牛c-Myc/TAT/9R融合表达载体构建及其原核表达研究
2013-11-30贾文超潘传英蓝贤勇
甘 鹏, 熊 亮, 贾文超, 潘传英,* , 蓝贤勇, 陈 宏
(1.西北农林科技大学 生命科学学院,陕西 杨凌 712100; 2. 中山大学 生命科学学院,广东 广州 510275;3. 西北农林科技大学 动物科技学院,陕西 杨凌712100)
原癌基因最早发现于伯基特淋巴瘤(Burkitt's lymphoma)患者的肿瘤细胞中,因其序列类似于骨髓细胞瘤病毒v-Myc,故被命名为c-Myc[1]。目前,许多研究表明:c-Myc蛋白在乳腺癌、胃癌及白血病等许多肿瘤中都存在异常高表达[2];c-Myc不仅是可易位基因,而且还是多种物质调节的可调节基因;此外,c-Myc也是一种可使细胞无限增殖且获得永生化功能、促进细胞分裂的基因[3-5]。2006 年,日本科学家利用c-Myc和Oct4等4个转录因子诱导小鼠胚胎成纤维细胞在国际上首次获得了诱导多潜能干细胞(iPS)[5]。深入研究发现,c-Myc等可通过MAPK 和 GSK3β 途径协同降低细胞死亡,维持干细胞全能性。由于体细胞重编程技术为实现自体细胞治疗和研究细胞多能性机理具有重要科学价值[3-4],为此,c-Myc基因及其表达在iPS研究中具有重要作用。目前,关于c-Myc等重要转录因子诱导哺乳细胞重编程研究中,介导外源基因进入靶细胞的方式主要涉及3个水平:① DNA水平,即病毒介导DNA分子[5];② RNA水平,即mRNA和miRNA分子重编程[6-7];③ 蛋白水平:转录因子蛋白介导[8]。其中,前两种方式具有低效性、难操作性、高毒性和复杂性[5-8],因此,利用转录因子蛋白直接重编程是获得iPS细胞的最佳选择。蛋白转导区域 (Protein transduction domain,PTD) 是一类能够帮助外源蛋白直接穿透细胞膜而在细胞内聚集的小阳离子肽家族[9],如来源于HIV病毒的TAT[10]、11聚精氨酸(poly(11R)[11]和9聚精氨酸(poly(9R))[12-14]等。作为特殊的小分子,它们能够突破细胞膜脂质双分子层的疏水特性从而穿透细胞膜进入细胞内[10-16]。有研究显示:利用poly(R)-Oct4、poly(R)-Sox2、poly(R)-Klf4和poly(R)-c-Myc的融合蛋白成功诱导并获得了小鼠[15]和人[16]的iPS细胞。这些研究提示,TAT和poly(9R)均可以将与之融合表达的诱导因子转导进入到靶细胞内实现诱导因子蛋白介导的细胞重编程。为此,本研究拟构建TAT-c-Myc-9R融合蛋白的原核表达载体,并对其进行表达分析,利用TAT和poly(9R)的双重跨膜作用将该蛋白转导进入细胞内,为以PTD介导的将该转录因子等直接重编程体细胞获得无外源基因整合牛iPS研究奠定基础。
1 材料与方法
1.1 材料与试剂
限制性核酸内切酶EcoRI、kpnI、xhoI、高保真PrimeSTAR· Max DNA Polymerase购自TaKaRa 公司;DNA Marker(DL2000和DL15000)购自天根生物公司,蛋白Marker(Protein Ruler II,12kDa-100kDa)购自北京全式金生物技术有限公司;pGEM-T Easy载体购自Promege公司;带有目的基因的原始质粒pMD18-T-bc-Myc由西北农林科技大学动物医学院吕长荣博士惠赠;表达的空质粒pTAT-HA由第四军医大学苟兴春博士惠赠;胶快速回收试剂盒、小量质粒抽提试剂盒、大肠杆菌(E.coli)JM109克隆菌株和BL21(DE3)PLyS.S表达菌株等购自北京全式金生物技术有限公司;其他普通试剂如Tris饱和酚和IPTG等,购自华美生物工程公司。
1.2 牛c-Myc基因的扩增
