3种短日照依赖型地被菊光周期调控研究
2013-09-19赵耀吕晋慧陈阳任磊李艳峰
赵耀,吕晋慧,陈阳,任磊,李艳峰
(山西农业大学 林学院,山西 太谷030801)
菊花是典型的短日照宿根草本花卉[1],在我国栽培历史悠久,品种资源十分丰富。其中,地被菊因其花量大、耐旱、耐盐碱和易管理而成为节日用花和园林绿化的重要材料[2],但目前国内外地被菊主栽品种大都是短日照敏感型品种,花期主要集中在秋季,少量短日照不敏感的早花品种与大多数地被菊品种存在花期不遇的问题。花期调控是保证杂交育种顺利进行的关键[3],其中,光周期调控是菊花栽培生产的主要技术[4]。有关菊花光周期调控的研究,陈晓[5]和马月萍等[6]认为,菊花对光照长度的反应作用于茎尖附近,且没有开花类物质传递的现象。李申军等[7]认为切花菊须要短日照处理21~28d可提前开花,多花型菊花则需要更长时间的短日照;刘慧琴等[8]对切花菊“神马”的研究报告指出,在短日照下,花蕾形成需要20多天,花朵露白需要50d左右;邝琦等[9]认为,从短日照至开花,切花小菊需要6~10周,大菊品种需要9~12周。有关地被菊不同短日照处理及其对不同地被菊品种开花习性的影响等未见报道。本研究就不同光周期处理对“毛白玉”、“紫娇”和“大红”成花影响进行研究,以期为其花期调控和杂交授粉提供科学依据。
1 材料与方法
1.1 材料
供试材料为短日照依赖型地被菊品种“毛白玉”、“紫娇”和“大红”,其自然始花期分别为8月下旬、9月中旬以及10月上旬。
1.2 方法
试验于2012年5月在山西农业大学苗圃内进行。取生长健壮、长约10cm的脚芽,插入沙床育苗。5月下旬,当苗长出2~3片新叶时,定植于苗床,进行常规管理。
每品种分为 A、B、C、D、E 5个光周期处理,即用黑白遮光布分别对其进行0d、20d、30d、40d和50d的遮光处理,其中A处理自然光照为对照,每处理4个重复。于6月5日开始短日照处理,遮光处理时间为每日18:00至次日8:00[4]。
植物成花过程中,芽开放后露出花序蕾,为花序出现期;当观测植株有一朵或同时有几朵花的花瓣开始完全开放,为始花期;当观测植株一半以上的花序蕾展开花瓣,为盛花期;当观测的植株上留有少数的花为末花期[10]。
记录不同地被菊品种花序出现期、始花期、盛花期和末花期时间,观察并记录株高、花色、花径、开花数量等指标。
从2012年6月5日至7月25日,于每日8:00揭棚以前分别测定棚内外气温、地表温度、5cm土层温度以及10cm土层温度。
2 结果与分析
2.1 不同短日照处理对地被菊花期的影响
由表1可见,不同短日照处理对“毛白玉”花序出现期没有影响,但短日照处理30d、40d、50d条件下,始花期、盛花期和末花期明显提前,分别提前27d、22d、25d,其中定植处理至始花期共53d。因此,“毛白玉”短日照处理30d以上即可实现提前开花。
不同短日照处理对“紫娇”花期影响明显。20 d和30d短日照处理后,花序出现期明显提前,分别比对照提前12d和43d,始花期、盛花期和末花期与对照差异不明显;短日照处理40d,花序出现期、始花期明显提前,分别提前43d和44d;短日照处理50d后花期均明显提前,花序出现期、始花期、盛花期、末花期分别比对照提前43d、44d、39 d、35d,其中定植处理至始花期共61d。因此,“紫娇”短日照处理50d可实现提前开花。
