无患子种质资源多样性与亲缘关系的ISSR 和SRAP 分析
2013-11-08柏明娥张加正洪利兴沈建军
洪 莉,柏明娥,张加正,洪利兴,沈建军
(1.浙江省台州市农业科学研究院,浙江 临海 317000;2.浙江省林业科学研究院,浙江 杭州 310023)
无患子 (Sapindus mukorossi Gaertn.)又名肥皂树,为无患子科无患子属落叶乔木树种,广泛分布我国东部、南部至西南各省区,日本、朝鲜、越南、老挝、柬埔寨、缅甸、泰国、马来西亚、尼泊尔和印度也有分布[1]。近年来研究表明,无患子果皮含有四环三萜类大戟烷型 (Tirucallane type)和达玛烷型 (Dammarane type)等多种皂苷[2-4],可作为天然活性物质用于洗涤和洗发香波及各种洁肤护肤化妆品中,具有抗皮真菌和念珠菌等抗菌、止痒、治疗脚癣和轮癣等功效[5],是当前日用化工业中非常引人关注的绿色洗涤生物化工原料,市场前景广阔。目前,人工种植的无患子大多使用天然种源的实生苗,良种选育尚处于起步阶段。辜夕蓉[6]对来自于四川和云南5个地方的无患子种源的种子品质进行研究和比较,以期从中找寻出优良种源,为无患子的栽后产量和品质打下基础。邵文豪等[7-8]通过对不同产地无患子果皮皂苷含量的测定分析和收集无患子自然分布区内不同种源、家系种子进行播种育苗定期观测,研究无患子不同种源皂苷含量和苗期生长变异规律与种源地生态因子之间的关系,以期为选择高皂苷含量的优良种源区划及遗传改良提供理论依据。
简单重复序列区间 (inter-simple sequence repeat,ISSR)是1994年由Zietkeiwitcz 创建的一种简单序列重复间扩增多态性分子标记方法[9],相关序列扩增多态性 (sequence-related amplified polymorphism,SRAP)是由美国加州大学Li 和Quiros 于2001年发展的一种基于PCR 反应的新型标记[10]。利用分子标记技术研究无患子种质资源遗传多样性的相关文献鲜见报道。本研究采用ISSR 和SRAP 分子标记技术,选取来自浙江、湖南、四川等地的16 份无患子材料,通过探讨不同产地无患子种源间的遗传差异和亲缘关系,旨在为科学实现远缘杂交或远亲杂交提供理论依据。
1 材料与方法
1.1 供试材料
2011年11-12月,在无患子不同分布区选择树龄20年以上、胸径18 cm 以上且生长正常的无患子采集成熟果实,共收集13个产地16 份果实材料 (表1)。果实采集带回室内,将种子剥离后湿沙埋藏,2012年3月进行播种试验。试验在浙江省台州市农业科学研究院大棚内进行。台州属亚热带季风性湿润气候,四季分明,年均气温17℃,年降水量1 550 mm。土壤为砂壤土,pH 值6.1,地势平坦,水源充足。2012年9月采集当年生幼树嫩叶作分析测试样品。