根据已发表的牛c-Myc基因序列(GenBank Accession No.NM_001046074.2)和原核表达空载体pTAT-HA,设计如下引物,由南京金斯瑞生物技术有限公司合成:Pro-bc-Myc-F:5'-GGGGTACCATGCCCCTCAACGTCAGCTTCG-3' (斜体加粗并带下划线部分表示KpnI 酶切位点),Pro-bc-Myc-R:5'-AAAAAACTCGAGGCGGCGTCTGCGTCTGCGGCGTCTGCGTTAGGCGCAAGAGTTCCG TATCTG-3'(上下游引物上斜体加粗部分分别表示KpnI和XhoI酶切位点,加粗下划线部分表示9R的编码序列的反向互补序列)。以pMD18-T-bc-Myc质粒为模板,利用以上引物进行PCR扩增,其反应体系如下(50 μL):ddH2O 22 μL,2×PrimeSTAR· Max缓冲液 (Mg2+ conc. (2×) = 2 mM; dNTP Mixture (2×)=0.4 mM each; PrimeSTAR Max DNA Polymerase, TaKaRa, R045A)25 μL,质粒DNA 1 μL,上游引物和下游引物各1 μL。根据高保真聚合酶特殊反应说明进行PCR扩增,然后,取PCR扩增产物,进行琼脂糖凝胶电泳检测,随后取45 μL PCR产物,与4 μL 的dNTPs(2.5 mM)和1 μLTaqDNA polymerase (5 U/μL)混匀,于PCR仪中72 ℃反应1 h,在PCR产物末端添加poly(A)尾巴。
1.3 重组表达载体pTAT-HA-bc-Myc-9R的构建
根据全式金的DNA快速胶回收试剂盒说明书操作回收bc-Myc-9R片段,并将其与pGEM-T Easy载体按3∶1 的摩尔比在T4 DNA连接酶作用下于4 ℃冰箱连接过夜,从而构建 pGEM-T-bc-Myc-9R重组克隆载体。将4 μL重组载体转化 40 μL感受态细胞 JM109,并在含有100 μL IPTG和 20 μL X-gal的氨苄青霉素琼脂糖平板上进行蓝、白斑筛选,然后,挑取部分白色菌落,进行PCR鉴定;PCR鉴定为阳性的克隆扩繁后提取质粒DNA(pGEM-T-bc-Myc-9R),进行EcoRI 酶切鉴定;PCR鉴定为阳性的克隆接种于含抗生素的LB液体培养基中200 r/min,37 ℃震荡培养 12~16 h。用全式金小提质粒抽提试剂盒提取质粒DNA,并送南京金斯瑞生物公司测序。用KpnI 和XhoI酶切测序正确的克隆载体pGEM-T-bc-Myc-9R获得带双酶切粘性末端的bc-Myc-9R基因片段。酶切体系如下(50 μL):5 μL 10×M buffer,2 μLKpnI 酶(10 U/μL),2 μLXhoI(10 U/μL)酶,2 μg 质粒,补水至50 μL并混匀后37 ℃孵育4 h。酶切产物经琼脂糖电泳后,通过DNA快速胶回收试剂盒回收。将带双酶切位点的目的片段克隆到具有相同酶切位点的pTAT-HA线性化载体上,构建重组表达载体 pTAT-HA-bc-Myc-9R,再利用KpnI 和XhoI酶进行双酶切鉴定。
1.4 重组表达载体pTAT-HA-bc-Myc-9R的诱导表达
将阳性重组质粒转化BL21(DE3)pLysS菌株,挑单菌落接种于含100 μg/mL 氨苄青霉素的3~5 mL LB液体培养基中,200~220 r /min,37 ℃过夜培养;次日,将过夜培养菌液按1∶100的比例接种到80 mL含有100 μg/mL 氨苄青霉素的 LB液体培养基,37 ℃振荡培养使OD600值达到0.6左右时,按照 5 mL/管分装菌液,设置终浓度分别为0.4,0.6,0.8和1.0 mmol/L 等4个IPTG浓度和4 h,6 h 和8 h 等3个时间梯度,37 ℃,200~220 r/min诱导表达。收集不同诱导条件下的菌液1.0 mL,10 000 r/min离心5 min;弃上清,沉淀中加入200 μL 1xSDS上样缓冲液彻底重悬菌体,沸水浴10 min,12 000 r/min离心15 min,取15 μL上清进行SDS-PAGE分析[8]。
2 结果与分析
2.1 pTAT-bc-Myc-9R原核表达载体的构建与鉴定
2.1.1bc-Myc-9R重组片段的获得 图1结果显示,目的基因bc-Myc-9R的片段大小为1 367 bp,与预期结果一致。
2.1.2 pGEM-T-bc-Myc-9R重组载体的菌液PCR鉴定 菌液PCR检测结果(图2)显示,所挑选的4个克隆均扩增到了1 367 bp的DNA片段,4个克隆均为阳性。
