清江流域生态系统植被不同林分因子随海拔梯度的变化
2013-12-07徐静静艾训儒
徐静静,艾训儒,2*,蒲 鑫,朱 江
(1.湖北民族学院 林学园艺学院,湖北 恩施 445000; 2.生物资源保护与利用湖北省重点实验室(湖北民族学院),湖北 恩施 445000)
清江流域生态系统植被不同林分因子随海拔梯度的变化
徐静静1,艾训儒1,2*,蒲 鑫1,朱 江1
(1.湖北民族学院 林学园艺学院,湖北 恩施 445000; 2.生物资源保护与利用湖北省重点实验室(湖北民族学院),湖北 恩施 445000)
选择清江流域的南部和北部进行垂直样带调查,将2个调查地段分别等距离划分为高 、中和低3个海拔区域,研究林分因子在不同海拔范围内的差异,并比较2个地段林分因子随海拔梯度变化的总体趋势.结果表明:2个地段的林分因子随海拔梯度的变化呈现不同的变化格局.在南部,林分因子(平均胸径、最大胸径、平均树高、最大树高、胸高断面积和蓄积量)随海拔的增加逐渐下降;在北部,林分因子(平均胸径、最大胸径、平均树高、最大树高、胸高断面积和蓄积量)随海拔梯度的变化呈单峰型变化趋势.尽管相同的林分因子在两个地段呈现不同的变化格局,但两个地段林分因子的变化都可以用二次曲线方程y=ax2+bx+c(a、b、c为常数)进行描述.
清江流域;林分因子;海拔梯度
植被沿海拔梯度的变化规律是当前生态学研究的热点问题之一.研究植被林分因子随海拔梯度变化的格局对揭示森林群落特征时空变化的驱动因素,从而合理调控植被群落分布具有重要意义.有研究结果表明,物种丰富度随海拔梯度的变化一般有5种变化规律,分别为随着海拔的增加呈单峰格局、单调下降、先下降后上升、单调上升和没有明显变化的格局[1-5].一般认为,前两种的林分因子随海拔梯度的变化格局是比较普遍的[6-9].研究表明,环境因子随海拔梯度的差异也发生相应的变化[10-13],林分因子的变化趋势受环境因素的影响[5,14-15];不同方式的干扰也可直接导致生态系统以及植被林分因子的变化[8,16-18].因此林分因子随海拔梯度的变化格局可能来源于环境因子和干扰等因素的影响.
清江流域地处湖北省西南部,位于东经108°35′~111°35′,北纬29°33′~30°50′,是长江在湖北省境内的第二大支流,发源于湖北省利川市齐岳山,流经利川、咸丰、恩施、宣恩、建始、巴东、鹤峰、五峰、长阳、宜都等10个县市,干流全长约423 km,流域面积达17000平方公里.清江流域的森林资源十分丰富,是省内第二大用材基地,育林面积达到93.3万hm2.目前对清江流域的研究多集中在清江流域的地质、文化或植被分子水平等方面[19-21],对清江流域生态系统植物多样性垂直梯度格局的研究报道较少.本研究选择在清江流域的南部和北部进行植物多样性的垂直样地调查,分析清江流域不同地段的生态系统植被林分因子在不同海拔范围内的变化规律,为了解清江流域植物多样性随海拔梯度变化的规律以及对生态系统群落的经营管理提供科学依据.