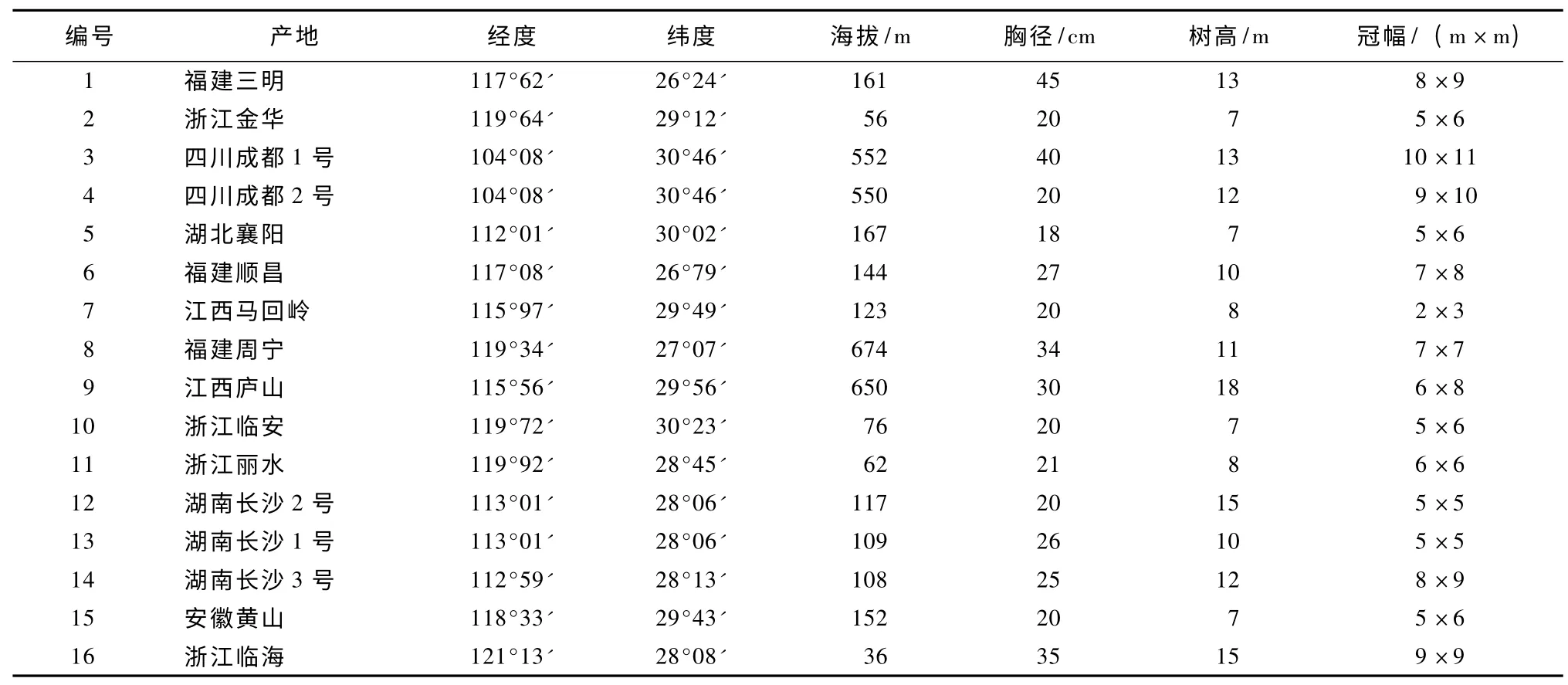
表1 供试无患子材料的情况
1.2 处理方法
1.2.1 无患子基因组DNA 的提取
基因组DNA 提取采用SDS-CTAB 法,DNA 粗提物再经Magabio 核酸纯化试剂盒进行纯化后,通过1.5%琼脂糖凝胶电泳和DNA/RNA 紫外分光光度计 (GeneQuant Pro,GE Healthcare)检测完整性、纯度及浓度。D260/D280>1.8 的DNA 样品用于后续PCR 扩增。
1.2.2 ISSR 引物的选用和PCR 的扩增
ISSR 和SRAP 通用引物如表2 所示。

表2 ISSR 和SRAP 通用的引物序列
ISSR 和SRAP 扩增反应均采用20 μL 体系。2.0 μL 10× PCR Buffer,2.0 μL dNTP (each 2.5 mmol·L-1),1.2 μL MgCl2(25 mmol·L-1),0.2 μL Taq DNA 聚合酶(1U),模板DNA 60 ng,ISSR 引物1 μL (10 μmol·L-1)或SRAP 上下游引物各1 μL (10 μmol·L-1),dd H2O 补足20 μL。
PCR 扩增反应在杭州博日公司的TC-XP 型扩增仪上进行。ISSR-PCR 扩增程序为94℃预变性4 min;94℃变性1 min,56℃退火30 s,72℃延伸2 min,35个循环,循环结束后72℃延伸7 min,4℃保存。SRAP-PCR 扩增程序为94℃变性1 min,35℃退火1 min,72℃延伸1.5 min,共5个循环,随后94℃变性1 min,50℃退火1 min,72℃延伸1.5 min,30个循环结束后72℃延伸8 min,4℃保存。
1.2.3 数据处理与统计分析
PCR 扩增产物用1.5% 琼脂糖凝胶电泳,用Quantity One 6.0 分析软件结合人工方法读带,记录电泳图谱中清晰且能重复出现的条带,在相同的迁移位置上,有带用1 表示,无带用0 表示。采用NTSYS-pc2.10e 软件的Simqual 程序计算样品间的Dice 相似系数,同时用类平均聚类法 (UPGMA)进行聚类分析,构建聚类图。
2 结果与分析
2.1 无患子种质的ISSR 和SRAP 分析
用优选出的13 条ISSR 随机引物和3 对SRAP引物 (Me3-em7、Me3-em15、Me3-em17)对无患子基因组DNA 进行PCR 扩增,结果均能扩展出清晰稳定、重复性和多态性较高的条带 (图1-2)。不同引物扩增的DNA 片段为300~1 300 bp,DNA片段数目为2~12 条,平均每个引物扩增出5 条主带,多态性条带比率约为61%。
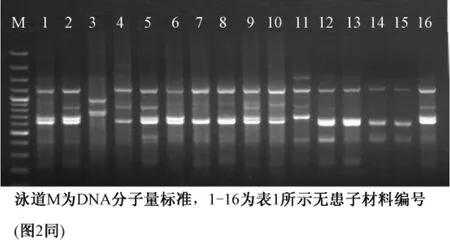
图1 ISSR 引物UBC862 的PCR 扩增图谱