“大红”短日照处理后和“紫娇”表现相近。短日照处理20d和30d后,花序出现期明显提前,分别比对照提前5d和64d,始花期、盛花期和末花期与对照没有明显差异;短日照处理40d条件下,花序出现期、始花期明显提前,分别提前64d和51 d。短日照处理50d后花期均明显提前,花序出现期、始花期、盛花期和末花期分别比对照提前64d、51d、54d和45d,其中定植处理至始花期共74d。因此,“大红”短日照处理50d可实现提前开花。
“紫娇”和“大红”短日照处理30d提前形成的花蕾不能正常开花,短日照处理40d提前形成的花蕾中,少数可以正常开花进入始花期,大多数花蕾不能正常开花,不能正常开花的花枝从顶端或侧枝发出营养枝,转为营养生长,出现成花逆转。
2.2 短日照处理对地被菊生长发育性状的影响
由表2可见,短日照处理对“毛白玉”株高影响不显著,“紫娇”和“大红”短日照处理30d、40d和50d后株高显著高于对照。短日照处理30d、40d和50d,“毛白玉”开花数量显著低于对照,短日照处理20d,与对照差异不显著;“紫娇”和“大红”短日照处理40d和50d后开花数量显著低于对照,短日照处理20d、30d开花数量与对照差异不显著。各品种不同处理间花径差异不显著。短日照处理后提前开花的地被菊中,“毛白玉”为白色花无褪色现象,“紫娇”、“大红”花色劣变。
2.3 短日照处理期间棚内外温差变化
由表3可见,总体棚内温度略高于棚外温度,而不同土层温差不明显。
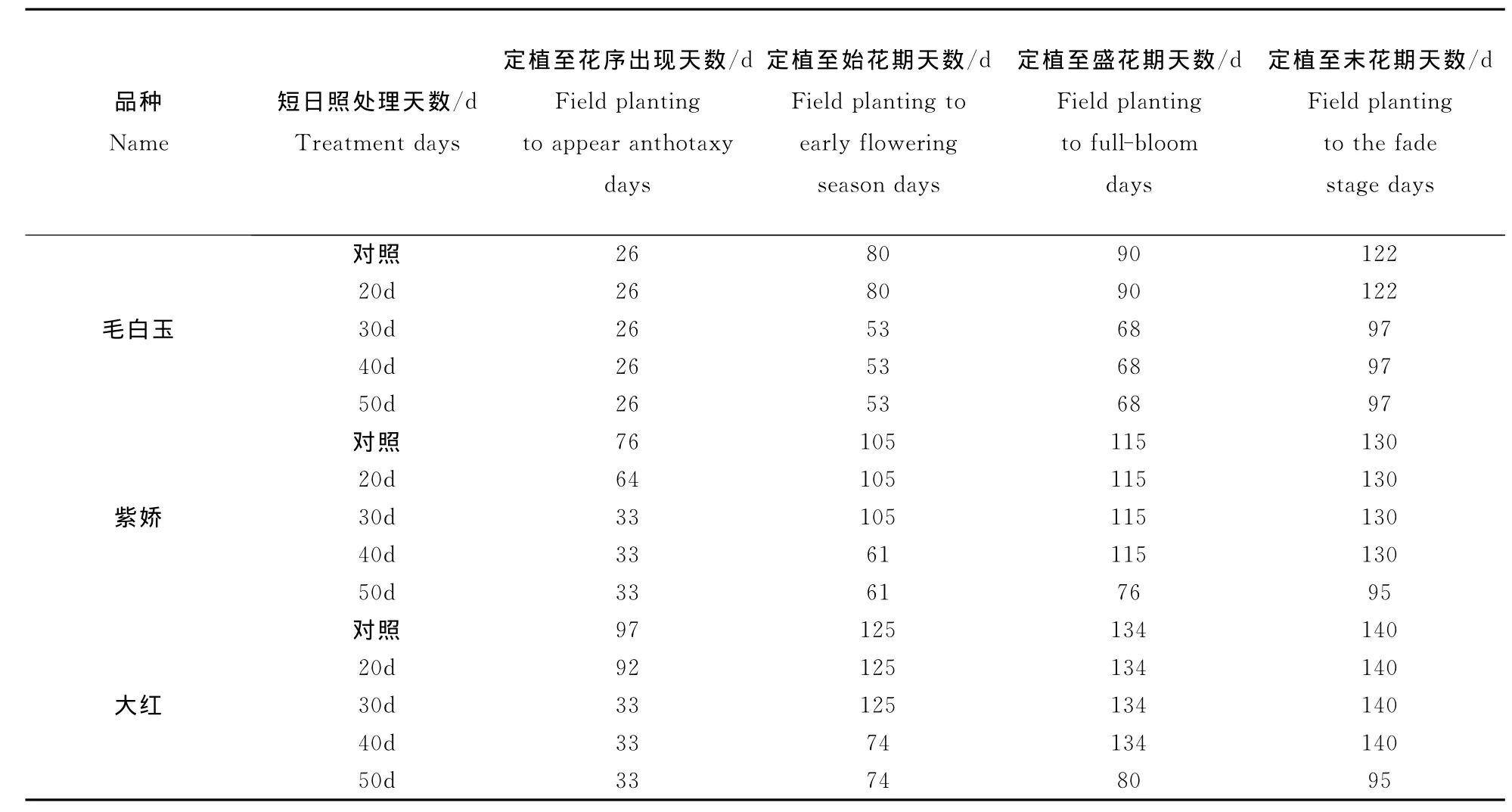
表1 不同短日照处理对地被菊花期的影响Table 1 The effect of short-day treatments to the florescence of Blackeyed Susanlight

表2 不同短日照处理后性状对比Table 2 Character comparison of different days'short-day treatments

表3 棚内外平均温度Table 3 Average temperature of inside and outside greenhouse
3 结论与讨论
3.1 不同短日照处理对地被菊花期的影响
有关菊花短日照诱导的报道,傅晓杰[11]对107个菊花品种进行短日照诱导,结果发现菊花需短日照诱导45~80d即可提前开花,薛守纪[1]对30多个菊花品种的短日照诱导研究认为,不同品种菊花诱导至开花需50~80d。本研究与报道结果基本相符,实验结果表明:“毛白玉”、“紫娇”、“大红”分别需短日照处理30d、50d、50d可提前开花,定植处理至始花期分别需53d、61d、74d。不同品种对短日照诱导花芽分化到开花所需天数不同,一方面由于菊花感受光周期要求一定的生理年龄,而生理年龄长短因品种不同有差异[12]。另一方面短日照处理后,晚花品种可能因花芽分化期间温度高而花蕾发育迟缓。因此,不同的生理特性和高温可能是品种差异的原因。
“紫娇”和“大红”短日照处理30d提前形成的花蕾全部不能正常开花,处理40d大多数花蕾不能正常开花,只有极少数花蕾正常开花。不能正常开花的花枝出现了成花逆转现象,成花逆转是指已经开始花芽分化的分生组织,在一定的条件下重新转向 叶 片 分 化 的 现 象[13]。目 前 龙 眼[14]、欧 白芥[15]、紫苏[16]、洋 狗 尾 草[17]、大豆[13]等 的 开花诱导研究中均发现成花逆转现象。成花逆转的原因与环境因素有关,引起逆转的条件同促进开花的条件相反[18],本文中引起成花逆转的环境因素是短日照中断。成花逆转与成花决定完成的程度有关,只有成花决定完成不彻底的植物在非诱导条件下可发生逆转,且花序的决定与花的决定并不是一个连续的过程[18]。