表1 研究区概况
1 研究区概况
选择位于我国湖北境内清江流域的南部和北部两个地段垂直群落学调查,在清江源的南部还包含有国家一级自然保护区星斗山自然保护区,研究地段的基本概况见表1.
2 研究方法
2.1样地设置
采用样方法进行群落调查,在清江流域的南部和北部(海拔815~1 753 m)两个地段进行布设样方,根据群落结构的相对完整性在每个地段的不同海拔高度随机选择样地及样方的个数,利用全站仪设置面积为20 m×20 m的样方,共计调查的样方为39个.
2.2调查与分析内容
将清江源的南部和北部的海拔分别尽量等间距划分为高、中和低3个海拔范围,分析清江流域植被群落的主要林分因子(平均胸径、最大胸径、平均树高、最大树高、林分密度、胸高断面积和蓄积量)随海拔梯度变化的不同规律.采用不同的函数对主要林分因子随海拔梯度变化的关系进行回归分析,所有的数据处理在Excel和Origin7.5软件中进行.
样方调查时,将每个样地划分为4块10 m×10 m的小样地,并对其中的乔木、灌木和草本进行调查,分别记录乔木活立木的种类、丛树、树高以及胸径;记录灌木的种名、丛树、树高以及基径;草本层调查记录植物的种类、丛数,并估测其盖度等指标.在上述样方指标调查完成后通过手持GPS测量样方的经纬度、海拔、坡向等地形因子,并记录群落的人为干扰、群落成熟状况等.调查样地的工作在2012年6~9月进行并完成.
2.3 蓄积量的计算方法
根据调查林分因子(树高、胸径)的数据计算每个样方内乔木和灌木胸径或基径>6 cm植被的材积;林分蓄积量是按照湖北省一元立木材积表[22]的经验公式计算每块样地内不同树种的材积.
鄂西南不同树种材积的计算公式:
D轮=A+B×D围
V=aD轮bHc
式中:D轮为植被在树高1.3 m处的轮径,D围为植被调查时乔木或灌木在树高1.3 m处的胸径或基径,V和H分别为每株乔木或灌木的材积和树高,A、B、a、b、c为常数,不同树种的A、B、a、b、c参考湖北省一元立木材积表取值不同.
3 结果与分析
3.1 林分概况
调查发现,在清江源南部植被群落在低海拔处以针叶林为主,中海拔处为常绿阔叶混交林,在高海拔处则为常绿阔叶林,植物群落的发育状况主要为成熟型群落,恢复型群落占有少数.在清江源北部植被群落随海拔梯度的增加依次为针叶林、落叶阔叶林和针阔混交林,植物群落的发育状况以恢复型群落为主.调查还发现,清江源南部不同海拔处的植物群落类型有所差异,海拔810~1 110 m处主要混生有杉木、秃杉(Taiwaniacryptomerioides)、毛竹、钩栲(CastanopsistibetanaHance)、丝栗栲(Castanopsisfargesii)、枫香(LiquidambarformosanaHance)等;海拔1 110~1 420 m处主要混生有有珙桐(DavidiainvolucrataBaill)、楠木(PhoebezhennanS. Lee et F. N. Wei)等;海拔1 420~1 760 m处主要混生有鹅耳枥(CarpinusturczaninowiiHance)、青冈(Cyclobalanopsisglauca(Thunberg) Oersted)、水青冈(FaguslongipetiolataSeem)等.在北部不同海拔处均混生有马尾松(PinusmassonianaLamb)和杉木(Cunninghamialanceolata),此外海拔1 130~1 240 m处还混生有的植被有毛竹(ph.edulis(carr)h.de)、猫儿屎(Decaisneainsignis);海拔1 240~1 350 m处混生有柳杉(CryptomeriafortuneiHooibrenk ex Otto et Dietr)、锥栗(Castaneahenryi(Skam) Rehd. et Wils.);海拔1 240~1 350 m处混生有锥栗、香椿(Toonasinensis)、盐肤木(Rhuschinensis)等.
3.2 林分因子随海拔梯度的变化规律
不同海拔高度的林分因子如表2所示.

表2 不同海拔高度的林分因子

图1 平均树高随海拔梯度的变化Fig.1 Mean tree height along an altitudinal gradient changes

图2 最大树高随海拔梯度的变化Fig.2 The maximum height along an altitudinal gradient changes
从表2中看出不同的林分因子的变化规律随着海拔高度的变化存在差异.在清江源南部,林分平均胸径在低海拔处最大,高海拔处最小,在清江源北部则是中海拔处最大,低海拔出最小.林分最大胸径在南部和北部的低、中海拔处相同,但在南部高海拔处林分最大胸径的值较小,在北部的高海拔处林分最大胸径的值较大.林分平均树高在2个地段呈现出不同的变化趋势:在南部,林分平均树高随着海拔梯度的增加逐渐减小;在北部,林分平均树高在中海拔处最大,低海拔处最小.林分最大树高在南部的变化趋势为中海拔>低海拔>高海拔,在北部林分最大树高在中、高海拔处相同且大于低海拔处.林分密度在南部和北部的变化呈相反的趋势,在南部林分密度随着海拔的增加逐渐减小,在北部林分密度随着海拔的增加逐渐增加.2个地段的林分胸高断面积变化趋势也较为复杂,在南部的低、中海拔处较为接近,高海拔处最小,在北部,林分胸高断面积与林分平均胸径的变化趋势表现出一致性,即为中海拔>高海拔>低海拔.在南部,林分蓄积量与林分胸高断面积的变化趋势相似,均是在低、中海拔处变化不大,高海拔处最小,在北部,林分蓄积量在中海拔处取得最大值,高海拔处次之,低海拔处最小.
3.3林分因子随海拔梯度的变化
3.3.1 平均树高随海拔梯度的变化 从图1中可以看出,清江源南部和北部林分平均树高随海拔梯度的变化呈现出不同的变化格局,在南部林分平均树高随着海拔梯度的增加呈现下降的趋势,在北部林分平均树高随海拔的增加呈现单峰型变化格局,但在清江源的南部P=0.022和北部P=0.024都达到了显著相关水平.
3.3.2 最大树高随海拔梯度的变化 从图2中可以看出,在清江源南部林分最大树高随海拔的变化呈下降的趋势,P=0.014达到了显著相关水平.在清江源北部林分最大树高随海拔梯度的增加呈现单峰型的变化格局,P=0.000达到了极显著相关水平.
3.3.3 平均胸径随海拔梯度的变化 从图3中可以看出,在清江源南部林分平均胸径随海拔梯度的变化呈现下降的趋势,达到了极显著相关水平P=0.004.在清江源北部林分平均胸径随海拔梯度的增加呈现单峰型变化格局,达到了极显著相关水平P=0.002.

