植被类型变化对长白山森林土壤碳矿化及其温度敏感性的影响
2013-09-19吕瑜良徐志伟王瑞丽何念鹏
王 丹,吕瑜良,徐 丽,何 秀,徐志伟,赵 宁,王瑞丽,何念鹏,*
(1.西南大学地理科学学院,重庆 400715;2.中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室,北京 100101;3.华中农业大学资源与环境学院,武汉 430070;4.中国科学院沈阳应用生态研究所,沈阳 110016)
森林生态系统是陆地生态系统最重要的碳库[1],森林土壤碳收支取决于新有机质输入和原土壤有机质的分解这两个重要过程,任何影响上述两个过程的因素均会对森林生态系统碳源汇功能产生重要影响[2]。原生林遭受干扰转变为次生林的过程中,植被类型以及生境条件发生动态演替,这种变化严重影响了森林碳循环过程的变化,进而对气候变化产生影响[3],因此,准确估计植被类型变化(或土地利用变化)对森林生态系统碳收支及其关键参数的影响是当前全球气候变化研究的重点内容之一[4-5]。
土壤有机质分解受多种环境因素的影响,其中最主要的是土壤温度和含水量[6-7]。大量研究已经表明温度和水分对土壤有机质分解具有重要影响[8-9],且存在显著的交互效应[10-11];弄清温度和水分对特定森林土壤有机质分解的影响及其交互效应,对揭示该森林生态系统碳循环至关重要。然而,在实地测定土壤碳矿化(或土壤呼吸)时,容易受到其它环境因素的交互影响,很难准确的将温度和水分对土壤碳矿化的影响区分开来;因此,室内培养就被广泛地应用于土壤有机质分解的机理性研究[12-14]。
本文以长白山地区典型的针阔混交林为研究对象,分别将其原生林和次生林的土壤进行不同温度和不同水分的室内培养,并定期测定其土壤碳矿化速率。本文的主要目的:1)探讨植被类型变化(原生林VS次生林)对森林土壤碳矿化及其温度敏感性的影响?2)揭示温度和水分对森林土壤碳矿化的交互影响?
1 试验样地及方法
1.1 试验样地概况
研究地点位于长白山国家自然保护区以及吉林省安图县二道白河镇境内,它东边受太平洋的影响,气候潮湿多雨,为典型的大陆性温带季风气侯,年平均气温为-4.8—2.9℃,年降水量为600—900 mm。全年日照时数为2271—2503 h,无霜期109—141 h[15],土壤为山地暗棕色森林土。
实验样地为中国科学院长白山森林生态系统定位研究站的长期实验样地,分别为原生的针阔混交林(地理坐标128°5'33″E,42°24'18″N)和砍伐后形成的次生混交林(地理坐标128°4'18″E,42°37'43″N)。针阔混交林样地为原生林,它是长白山地区的地带性植被类型,主要树种有红松(Pinus koraiensis)、春榆(Ulmus davidiana var.japonica)、紫椴(Tilia amurensis)、槭类等;次生林是原生林(针阔混交林)在皆伐后形成的次生林(—20a),主要树种为蒙古栎(Quercus mongolica)、红松(P.koraiensis)、山杨(Populus davidiana)、栎类等,草本植物较发达[15-16],调查的样方情况如下(表1)。

表1 调查样方植被信息Table 1 Vegetation characteristics of the experimental plots
1.2 样地设置及研究方法
2012年8月,根据地形和地势的情况,分别在原生林和次生林建立了4个30 m×40 m样地。每个样地,设置30—40个采集点,在去除地表凋落物后,采用土钻法对0—30 cm土壤样品进行取样;多个采集点的土壤样品形成一个混合土壤样品(>5 kg);即分别在原生林和次生林获得4个混合土壤样品。取样后,在室内对土壤样品进行过筛处理(2 mm土壤筛),并手工挑除根系和杂质。经过预处理的少量土壤样品(约100 g)风干处理,其余部分4℃冷藏。
1.3 室内测试与培养