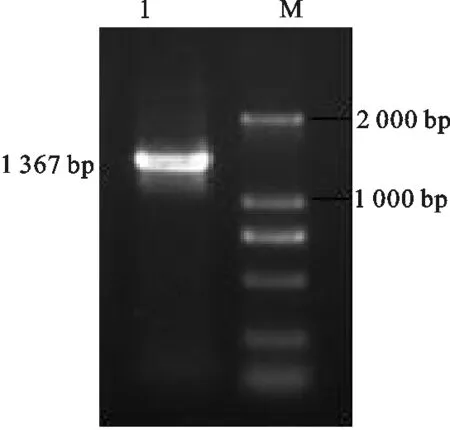
图1 bc-Myc-9R重组片段的PCR产物电泳检测 1.PCR扩增bc-Myc-9R重组片段;M.DNA marker D2000Fig.1 Detection of PCR product of bc-Myc-9R byagarose gel electrophoresis 1.Amplified fragment of bovine-c-Myc-9R gene by PCR;M.DNA marker D2000

图2 菌液PCR电泳检测1-4.挑选的克隆的PCR扩增产物;M.DNA marker D2000Fig.2 Detection of PCR product of bc-Myc-9R frombacteria by agarose gel electrophoresis 1-4.Amplified fragments of different clones;M. DNA marker D2000
2.1.3 pGEM-T-bc-Myc-9R重组质粒的EcoRI单酶切鉴定和测序鉴定 由于在pGEM-T easy 载体上有2个EcoRI的酶切位点,故pGEM-T-bc-Myc-9R重组质粒经EcoRI 单酶切获得了3 015 bp的pGEM-T片段和1 367 bp的bc-Myc-9R重组DNA片段(图3),进一步证实pGEM-T-bc-Myc-9R重组质粒构建成功。通过DNA测序鉴定,发现插入pGEM-T easy载体中的bc-Myc-9R的序列完全正确,确认pGEM-T-bc-Myc-9R重组质粒构建成功。
2.1.4 pTAT-bc-Myc-9R重组质粒的酶切鉴定分析 重组质粒pTAT-bc-Myc-9R的载体模式图和酶切鉴定结果如图4所示。由图4b可知, pTAT-bc-Myc-9R原核表达载体经KpnI和XhoI双酶切后获得了3 000 bp的pTAT-HA片段和1 367bp的bc-Myc-9R片段,与理论结果完全相符,证实原核表达载体pTAT-bc-Myc-9R构建成功。

图3 pGEM-T-bc-Myc-9R重组质粒的EcoRI酶切鉴定1-4.不同克隆质粒的EcoRI 的酶切产物;M.DNA marker D15000 Fig.3 EcoRI digestion of the recombinant plasmid ofpGEM-T and bc-Myc-9R gene 1-4.EcoRI digestion of the recombinant plasmid of pGEM-T- bc-Myc-9R from different clonies;M. DNA marker D15000
2.2 pTAT-bc-Myc-9R融合蛋白的原核表达
原核表达载体pTAT-bc-Myc-9R转化大肠杆菌细胞BL21(DE3)pLys S后,经不同浓度梯度的IPTG诱导后,发现IPTG的最佳诱导浓度为0.6 mM(终浓度)(结果未显示)。结果发现,未添加IPTG的 pTAT-bc-Myc-9R有少量的泄露表达。添加IPTG诱导表达的细菌细胞比未诱导的细菌细胞中目的蛋白的表达量明显增加。终浓度0.6 mM的IPTG 37℃诱导4 h和6 h均可很好的表达融合蛋白(如图5中红色箭头所示)。相同的IPTG的诱导浓度下,诱导6 h的细胞比诱导4 h的细胞中目的蛋白的表达量要大。
3 讨 论
哺乳动物c-Myc基因是最受科学家亲睐的原癌基因之一,其编码蛋白在人类基因组中大约有25000个结合位点。c-Myc蛋白的N端能够影响组蛋白乙酰化酶、ATP酶的作用,而其C端参与调节基因的表达[17]。1993年,c-Myc缺失的小鼠胚胎不能在妊娠中存活的报道,引起了科学家的关注,也开创了c-Myc与干细胞研究的先河。此后,c-Myc还被证明与细胞周期的调控相关,其通过若干种途径推进细胞从G1期进入S期,缩短了G1期,从而使细胞增殖速度加快,细胞周期缩短,进一步提示该基因与ES细胞或iPS细胞特性密切相关[18]。目前,对体细胞重编程过程中,c-Myc的作用有以下两种主要假设[19]:① c-Myc和Klf4使终末分化细胞产生肿瘤样转变,因此赋予了体细胞无限增殖以及ES细胞样的快速生长能力;② c-Myc可对体细胞染色质进行修饰,使得体细胞染色体结构变得松散,便于转录因子与重编程中所需的基因结合,从而调控基因表达。虽然c-Myc基因对iPS形成来说并不是必须的,但是,已有资料表明,不使用c-Myc的转录因子重编程研究中效率相对很低,而且需要的时间也更长一些。目前,关于牛体细胞重编研究甚少,为提高牛iPS研究的效率,同时缩短其周期,本研究进行了牛转录因子c-Myc基因的蛋白表达研究。