图3 平均胸径随海拔梯度的变化Fig.3 The average DBH with elevation gradient

图4 最大胸径随海拔梯度的变化Fig.4 The maximum DBH with elevation gradient
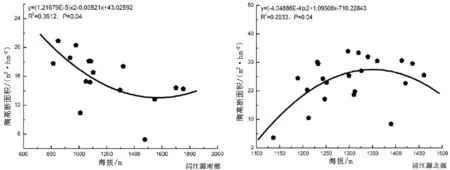
图5 胸高断面积随海拔梯度的变化Fig.5 Total basal area with elevation gradient

图6 林分密度随海拔梯度的变化Fig.6 Stand density with altitude gradient

图7 蓄积量随海拔梯度的变化Fig.7 Volume with an altitudinal gradient
3.3.4 最大胸径随海拔梯度的变化 从图4中可以看出,在清江源南部林分最大胸径随着海拔梯度的变化呈下降的趋势,在清江源北部林分最大胸径随海拔的增加呈现单峰型变化格局,在南部P=0.02则达到了显著相关水平,在北部P=0.000达到了极显著相关水平.
3.3.5 胸高断面积随海拔梯度的变化 从图5中可以看出,在清江源南部林分胸高断面积随着海拔的变化呈现出下降的趋势,在清江源北部林分胸高断面积随海拔梯度的增加呈现单峰型变化格局,并且在南部P=0.04和北部P=0.04和都达到了显著相关水平.
3.3.6 林分密度随海拔梯度的变化 从图6中可以看出,在清江源南部(P=0.838)和北部(P=0.473)林分密度随海拔梯度梯度的变化没有达到显著相关水平,在南部林分密度随着海拔的增加呈现单峰型的变化格局,但在北部林分密度随着海拔梯度的变化呈现上升的趋势.
3.3.7 蓄积量随海拔梯度的变化 从图7中可以看出,在南部林分蓄积量随着海拔梯度的增加呈现下降的趋势,在清江源北部林分蓄积量随海拔的变化呈现单峰型变化格局,在南部P=0.045和北部P=0.04均达到了显著相关水平.
4 小结
数据分析表明,清江源南部和北部生态系统植被不同林分因子(平均胸径、最大胸径、平均树高、最大树高、胸高断面积和蓄积量)随着海拔梯度的变化呈现不同的变化趋势,清江源南部林分因子随着海拔的升高呈下降的趋势,为负相关关系;在清江源北部,林分因子随着海拔的增加呈现单峰型变化格局.在本研究中,清江流域气候湿润,雨量充沛,地形复杂,土壤厚度及养分状况良好,光照强度由东北向西南逐渐递减,降水和风速随着海拔的升高而逐渐加大,气温则随海拔升高呈递减的趋势[22-23].样方调查时,在清江源南部,样地选择为天然林群落,植被在受环境因素的影响较大的背景下,林分因子随海拔梯度的升高而呈下降的趋势;在清江源北部,低海拔区群落的选择包含有成熟型和恢复型群落,受人为干扰和环境因素的影响[18],林分因子随海拔梯度的变化呈单峰型变化格局.因此,清江源林分因子的变化对该区域的环境梯度变化都有较好的指示作用.
研究结果显示,虽然清江源的南部和北部两个地段林分因子随海拔梯度的变化呈现不同的变化格局,但清江源的林分因子平均胸径、最大胸径、平均树高、最大树高、林分密度和蓄积量(y)随海拔高度(x)的变化可用方程y=ax2+bx+c(a、b、c为常数)进行描述.
[1] 朱源,康慕谊,江源,等,贺兰山木本植物群落物种多样性的海拔格局[J].植物生态学报,2008,32(3): 574-581.
[2] 刘兴良,岳永杰,郑绍伟,等.川滇高山栎种群统计特征的海拔梯度变化[J].四川林业科技,2005, 26(4):9-15.
[3] 段敏杰,高清竹,郭亚奇,等.藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局[J].草业科学,2011,28(10):1845-1850.
[4] 刘贵峰, 臧润国,郭仲军,等.不同经度天山云杉群落物种丰富度随海拔梯度变化[J].应用生态学报,2008,19(7):1407-1413.
[5] 刘贵峰, 臧润国,张新平,等.不同经度天山云杉林分因子随海拔梯度的变化[J].林业科学, 2009,45(8):9-13.
[6] 唐志尧,方精云.植物物种多样性的垂直分布格局[J].生物多样性,2004,12(1):20-28.
[7] 冯建孟,王襄平,徐成东,等.玉龙雪山植物物种多样性和群落结构沿海拔梯度的分布格局[J].山地学报,2006,24(1):110-116.
[8] 唐志尧,柯金虎.秦岭牛背梁植物物种多样性垂直分布格局[J].生物多样性,2004,12(1):108-114.
[9] 沈泽昊,刘增力,方精云.贡嘎山海螺沟冷杉群落物种多样性与群落结构随海拔的变化[J].生物多样性,2004,12(2):237-244.
[10] Barry R G.Mountain Weather and Climate[M].England:Cambridge University Press,1981.
[11] Grytnes J A.Species-richness patterns of vascular plants along seven altitudinal transects in Norway[J].Ecography,2003,26:291-300.
[12] Carpenter C.The environmental control of plant species density on a Himalayan elevation gradient[J].Journal of Biogeography,2005,32:999-1018