培养实验共设置3个水分梯度(30%、60%、90%土壤饱和含水量(SSM))和6个温度梯度(5、10、15、20、25和30℃),4次重复,共计144个培养样品。其中,土壤饱和含水水量采用简易法进行测定,重复3次,详细方法可参见文献[16]。
培养实验开始前,先采用烘干法测定土壤含水量,经过含水量换算后,再称取土壤鲜样40 g和石英砂10 g,装入150 mL塑料圆瓶,摇匀后加蒸馏水分别调节至30%,60%和90%SSM。样品先在20℃培养1周,并测定土壤碳矿化速率。随后,培养样品分别放入5、10、15、20、25和30℃的恒温恒湿培养箱内进行培养。
在8周测定期内,土壤碳矿化释放速率共测定9次,分别为培养1、3、5、7、15、21、28、42和56 d时测试。在培养过程中,每隔2—3 d采用称重法给土壤样品补水1次。土壤碳矿化速率采用自主研发的土壤微生物呼吸速率测定系统进行测定,详见代景忠等[17]。
土壤全碳和全氮含量采用元素分析仪测定,土壤pH值利用pH计测定,土壤电导率使用电导仪测定。实验样地的土壤性质见表2。

表2 土壤理化特性Table 2 Soil properties of experimental plots
1.4 数据处理
土壤碳矿化速率采用如下公式进行计算[17]:

式中,R为土壤微生物呼吸速率(μgC g-1d-1),C为测试时间内CO2浓度变化的直线斜率,V是培养瓶和管线的总体积,m是培养瓶内土壤干重,α是CO2气体质量转化系数,β是时间转化系数。
Q10值采用指数模型关系进行计算:

式中,a,b为参数。
建立双因素关系模型探讨温度和水分对土壤碳矿化的影响:

式中,R为土壤碳矿化累积量,T为培养温度,w为土壤含水量,a、b、c、d、e为参数。
利用SPSS 13.0统计软件包对实验样地各类指标进行成对T检验,采用单变量多因素分析方法检验了温度、水分和植被类型对土壤碳矿化及其温度敏感性的影响,显著性差异水平为 P=0.05。利用Sigmaplot 10.0拟合土壤碳矿化速率随着培养温度和水分的变化趋势。
2 结果与分析
2.1 植被类型变化对土壤碳矿化累积量的影响
植被类型变化对土壤碳矿化累积量具有显著影响(F=3505.19,P<0.001)(表3)。在相同温度下,次生林的土壤碳矿化累积量显著高于原生林,温度越高这种差距越明显(图1)。以90%SSM为例,整个培养期内(56 d),温度为5℃时,次生林土壤碳矿化累积量为110.51 μgC/g,而原生林土壤碳矿化累积量为101.28 μgC/g;当培养温度为30℃时,次生林土壤碳矿化累积量(346.41 μg C/g)显著高于原生林(241.01 μgC/g)。

图1 土壤碳矿化累积量的动态Fig.1 Dynamics of soil C mineralization in the 56 d incubationSSM:土壤饱和含水量

表3 植被类型、培养温度、水分和培养时间对土壤碳矿化速率的影响Table 3 Effects of vegetation type,incubation temperature,incubation moisture and time on soil C mineralization rate
2.2 温度和水分对土壤碳矿化的影响
土壤温度(F=3508.19,P<0.001)和水分(F=893.68,P<0.001)对土壤碳矿化速率有显著影响(表3)。土壤碳矿化累积量与温度成正相关,在不同含水量状况下土壤矿化累积量均随温度升高而增多,30℃时达到最高(图1)。次生林和原生林的土壤碳矿化速率对温度变化的响应强度存在差异;在不考虑水分的状况下,次生林土壤碳矿化速率的上升趋势显著高于原始林(图2)。
土壤含水量对土壤碳矿化速率的影响极显著(P<0.001),相同水分条件下,次生林土壤碳矿化速率显著高于原始林(图3)。在低温时(<15℃),60%SSM的原始林土壤碳矿化速率最高,当温度超过15℃时,原始林的土壤碳矿化速率随水分上升而上升。而在低温时(<25℃),次生林土壤碳矿化速率在60%SSM时相对低,而在90%SSM时略高;而当温度为30℃时,土壤碳矿化速率随水分升高而升高。