图4 重组质粒pTAT-bc-Myc-9R的酶切鉴定分析a.pTAT-bc-Myc-9R融合表达载体构建的模式;b.pTAT-bc-Myc-9R原核表达载体的KpnI和XhoI的双酶切鉴定;1.pTAT-bc-Myc-9R重组质粒;2,3.pTAT-bc-Myc-9R KpnI和XhoI的双酶切产物; M1.DNA marker D15000;M2.DNA marker D2000Fig. 4 Restriction enzyme analysis of pTAT-bc-Myc-9R recombined vectora.Schematic diagram of recombinant pTAT-bc-Myc-9R;b. Aagarose gel electrophoresis of the fragments of pTAT-bc-Myc-9Rdigested by KpnI and XhoI;1. plasmid DNA of pTAT-bc-Myc-9R; 2,3. digestion of pTAT-bc-Myc-9R recombinant plasmid by KpnI and XhoI;M1. DNA marker D15000 ; M2.DNA marker D 2000

图5 考马斯亮蓝染色检测pTAT-bc-Myc-9R的诱导表达1.0.6 mM IPTG 37 ℃诱导4 h 的结果;2.pTAT-bc-Myc-9R未诱导的结果;3.0.6mM IPTG 37 ℃诱导6 h 的结果;M.ProteinRuler II (12 kDa-100 kDa) Fig. 5 Detection of expression of pTAT-bc-Myc-9Rby Coomassie brilliant blue staining 1.expression of pTAT-b c-Myc -9R induced by 0.6 mMIPTG for 4 h;2. expression of pTAT-b c-Myc -9R withoutIPTG induction for culture 4 h;3.expression of pTAT-b c-Myc -9Rinduced by 0.6 mM IPTG for 6 h; M. Protein Ruler II
由于原癌基因c-Myc的特殊身份,为了避免其对靶细胞的整合,故本研究首先表达c-Myc蛋白,以便于后续的利用蛋白转导靶细胞以实现转录因子蛋白直接重编程黄牛成纤维细胞的研究。但是,正常情况下,绝大部分蛋白不能够穿透细胞膜进入细胞内,而特殊小肽分子PTD(如:TAT和9R) 则既能携带核酸也能携带蛋白穿过细胞膜进入到细胞内[20-25]。人类iPS的研究中也证明,TAT介导的TAT-转录因子融合蛋白能够进入到细胞内[25]。2009年,Kim等[26]构建了稳定表达9R-转录因子融合蛋白的真核表达细胞系(4个HEK293细胞系),并通过这些细胞系的细胞提取物成功诱导了人成纤维细胞重编程。目前,关于TAT、11R和9R的研究报道依然不少。刘雪梅等[10]利用TAT肽获得TAT-Apoptin 融合蛋白并检测到活性;徐丽娟等[11]利用11R获得人类Oct4和Sox2的融合表达产物;姜扩等[13]利用9R获得人抗HER2单链抗体/精氨酸九聚体融合蛋白;丁道芳等[12]利用9R获得具有穿透性的人类Sox2融合蛋白;2013年,刘源等[14]发现9R穿膜肽可增强P53抗肿瘤效果。然而,截止目前,同时利用TAT和多聚精氨酸(如:9R或8R) 介导牛靶蛋白进入细胞内并发挥功能的研究尚未见报道。
