology in Xiangshan Bay.Beijing:Ocean Press,2002:1-132.
[2] Zeng X M,Guan W B,Pan C.Cumulative influence of long term reclamation on hydrodynamics in the Xiangshangang Bay.Journal of Marine Sciences,2011,29(1):73-83.
[3] Lü H Q,Chang K M,Shi G D.Circulation and distribution of nitrogen and phosphorus in Xiangshan Bay,China.Oceanologia et Limnologia Sinica,2009,40(2):138-144.
[4] Zhang L X,Jiang X S,Cai Y H,Li Z E.Comprehensive assessment on the situation of water quality at the red tide monitoring area of Xiangshan Harbor in recent four years.Transactions of Oceanology Limnology,2007,(4):98-103.
[5] Yang H,Li C X,Yin C S,Chu M.Contrast of simulation nutrients transport and transformation with pelagic ecosystems of mesocosm in different temperature sections in Xiangshan Bay.Journal of Fisheries of China,2011,35(7):1030-1036.
[6] Liu J J,Zeng J N,Chen Q Z,Jiang Z B,Zhao Y Q,Liao Y B,Shou L,Xu X Q,Huang Y J,Du P.The bacterial ecological distribution in water and sediment of cage culture area in Xiangshangang Bay.Acta Ecologica Sinica,2010,30(2):377-388.
[7] Wang H L,Yang J F,Tu H X,Chen J G,Feng H Q.Spatiotemporal distribution of marine viruses with related to environmental factors in the Xiangshan Bay.China Environmental Science,2011,31(5):834-844.
[8] Wang C S,Liu Z C,He D H.Seasonal dynamics of zooplankton biomass and abundance in Xiangshan Bay.Journal of Fisheries of China,2003,27(6):595-599.
[9] Wang X B,Qiu W S,Qin M L,Wei Y J.Studies on ecological community distribution of zooplankton in Xiangshan Bay.Marine Environmental Science,2009,29(增刊1):62-64.
[10] Du P,Liu J J,Xu X Q,Chen Q Z,Zeng J N,Jiang Z B,Wang Q.Characteristics comparison of zooplankton in different habitats of Xiangshan Bay in winter,2010.Fisheries Science and Technology Information,2011,(2):92-99.
[11] Wu R,Lan D Z,Gao Y H,Chen C P,Lan B B.Preliminary study on the distribution of benthic diatoms and their environments in intertidal zone of Xiangshan Bay.Journal of Oceanography in Taiwan Strait,2008,27(4):445-451.
[12] Gao A G,Chen Q Z,Hu X G,Yang J Y,Dong Y T,Zeng J N,Ning X R,Zhang J.Ecological characteristics on macrobenthos of netcage cultural areas in the Xiangshan Bay.Acta Oceanologica Sinica,2005,27(4):108-113.
[13] Gu X Y,Tao L,Shi H X,Lou D,Jiao H F,You Z J.Macrobenthic faunal diversity in Xiangshan Bay.Chinese Journal of Applied Ecology,2010,21(6):1551-1557.
[14] Liao Y B,Shou L,Zeng J N,Gao A G,Jiang Z B.A comparative study of macrobenthic community under different mariculture types in Xiangshan Bay,China.Acta Ecologica Sinica,2011,31(3):646-653.
[15] Gao Y H,Yu Q B,Qi Y Z,Zhou J Z,Lu D D,Li Y,Chen C P.Species composition and ecological distribution of planktonic diatoms in the Changjiang River estuary during spring.Chinese Journal of Applied Ecology,2003,14(7):1044-1048.
[16] Luan Q S,Sun J,Song S Q,Shen Z L,Yu Z M.Canonical correspondence analysis of summer phytoplankton community and its environment in the Yangtze River Estuary,China.Journal of Plant Ecology,2007,31(3):445-450.
[17] Li Y,Li H,Lü S H,Jiang T J,Xiao Y P,Chen S.Species diversity and distribution of phytoplankton in Nanji Islands national nature reserve.Acta Hydrobiologica Sinica,2010,34(3):618-628.
[18] Editorial Committee of the Bay Chorography in China.The Bay Chorography in China:part 5.Beijing:Ocean Press,1992:166-233.