图2 温度对土壤碳矿化速率的影响Fig.2 The effect of temperature on the rates of soil C mineralization rate

图3 水分对土壤呼吸速率的影响Fig.3 The effect of incubation water on soil C mineralization rate
2.3 土壤温度和湿度对土壤碳矿化的模型拟合结果
本文采用双因素关系模型R=(a×ebT)×(c×w2+d×w+e)来分析土壤温度和湿度对土壤碳矿化累积量的影响。实验结果表明:培养7 d,土壤温度和湿度共同解释了土壤碳矿化能力的85.7%—95.9%,而培养56 d,温度和水分共同解释了土壤碳矿化的82.7%—89.2%(图4,表4)。此外,模型中b为温度敏感性指数,而c为水分敏感性指数;由表3可知短期培养期内(培养7 d)次生林土壤碳矿化对温度变化比较敏感,而原生林土壤碳矿化更易受土壤含水量的影响。在经过较长时间培养后(培养56 d),根据对b和c分析可以看出,次生林的土壤碳矿化对温度和湿度的交互影响更为敏感(表4)。
2.4 Q10与植被类型和水分的关系
植被类型对土壤碳矿化的温度敏感性(Q10)有显著影响(F=483.81,P<0.001),次生林的Q10值显著高于原始林(图5)。水分对土壤碳矿化的温度敏感性Q10具有显著影响(F=105.59,P<0.001),且植被类型和水分二者间存在显著的交互影响(P<0.001)(表4)。培养前7 d,次生林的Q10值随水分显著升高,而到了56 d时则表现为60%SSM最高(1.61)。在原始林,在培养7 d时,30%SSM时Q10值为1.26,60%SSM时Q10值为1.23,而90%SSM时Q10值为1.46;在培养56 d时Q10随培养水分的升高而升高。

图4 温度和水分对土壤碳矿化影响的模型模拟Fig.4 The effect of water and temperature on soil C mineralization
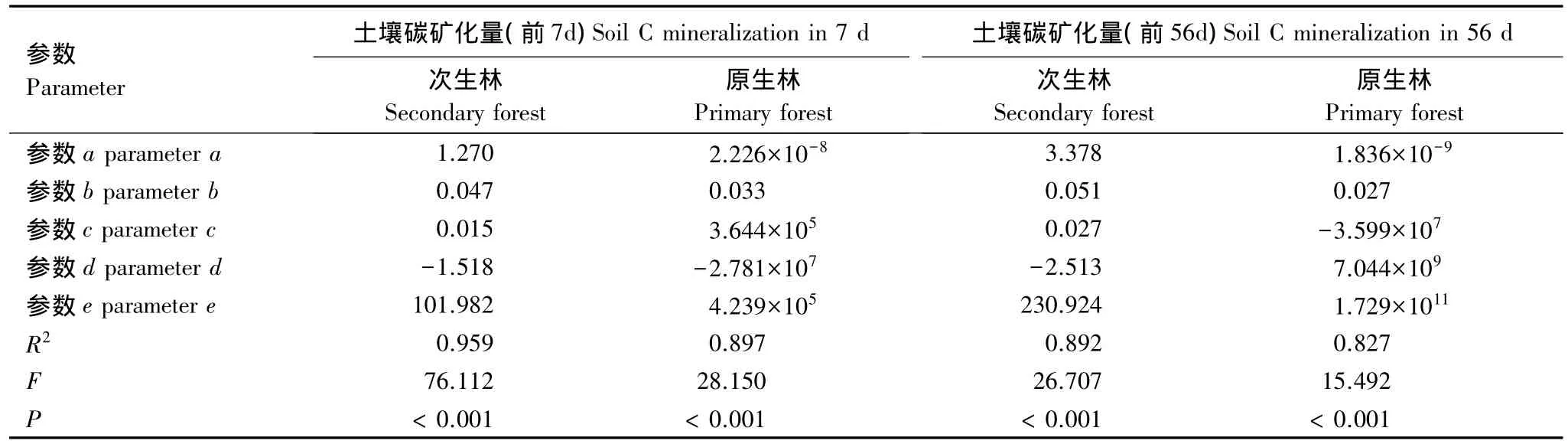
表4 土壤碳矿化与培养温度和水分模拟模型的主要参数Table 4 Model parameters of soil C mineralization with incubation moisture and incubation temperature

表5 植被类型和水分对土壤碳矿化的温度敏感性(Q10)的影响Table 5 The effect of vegetation types and incubation moisture on the temperature sensitivity(Q10)of soil C mineralization
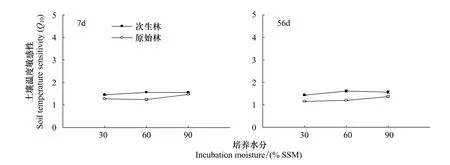
图5 土壤碳矿化温度敏感性Q10随水分的变化Fig.5 The effect of moisture on the temperature sensitivity(Q10)of soil C mineralization
3 讨论
3.1 植被类型变化与土壤碳矿化
次生林的土壤碳矿化累积量显著高于原始林,意味着植被类型对土壤碳矿化具有显著影响。这与其他学者所得出的结论相似[18-19],研究发现森林生态系统中的原生林经过人为干扰转变为次生林后土壤呼吸增强[3,20],产生这种变化是因为次生林是原生林在遭受认为干扰后形成的更新林,林分种类较原生林简单,群落分盖度低于原生林,从而导致其林内气候更易受到外界影响,进而影响土壤有机质的分解速率;此外,森林演替过程中植被群落结构、类型和数量不断变化,并和周围环境和土壤相互作用,形成不同土壤环境[21],进而引起微生物活动过程发生变化,或形成不同的土壤的物理性质 (孔隙、团聚体)[21-23]。因此,进一步深入研究植被类型对土壤碳矿化的影响及其机理对于研究森林碳循环对气候变化的响应非常必要。