为此,本研究同时考虑TAT和9R,进行牛c-Myc基因和TAT、9R的融合蛋白表达研究,从而实现牛体细胞重编程。本试验的具体思路如下:直接设计只添加有相应酶切位点而不含TAT标签的引物,从已有的含有目的基因的质粒中扩增目的基因,通过简单的酶切,直接连接到多克隆位点上游含有TAT和HA融合表达标签的pTAT-HA质粒,减少了多次扩增可能导致基因突变的概率。此外,在c-Myc的C端和N端分别添加了均有跨膜转导功能的TAT和9R。通过一系列的分子生物学和基因工程操作,成功克隆得到了重组质粒pTAT-bc-Myc-9R,通过对IPTG的诱导浓度和诱导时间等诱导条件的优化,确定了最佳的诱导条件为终浓度0.6 mmol/L的IPTG 37 ℃诱导6 h。虽然未添加IPTG的 pTAT-bc-Myc-9R有少量的泄露表达,可能跟培养基中残存有少量的乳糖(最后转化成半乳糖而诱导目的蛋白少量的表达)有关,此现象跟梁英民等[26]的研究报道一致。但是从BL21(DE3)pLys S自身的背景条带对比可以看出,添加IPTG诱导表达的细菌比未诱导的细菌中目的蛋白的量明显增加。此外,由于TAT-bc-Myc-9R融合蛋白中含有6个His标签蛋白和9个精氨酸,这些碱性氨基酸导致此融合蛋白在SDS-PAGE中迁移变慢,从而出现分子量偏大的现象;这与唐威华等[27]的研究结果相似。这些研究结果可为后续利用TAT-转录因子融合蛋白直接诱导获得黄牛iPS细胞奠定了基础。
参考文献:
[1] Nilsen T W, Maroney P A.Translational efficiency of cMyc mRNA in Burkitt lymphoma cells[J]. Mol Cell Biol, 1984, 4(10):2 235-2 238.
[2] Shortt J, Martin B P, Newbold A, et al. Combined inhibition of PI3K-related DNA damage response kinases and mTORC1 induces apoptosis in MYC-driven B-cell lymphomas[J].Blood, 2013,121(15):2 964-2 974.
[3] Rezanejad H, Matin M M. Induced pluripotent stem cells: progress and future perspectives in the stem cell world [J].Cell Reprogram, 2012, 14(6):459-470.
[4] Bellin M, Marchetto M C, Gage F H, et al. Induced pluripotent stem cells: the new patient[J].Nat Rev Mol Cell Biol, 2012, 13(11):713-726.
[5] Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors [J]. Cell, 2006,126(4):663-676.
[6] Mandal P K, Rossi D J.Reprogramming human fibroblasts to pluripotency using modified mRNA[J].Nat Protoc, 2013, 8(3):568-582.
[7] 潘传英, 陈 宏, BISHOP E.Colin.利用诱导因子Oct4、Sox2和SV40 T的mRNA介导体细胞重编程研究 [J]. 畜牧兽医学报, 2012, 43(11): 1 733-1 739.
[8] Pan C, Lan X, Chen H, et al. An economical single-sided antibody incubation method for western blotting [J]. J Virol Methods, 2010, 169:409-411.