[19] Zhang H B,Cai Y H,Xiang Y T.Study on the phytoplankton and HAB in Xiangshan Bay.Marine Science Bulletin,2005,24(1):92-96.
[20] Gu X G,Xu Z L.Central and western parts of Xiangshangang,Zhejiang.Marine Fisheries,1993,15(2):199-209,215-215.
[21] You Z J,Jiao H F.Study on the Ecology and Environmental Protect and Restored Technology of Xiangshan Bay.Beijing:Ocean Press,2011:125-137.
[22] Jiang Z B,Chen Q Z,Zeng J N,Liao Y B,Shou L,Liu J J.Phytoplankton community distribution in relation to environmental parameters in three aquaculture systems in a Chinese subtropical eutrophic bay.Marine Ecology Progress Series,2012,446:73-89.
[23] General Administration of Quality Supervision,Inspection and Quarantine.Marine Investigation Criterion(GB/T12763.6-2007).Beijing:China Standard Press,2007.
[24] Lepš J,Šmilauer P.Multivariate Analysis of Ecological Data Using CANOCO.London:Cambridge University Press,2003:1-280.
[25] Zeng J N,Pan J M,Liang C J,Chen Q Z.Integrated Investigation Report of the Ecology and Environment of Key Bay in Zhejiang Province.Beijing:Ocean Press,2011:189-194.
[26] Miao Z Q,Yang D F.Solar light,seawater temperature,and nutrients,which one is more important in affecting phytoplankton growth?Chinese Journal of Oceanology and Limnology,2009,27(4):825-831.
[27] Zeng G N,Guang W B,Zeng J N,Chen Q Z,Ai N.3D modeling of the thermal effluents dispersion from power plants in the Xiangshan Bay.Energy Education Science and Technology Part A:Energy Science Research,2011,28(1):71-82.
[28] Liao Y B,Chen Q Z,Zeng J N,Gao A G,Liu J J,Jiang Z B.Thermal effect on the marine phytoplankton in Xiangshan Bay,China.Acta Ecologica Sinica,2008,28(9):4203-4212.
[29] Kolasa J,Pickett S T A.Ecological Heterogeneity.New York:Springer Verlag,1991:1-332.
[30] Pinel-Alloul B,Ghadouani A.Spatial heterogeneity of planktonic microorganisms in aquatic systems//Franklin R B,Mills A L,eds.The Spatial Distribution of Microbes in the Environment.Dordrecht:Springer,2007:203-310.
[31] Wu J G.Landscape Ecology.Beijing:Higher Education Press,2000:1-258.
[32] Martin A P.Phytoplankton patchiness:the role of lateral stirring and mixing.Progress in Oceanography,2003,57(2):125-174.
[33] Seuront L.Hydrodynamic and tidal controls of small scale phytoplankton patchiness.Marine Ecology Progress Series,2005,302:93-101.
[34] Nobre A M,Ferreira J G,Nunes J P,Yan X J,Bricker S,Corner R,Groom S,Gu H F,Hawkins A J S,Hutson R,Lan D Z,Lencart E S J D,Pascoe P,Telfer T C,Zhu M Y,Zhang X L.Assessment of coastal management options by means of multilayered ecosystem models.Estuarine,Coastal and Shelf Science,2010,87(1):43-62.
[35] Huang S Y,Lin J M.Effect of light and temperature on the cell division in Coscinodiscus jonesianus.Journal of Jimei University,1998,3(3):126-131.
[36] Morozova T V,Orlova T Y.Monitoring of phytoplankton in the area of a sea farm in Vostok Bay(Sea of Japan).Russian Journal of Marine Biology,2005,31(1):1-6.
[37] Li T,Liu S,Huang L M,Huang H,Lian J S,Yan Y,Lin S J.Diatom to dinoflagellate shift in the summer phytoplankton community in a bay impacted by nuclear power plant thermal effluent.Marine Ecology Progress Series,2011,424:75-85.
[38] Zhou M J,Shen Z L,Yu R C.Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang(Yangtze)River.Continental Shelf Research,2008,28(12):1483-1489.
参考文献:
[1] 宁修仁,胡锡钢.象山港养殖生态和网箱养鱼的养殖容量研究与评价.北京:海洋出版社,2002:1-132.
[2] 曾相明,管卫兵,潘冲.象山港多年围填海工程对水动力影响的累积效应.海洋学研究,2011,29(1):73-83.