3.2 土壤碳矿化对温度和水分变化的响应
温度和水分是影响森林生态系统土壤碳矿化的主要因素,土壤碳矿化的大部分变化是由温度和湿度引起的[9,24-25]。从本文分析可知,温度对土壤碳矿化有显著影响,原生林和次生林的土壤碳矿化能力都随温度上升,这与前期的研究结论一致[26],引起这种变化的主要原因可能是温度上升会促使土壤中微生物、土壤中酶的活性和根的呼吸增强等[24]。
水分对原生林和次生林土壤碳矿化有着显著影响,原始林在低温时,60%SSM的土壤碳矿化速率最高,当温度超过15℃时,原始林的土壤碳矿化速率随水分上升而上升。次生林土壤碳矿化能力都表现为90%SSM时最高,大量研究表明:含水量为60%—70%之间的水分最利于土壤呼吸作用的进行,土壤水分或过低过多会抑制土壤CO2的释放量[27],含水量过高会降低土壤的空隙和氧气含量,从而抑制土壤微生物呼吸与气体交换过程[28],而含水量过低时微生物和酶的活性会降低,不利于土壤呼吸作用的进行。这与次生林在低温时所得出的结论相似,温度超过15℃时,水分上升,碳矿化速率加快,其原因可能是温度超过15℃时温度和水分交互影响时温度的作用占主导地位,而原生林土壤碳矿化速率在90%SSM时最高,其原因可能是由于水分较高时交互效应影响引起的。
双因素关系模型R=(a×ebT)×(c×w2+d×w+e)进行分析可以很好地解释土壤碳矿化的变异,也说明温度和水分是土壤碳矿化的重要影响因素。在培养7 d时,土壤温度和湿度共同解释了长白山森林土壤碳矿化能力的85.7%—95.9%,而在培养56 d时,温度和水分共同解释了森林土壤碳矿化的82.7%—89.2%。Rey等在研究指出温度和湿度共同解释栎树矮林土壤呼吸年变化的91%[29],Qi等采用双因素关系模型发现温度和水分共同解释了不同森林土壤呼吸季节变化的80%—96%[30]。而整个培养期内,由参数b和c分析可知,次生林的参数b值(温度敏感性因子)显著大于原始林,而原生林的c值(水分敏感性因子)大于次生林,但温度和水分的交互影响结果表现为原始针阔混交林的土壤碳矿化累积量显著高于次生针阔混交林,这其中的原因可能是温度对原始针阔混交林和次生混交林的影响力大于水分。土壤碳矿化受温度和水分的影响程度不同,因此采用单因素模型来解释温度和水分对土壤碳矿化或土壤呼吸的结果时要更加谨慎,未来应进一步加强水分和温度对土壤碳矿化的交互效应的研究。
3.3 Q10及其主要影响因素
植被类型和土壤水分对原始林和次生林土壤碳矿化的温度敏感性(Q10)都具有显著影响,且这两种因素之间存在着显著的交互效应。培养7 d和培养56 d,无论是原始的针阔混交林还是次生的针阔混交林,植被类型变化都对其土壤碳矿化有着显著影响,相同水分条件下次生林的Q10值大于原始林,这种现象的形成可能是因为不同类型森林土壤中的底物对温度的不同反应所引起的,还有可能是其土壤微生物群落差异所导致的[31-32]。不同森林类型间的Q10值存在着相当大的差异,说明植被类型变化会强烈影响土壤碳矿化及其对温度变化的敏感程度[33]。植被类型发生变化会对土壤有机碳的数量与结构产生明显的影响、并改变植被类型以及生物量、植被盖度、根系分布和微生物种类和数量等[34],从而导致Q10值的巨大变异[33];未来应仔细深入开展底物和土壤微生物是如何影响两种森林类型的温度敏感性,为揭示该地区的碳周转提供理论依据。
整个培养期内(培养56 d),次生林的Q10值在饱和含水量为60%SSM时候最高,而原始林的Q10则是随着水分升高而升高。水分显著影响土壤碳矿化,但水分对不同生态系统Q10的影响方向和程度不同[27],因此水分对温度敏感性的影响也不一致,Gulledge和Schimel发现水分增加会促进土壤呼吸敏感性[35],而Dörr[36]则发现水分变化对土壤呼吸温度敏感性Q10没有产生显著影响。这些不一致的结论可能是由于研究地所处的生态系统中土壤基质不同,也可能是因为不同生态系统中水分对土壤通透性,植物根系及微生物活动的影响不一样所导致的[37]。