[9] Regberg J, Eriksson J N, Langel U.Cell-penetrating peptides: from cell cultures to in vivo applications [J].Front Biosci (Elite Ed), 2013, 5:509-516.
[10] 刘雪梅,崔剑,侯伟健,等.TAT-Apoptin 融合蛋白分泌表达载体的构建及其活性检测[J].中国医科大学学报, 2013,42(1): 45-48.
[11] 徐丽娟,安群星,陈晓鹏.带有穿膜结构域的转录因子蛋白Oct4和Sox2的融合表达[J].生物技术通讯, 2012, 23(5):680-682.
[12] 丁道芳, 王瑧, 李晓锋,等. 穿透性Sox2蛋白的质粒构建及功能鉴定[J].中国药理学通报, 2012, 28(12): 1 671-1 674.
[13] 姜 扩,陈 锐, 王立锋, 等. 人抗HER2单链抗体/精氨酸九聚体融合蛋白的基因构建、表达及鉴定[J].细胞与分子免疫学杂志, 2011, 27( 11): 1 201-1 203.
[14] 刘 源,陈 睿,张 楠, 等. 9R穿膜肽可增强P53 抗肿瘤效果[J].生物工程学报, 2013, 29(7): 955-964.
[15] Zhou H, Wu S, Joo J Y, et al. Generation of induced pluripotent stem cells using recombinant proteins [J]. Cell Stem Cell, 2009, 4(5):381-384.
[16] Kim D, Kim C H, Moon J I, et al. Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins[J]. Cell Stem Cell, 2009, 4(6):472-476.
[17] Prochownik EV.c-Myc: linking transformation and genomic instability [J].Curr Mol Med, 2008, 8(6):446-458.
[18] Hydbring P, Larsson L G.Cdk2: a key regulator of the senescence control function of Myc[J].Aging (Albany NY),2010,2(4):244-250.
[19] Heffernan C, Sumer H, Malaver-Ortega L F, et al. Temporal Requirements of cMyc Protein for Reprogramming Mouse Fibroblasts[J].Stem Cells Int,2012, Article ID 541014, 12 pages. doi:10.1155/2012/541014.
[20] Wadia J S, Stan R V, Dowdy S F. Transducible TAT-HA fusogenic peptide enhances escape of TAT-fusion proteins after lipid raft macropinocytosis [J]. Nat Med 2004, 10:310-315.
[21] DeRouchey J, Hoover B,Rau D C. A comparison of DNA compaction by arginine and lysine peptides: a physical basis for arginine rich protamines [J].Biochemistry,2013, 52(17):3 000-3 009.
[22] Kanwar J R, Gibbons J, Verma A K, et al. Cell-penetrating properties of the transactivator of transcription and polyarginine (R9) peptides, their conjugative effect on nanoparticles and the prospect of conjugation with arsenic trioxide [J].Anticancer Drugs,2012,23(5):471-482.
[23] DeRouchey J E, Rau D C.Role of amino acid insertions on intermolecular forces between arginine peptide condensed DNA helices: implications for protamine-DNA packaging in sperm [J].J Biol Chem,2011, 286 (49): 41 985-41 992.
[24] Kwon H Y, Eum W S, Jang H W, et al. Transduction of Cu, Zn-superoxide dismutase mediated by an HIV-1 Tat protein basic domain into mammalian cells [J]. FEBS Lett ,2000,485(2-3):163-167.
[25] Pan C, Lu B, Chen H, et al. Reprogramming human fibroblasts using HIV-1 TAT recombinant proteins Oct4, Sox2, Klf4 and c-MYC [J]. Mol Biol Rep, 2010, 37:2 117-2 124.
[26] 梁英民,孙 强,蒋姗姗,等.PTD-bcr/abl融合癌蛋白片段的表达及其跨膜转运[J].细胞与分子免疫学杂志,2001,17(4):377-380.
[27] 唐威华,张景六,王宗阳,等.SDS-PAGE法测定His-tag融合蛋白分子量产生偏差的原因[J].植物生理学报, Acta Phytophysiologica Sinica, 2000,26(1):64-68.