[13] Bhattarai K R,Vetaas O R.Variation in plant species richness of different life forms along a subtropical elevation gradient in the Himalayas,east Nepal[J].Global Ecology and Biogeography,2003,12:327-340.
[14] 潘红丽,李迈和,蔡小虎,等.海拔梯度上的植物生长与生理生态特性[J].生态环境学报,2009,18(2):722-730.
[15] Bergengren J,Thompson S,Pollard D,et al.Modeling global climate-vegetation interactions in a doubled CO2world[J].Netherlands,2001,50:31-75.
[16] 李新彬,王襄平,罗菊春,等.干扰对长白山北坡次生林群落结构和树种多样性的影响[J].林业科学报, 2006,42(2):105-110.
[17] 王广慧,乌兰,于军.干扰与生态系统的关系[J].内蒙古草业,2007,19(1):15-18.
[18] 艾训儒.人为干扰对森林群落及生物多样性的影响[J].福建林业科技,2006, 33(3):5-9.
[19] 吴茜.清江流域非物质文化遗产的类别、现状及保护[J].湖北民族学院学报:哲学社会科学版,2007,25(5):48-51.
[20] 毕庆文,于建军,汪键,等.清江流域烤烟类胡萝卜素降解产物的因子分析和综合评价[J].中国烟草科学,2009,30(6):25-29.
[21] 艾训儒.湖北省清江流域土家族生态学研究[D].北京:北京林业大学,2006,1-202.
[22] 沈作奎.湖北省一元立木材积表[M].武汉:湖北省林业勘察设计院,1985.
[23] 罗书文,张远海,陈伟海,等.基于流域水文地貌系统的清江流域地貌研究[J].安徽农业科学,2010,38(3):1646-1649
StandFactorsofQingjiangRiverBasinalongtheAltitudinalGradientinDifferentLongitudes
XU Jing-jing1,AI Xun-ru1,2*,PU Xin1,ZHU Jiang1
(1.School of Forestry and Horticulture,Hubei University for Nationalities,Enshi 445000,China;2.Key Laboratory of Biologieal Resources Protection and Utilization of Hubei Province, Hubei University for Nationalities,Enshi 445000,China)
The vertical sample strips of Southern and northern in Qingjiang River Basin were investigated.Each site was divided into three altitudinal ranges (high,middle and low) and the difference in stand factors within each site range was analyzed.The variation of main stand factors along the altitudinal gradient was investigated. The results showed that variation of different stand factors within each site presented different patterns.In the southern,the stand factors (mean and maximum DBH,mean and maximum tree height,total basal area and stand volume) gradually deereases with the increase of altitude;in the northern,the stand factors showed hump- shaped curves with increasing of the altitude.Although the same stand factors in the two areas present different change pattern, the stand factors in the two areas could be described by the quadratic curve equationsy=ax2+bx+c(a、b、c for constant).
Qingjiang river basin; stand factors; altitudinal gradient
2013-03-31.
湖北省教育科学技术研究计划重点项目.
徐静静(1986- ),女,硕士生,主要从事民族植物与生态的研究;*
:艾训儒(1967- ),男,教授,博士,主要从事生态学与民族学研究.
S718.55
A
1008-8423(2013)02-0153-05