4 结论
植被类型变化对土壤碳矿化速率具有显著影响,原始的针阔混交林在砍伐后形成次生林,其土壤碳矿化能力显著增强。温度和水分对原始林和次生林的土壤碳矿化均具有显著影响,且二者间存在显著的交互效应。利用双因素模型R=(a×ebT)×(c×w2+d×w+e)可很好模拟长白山森林土壤碳矿化速率的变异(82.7%—95.9%)。次生林土壤碳矿化速率及其温度敏感性均高于原始林,意味着森林采伐会加速土壤有机质的分解,从而导致土壤碳贮量下降。
[1] Alvarez R,Alvarez C.Temperature regulation of soil carbon dioxide production in the Humid Pampa of Argentina:estimation of carbon fluxes under climate change.Biology and Fertility of Soils,2001,34(4):282-285.
[2] Xiao H L.Climate change in relation to soil organic matter.Soil and Environmental Sciences,1999,8(4):300-304.
[3] Wang X,Zhou G S,Jiang Y L,Jia B R,Wang F Y,Zhou L.Soil respiration in natural mixed(Betula platyphylla and Populus davidiana)secondary forest and primary broad-leaved Korean pine forest.Journal of Plant Ecology(Chinese Version),2007,31(3):348-354.
[4] Post W M,Kwon K C.Soil carbon sequestration and land-use change:processes and potential.Global Change Biology,2000,6:317-327.
[5] Vitousek P M,Mooney H A,Lubchenco J,Melillo J M.Human domination of Earth's ecosystems.Science,1997,278(5325):494-499.
[6] Kucera C L,Kirkham D R.Soil respiration studies in tallgrass prairie in Missouri.Ecology,1971,52(5):912-915.
[7] Wildung R E,Garland T R,Buschbom R L.The interdependent effects of soil temperature and water content on soil respiration rate and plant root decomposition in arid grassland soils.Soil Biology and Biochemistry,1975,7(6):373-378.
[8] Ceccon C,Panzacchi P,Scandellari F,Prandi L,Ventura M,Russo B,Millard P,Tagliavini M.Spatial and temporal effects of soil temperature and moisture and the relation to fine root density on root and soil respiration in a mature apple orchard.Plant and Soil,2011,342(1/2):195-206.