[3] 吕华庆,常抗美,石钢德.象山港氮、磷营养盐环流和分布规律的研究.海洋与湖沼,2009,40(2):138-144.
[4] 张丽旭,蒋晓山,蔡燕红,李志恩.近4年来象山港赤潮监控区水质状况综合评价.海洋湖沼通报,2007,(4):98-103.
[5] 杨红,李春新,印春生,储鸣.象山港不同温度区围隔浮游生态系统营养盐迁移—转化的模拟对比.水产学报,2011,35(7):1030-1036.
[6] 刘晶晶,曾江宁,陈全震,江志兵,赵永强,廖一波,寿鹿,徐晓群,黄逸君,杜萍.象山港网箱养殖区水体和沉积物的细菌生态分布.生态学报,2010,30(2):377-388.
[7] 王海丽,杨季芳,屠霄霞,陈吉刚,冯辉强.象山港海洋病毒时空分布特征及其环境影响因素.中国环境科学,2011,31(5):834-844.
[8] 王春生,刘镇盛,何德华.象山港浮游动物生物量和丰度的季节变动.水产学报,2003,27(6):595-599.
[9] 王晓波,邱武生,秦铭俐,魏永杰.象山港浮游动物生态群落分布的研究.海洋环境科学,2009,29(增刊1):62-64.
[10] 杜萍,刘晶晶,徐晓群,陈全震,曾江宁,江志兵,王琪.象山港不同生境冬季浮游动物生态特征比较研究.水产科技情报,2011,(2):92-99.
[11] 吴瑞,蓝东兆,高亚辉,陈长平,兰彬斌.象山港潮间带底栖硅藻的分布及其与环境关系探讨.台湾海峡,2008,27(4):445-451.
[12] 高爱根,陈全震,胡锡钢,杨俊毅,董永庭,曾江宁,宁修仁,张健.象山港网箱养鱼区大型底栖生物生态特征.海洋学报,2005,27(4):108-113.
[13] 顾晓英,陶磊,施慧雄,楼丹,焦海峰,尤仲杰.象山港大型底栖动物生物多样性现状.应用生态学报,2010,21(6):1551-1557.
[14] 廖一波,寿鹿,曾江宁,高爱根,江志兵.象山港不同养殖类型海域大型底栖动物群落比较研究.生态学报,2011,31(3):646-653.
[15] 高亚辉,虞秋波,齐雨藻,邹景忠,陆斗定,李扬,陈长平.长江口附近海域春季浮游硅藻的种类组成和生态分布.应用生态学报,2003,14(7):1044-1048.
[16] 栾青杉,孙军,宋书群,沈志良,俞志明.长江口夏季浮游植物群落与环境因子的典范对应分析.植物生态学报,2007,31(3):445-450.
[17] 李扬,李欢,吕颂辉,江天久,萧云朴,陈舜.南麂列岛海洋自然保护区浮游植物的种类多样性及其生态分布.水生生物学报,2010,34(3):618-628.
[18] 中国海湾志编纂委员会.中国海湾志第五部分.北京:海洋出版社,1992:166-233.
[19] 张海波,蔡燕红,项有堂.象山港水域浮游植物与赤潮生物种群动态研究.海洋通报,2005,24(1):92-96.
[20] 顾新根,徐兆礼.象山港中、西部浮游植物生态及赤潮研究.海洋渔业,1993,15(2):199-209,215.
[21] 尤仲杰,焦海峰.象山港生态环境保护与修复技术研究.北京:中国海洋出版社,2011:125-137.
[23] 国家质量监督检验检疫局.海洋调查规范(GB/T12763.6-2007).北京:中国标准出版社,2007.
[25] 曾江宁,潘建明,梁楚进,陈全震.浙江省重点港湾生态环境综合调查报告.北京:海洋出版社,2011:189-194.
[28] 廖一波,陈全震,曾江宁,高爱根,刘晶晶,江志兵.海洋浮游植物的热效应.生态学报,2008,28(9):4203-4212.
[31] 邬建国.景观生态学.北京:高度教育出版社,2000:1-258.
[35] 黄世玉,林均民.琼氏圆筛藻细胞分裂的光温效应.集美大学学报,1998,3(3):126-131.