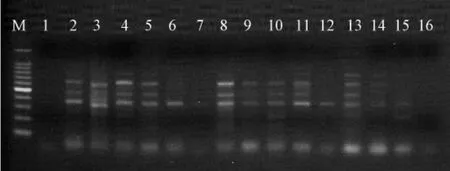
图2 SRAP 引物对Me3-em7 的PCR 扩增图谱
2.2 遗传相似系数的估算和聚类分析
图3 为通过Dice 相似系数利用UPGMA 法构建的不同产地间无患子种质聚类分析树状图。
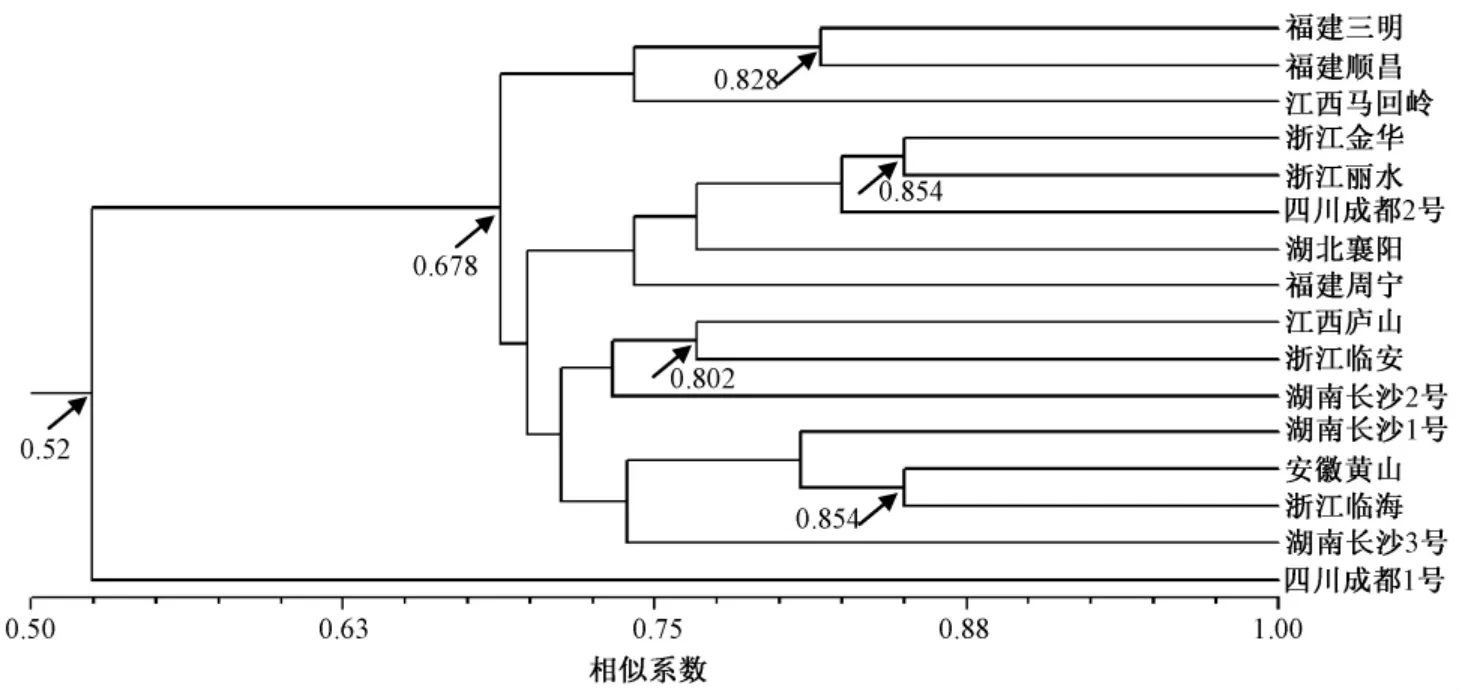
图3 16个无患子种质的UP GMA 聚类树
聚类结果显示,当Dice 相似系数为0.52 时,可将供试的无患子种质分为2个大类,第1 大类包括了15个无患子种质,第2 大类仅为四川成都1号1个种质;当Dice 相似系数为0.678 时,可以将第1 大类再分为2个亚类,第1 亚类包括来自福建和江西的3个无患子种质,第2 亚类包括来自浙江、湖南、湖北、安徽、四川、江西、福建的12个无患子种质。在第1 亚类中,来自福建三明和福建顺昌的无患子种质以0.828 的Dice 相似系数聚在一起;在第2 亚类中,来自浙江金华和浙江丽水的无患子种质以0.854 的Dice 相似系数聚在一起,来自江西庐山和浙江临安的以0.802 的Dice 相似系数聚在一起,来自安徽黄山和浙江临海的以0.854 的Dice 相似系数聚在一起。从以上聚类结果可以看出,各无患子种质并未严格按地理界限聚在一起,仅部分来源于同一地区的无患子种质聚在一起,出现“大杂居、小聚居”的现象。
3 小结和讨论
利用ISSR 和SRAP 分子技术对16个地区无患子种质进行遗传多样性分析,PCR 扩增图谱上存在明显的多态性,说明利用ISSR 和SRAP 标记可以从分子水平上揭示出不同产地无患子的基因组差异,在无患子种质鉴定和遗传多样性研究上具有重要价值。本研究中无患子种质的多态性比率约为61%,表明分布于不同产地的无患子具有较高的遗传多样性,各分布区的个体遗传差异较大,这可能与其具有很强的适应不同气候环境的能力相关。
在亲缘关系上,遗传距离越小,遗传一致性越大,材料间的亲缘关系也就越近,反之,亲缘关系就较远。在本研究的16个无患子种质中,亲缘关系最近的是在第2 亚类中聚在一起的分布于浙江丽水和金华的种质,以及分布于安徽黄山和浙江临海的种质。其次,分布于福建三明和顺昌的种质,以及分布于江西庐山和浙江临安的种质也有着较近的亲缘关系,这些分布区的地理距离相对较近。亲缘关系最远的是分布于福建三明和四川成都的种质,相对地理距离较远。此外,基于2种分子标记得到的无患子种质聚类结果并不与其地理分布一致,如同是分布于四川成都和湖南长沙的种质并未聚在一起,显示了较大的基因组差异,表明无患子种质的遗传变异与地理分布没有呈现明显的相关性。
无论ISSR 和SRAP 分子图谱还是分子聚类结果,均显示四川成都1号种质与其他种质在分子水平上存在明显差异,这为我们后续在无患子的杂交育种计划中亲本选择提供了优先选择的材料。