[9] Davidson E A,Belk E,Boone R D.Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest.Global Change Biology,1998,4(2):217-227.
[10] Yang Y S,Chen G S,Wang X G,Xie J S,Gao R,Li Z,Jin Z.Response of soil CO2efflux to forest conversion in subtropical zone of China.Acta Ecologica Sinica,2005,25(7):1684-1690.
[11] Wang X G,Zhu B,Wang Y Q,Zheng X H.Soil respiration and its sensitivity to temperature under different land use conditions.Acta Ecologica Sinica,2007,27(5):1960-1968.
[12] Reichstein M,Bednorz F,Broll G,Kätterer T.Temperature dependence of carbon mineralisation:conclusions from a long-term incubation of subalpine soil samples.Soil Biology and Biochemistry,2000,32(7):947-958.
[13] Reichstein M,Subke J A,Angeli A C,Tenhunen J D.Does the temperature sensitivity of decomposition of soil organic matter depend upon water content,soil horizon,or incubation time?Global Change Biology,2005,11(10):1754-1767.
[14] Fang C,Moncrieff J B.The dependence of soil CO2efflux on temperature.Soil Biology and Biochemistry,2001,33(2):155-165.
[15] Zhao S Q,Fang J Y,Zong Z J,Zhu B,Shen H H.Composition,structure and species diversity of plant communities along an altitudinal gradient on the northern slope of Mt.Changbai,Northeast China.Biodiversity Science,2004,12(1):164-173.
[16] Shen C C,Xiong J B,Zhang H Y,Feng Y Z,Lin X G,Li X Y,Liang W J,Chu H Y.Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain.Soil Biology and Biochemistry,2013,57:204-211.
[17] Dai J Z,Wei Z J,He N P,Wang R M,Wen X H,Zhang Y H,Zhao X N,Yu G R.Effect of grazing enclosure on the priming effect and temperature sensitivity of soil C mineralization in Leymus chinensis grasslands,Inner Mongolia,China.Chinese Journal of Plant Ecology,2012,36(12):1226-1236.
[18] Dube F,Zagal E,Stolpe N,Espinosa M.The influence of land-use change on the organic carbon distribution and microbial respiration in a volcanic soil of the Chilean Patagonia.Forest Ecology and Management,2009,257(8):1695-1704.
[19] Zheng D L,Chen J Q,LeMoine J M,Euskirchen E S.Influences of land-use change and edges on soil respiration in a managed forest landscape,WI,USA.Forest Ecology and Management,2005,215(1/3):169-182.
[20] Huang C C,Ge Y,Chang J.Effect of artificial disturbance on soil respiration of forest ecosystem.Journal of Zhejiang Forestry Science&Technology,1999,19(4):19-27.
[21] Trumbore S E,Davidson E A,de Camargo P B,Nepstad D C,Martinelli L A.Belowground cycling of carbon in forests and pastures of Eastern Amazonia.Global Biogeochemical Cycles,1995,9(4):515-528.
[22] Larionova A A,Yermolayev A M,Blagodatsky S A,Rozanova L N,Yevdokimov I V,Orlinsky D B.Soil respiration and carbon balance of gray forest soils as affected by land use.Biology and Fertility of Soils,1998,27(3):251-257.
[23] Lal R,Kimble J,Follett R.Land use and soil C pool in terrestrial ecosystems//Lal R,Kimbkle J,Follett R,Stewarteds B A,eds.Management of Carbon Sequestration in Soil.Boca Raton:CRC Press,1998:1-10.
[24] Liu Y,Han S J.Factors controlling soil respiration in four types of forest of Changbai Mountains,China.Ecology and Environmental Sciences,2009,18(3):1061-1065.
[25] Jia B,Zhou G,Wang Y,Wang F,Wang X.Effects of temperature and soil water-content on soil respiration of grazed and ungrazed Leymus chinensis steppes,inner Mongolia.Journal of Arid Environments,2006,67(1):60-76.
[26] Liu H S,Liu H J,Wang Z P,Xu M,Han X G,Li L H.The temperature sensitivity of soil respiration.Progress in Geography,2008,27(4):52-58.