[1]中国植物志编辑委员会.中国植物志 四十七卷 (第一分册)[M].北京:科学出版社,1998.
[2]Kimata H,Nakashima T,Kokubun S,et al.Saponins of pericarps of Sapindus mukurosii Gaerth.and Solubilization of monodesmoides by bisdesmosides [J].Chem Pharm Bull,1983,31 (6):1998-2005.
[3]Nakayama K,Fujino (nee Kimata) H,Kasai R,et al.Solubilizing properties of saponins from Sapindus mukurossi Gaerth[J].Chem Pharm Bull,1986,34 (8):3279-3283.
[4]Huang H C,Tsai W J,Morris-Natschke S L,et al.Sapinmusaponins F-J,bioactive tirucallane-type saponins from thf galls of Sapindus mukorossi [J].J Nat Prod,2006,69(5):763-767.
[5]孙洁如,陈孔常,周鸣方,等.无患子表面活性物及其复配体系的性质研究[J].日用化学工业,2002,32 (4):16-18.
[6]辜夕容.不同种源无患子的种子品质差异分析[J].西南大学学报:自然科学版,2009,31 (6):51-54.
[7]邵文豪,姜景民,董汝湘,等.不同产地无患子果皮皂苷含量的地理变异研究[J].植物研究,2012,32 (5):627-631.
[8]邵文豪,岳华峰,姜景民,等.不同种源无患子苗期生长性状遗传变异研究[J].浙江林业科技,2012,32 (1):21-25.
[9]Zietkiewicz E,Rafalski A,Labuda D.Genome fingerprinting by simple sequence repeats (SSR)-anchored PCR amplification[J].Genomics,1994,20:176-183.
[10]Li G,Quiros C F.Sequence-related amplified polymorphism(SRAP),a new marker system based on a simple PCR reaction:its application to mapping and gene tagging in Brassica[J].Theoretical and Applied Genetics,2001,103:455-461.