[27] Bowden R D,Newkirk K M,Rullo G M.Carbon dioxide and methane fluxes by a forest soil under laboratory-controlled moisture and temperature conditions.Soil Biology and Biochemistry,1998,30(12):1591-1597.
[28] Merilä P,Ohtonen R.Soil microbial activity in the coastal Norway spruce[Picea abies(L.)Karst.]forests of the Gulf of Bothnia in relation to humus-layer quality,moisture and soil types.Biology and Fertility of Soils,1997,25(4):361-365.
[29] Rey A,Pegoraro E,Tedeschi V,De Parri I,Jarvis P G,Valentini R.Annual variation in soil respiration and its components in a coppice oak forest in Central Italy.Global Change Biology,2002,8(9):851-866.
[30] Qi Y,Xu M.Separating the effects of moisture and temperature on soil CO2efflux in a coniferous forest in the Sierra Nevada mountains.Plant and Soil,2001,237(1):15-23.
[31] Balser T C,Wixon D L.Investigating biological control over soil carbon temperature sensitivity.Global Change Biology,2009,15(12):2935-2949.
[32] Yang Y,Huang M,Liu H S,Liu H J.The Interrelation between temperature sensitivity and adaptability of soil respiration.Journal of Natural Resources,2011,26(10):1811-1820.
[33] Zhang J B,Song C C,Yang W Y.Temperature sensitivity of soil respiration and its effecting factors in the different land use.Acta Scientiae Circumstantiae,2005,25(11):1537-1542.
[34] Raich J W,Schlesinger W H.The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate.Tellus B,1992,44(2):81-99.
[35] Gulledge J,Schimel J P.Controls on soil carbon dioxide and methane fluxes in a variety of taiga forest stands in interior Alaska.Ecosystems,2000,3(3):269-282.
[36] Dörr H,Münnich K.Annual variation in soil respiration in selected areas of the temperate zone.Tellus B,1987,39(1/2):114-121.
[37] Chen Q S,Li L H,Han X G,Yan Z D.Effects of water content on soil respiration and the mechanisms.Acta Ecologica Sinica,2003,23(5):972-978.
参考文献:
[2] 肖辉林.气候变化与土壤有机质的关系.土壤与环境,1999,8(4):304-304.
[3] 王旭,周广胜,蒋延玲,贾丙瑞,王风玉,周莉.山杨白桦混交次生林与原始阔叶红松林土壤呼吸作用比较.植物生态学报,2007,31(3):348-354.
[10] 杨玉盛,陈光水,王小国,谢锦升,高人,李震,金钊.中国亚热带森林转换对土壤呼吸动态及通量的影响.生态学报,2005,25(7):1684-1690.
[11] 王小国,朱波,王艳强,郑循华.不同土地利用方式下土壤呼吸及其温度敏感性.生态学报,2007,27(5):1960-1968.
[15] 赵淑清,方精云,宗占江,朱彪,沈海花.长白山北坡植物群落组成、结构及物种多样性的垂直分布.生物多样性,2004,12(1):164-173.
[17] 代景忠,卫智军,何念鹏,王若梦,温学华,张云海,赵小宁,于贵瑞.封育对羊草草地土壤碳矿化激发效应和温度敏感性的影响.植物生态学报,2012,36(12):1226-1236.
[20] 黄承才,葛滢,常杰.人为扰动对森林生态系统土壤呼吸的影响.浙江林业科技,1999,19(4):19-27.
[24] 刘颖,韩士杰.长白山四种森林土壤呼吸的影响因素.生态环境学报,2009,18(3):1061-1065.
[26] 刘洪升,刘华杰,王智平,徐明,韩兴国,李凌浩.土壤呼吸的温度敏感性.地理科学进展,2008,27(4):52-58.
[32] 杨毅,黄玫,刘洪升,刘华杰.土壤呼吸的温度敏感性和适应性研究进展.自然资源学报,2011,26(10):1811-1820.
[33] 张金波,宋长春,杨文燕.不同土地利用下土壤呼吸温度敏感性差异及影响因素分析.环境科学学报,2005,25(11):1537-1542.
[37] 陈全胜,李凌浩,韩兴国,阎志丹.水分对土壤呼吸的影响及机理.生态学报,2003,23(5):972-978.