近年来,随着分子生物学的发展,人们认为植物成花和成花逆转是遗传基因和环境因素共同作用的结果。菊花属于短日照植物,对光周期信号非常敏感,PHYA-PHYB类光敏色素感受昼夜长短产生昼夜节律,随着昼夜节律的变化,CCA1、LHY等昼夜节律类基因的mRNA水平改变,CDF1和GI感受到这种变化,并把信号传递给CO[19],CO基因的表达量调节FT的表达[20],目前已在地被菊中分离出FT同源基因[21],并把拟南芥FT基因转入菊花证明其表达可促进菊花提早开花[22]。若昼夜节律发生变化,即短日照中断后,感受光周期的基因PHYA-PHYB可能表达丰度发生变化,进而影响其它成花基因的表达,导致花枝重新转向叶片分化,发生成花逆转。植物成花逆转在分子方面的报道尚不足够,这方面还需要做进一步研究。
3.2 短日照处理对地被菊生长发育性状的影响
短日照处理后植株株高显著增加,但花量显著减少。这一现象可能是由于遮荫导致植物各器官生物量分配发生变化,刘兵等[23]在不同光处理(自然光照、光富集、遮荫)下大豆干物质分配影响的研究中认为,光富集后大豆积累的光合产物更多地分配到库(消耗养料或储藏养料的器官,如花、果、种子),遮荫后大豆积累的光合产物更多分配给源(制造养料,并向其他器官提供营养物质的器官,如叶片)。可见遮荫导致了光合产物更多分配给了营养器官,向生殖器官转运的很少,因此植株株高显著增加,花量减少。
在花期调控过程中,植株花朵花色常发生变化,本文经短日照处理使3个地被菊品种花期提前,但“紫娇”和“大红”花色劣变。花青素的合成是影响花色的关键因素[24]。研究表明养分和光照充足,蔗糖含量高,对花青素的生物合成有促进作用,花青素含量越高,花朵越鲜艳。光周期处理后菊花花芽分化和花蕾孕育阶段提前至高温季节,同时,通过对短日照处理期间棚内外气温和土温的跟踪测量表明,棚内气温比棚外气温高。气温高,植株呼吸作用强,不利于碳水化合物的积累,抑制花青素的形成。另外,短日照处理缩短了光照时间,相应缩短了光合作用时间。气温高、光照不足,是造成花色劣变的主要原因。
4 结论
为实现菊花花期调控,本文对3种不同地被菊进行短日照诱导试验,得到诱导开花的最短天数,但同时发现要准确预定花期,需对菊花生长发育特性有透彻了解,掌握各品种的短日照诱导开花的天数、营养生长期等才能准确推算短日照处理起点,并且为保证其开花质量,需要在处理期间采取一定降温措施。这个过程既耗费人力物力又占用太多时间,且不可控因素如风雨虫害、人畜破坏等也会影响短日照诱导效率。因此笔者认为,通过杂交等方式培育早花期菊花新品种才是实现提前开花的根本,而短日照处理可作为培育新品种的辅助手段,确保杂交授粉高效进行。
[1]薛守纪.菊花栽培[M].北京:中国林业出版社,1981:14.
[2]王彭伟,陈俊愉.地被菊新品种选育研究[J].园艺学报,1990,17(3):223-228.
[3]吕晋慧,赵耀,王媛,等.地被菊花粉活力和贮藏性研究[J].园艺学报,2012,39(12):2483-2490.
[4]毛洪玉,顾钊宇,祝朋芳.不同光周期处理对菊花C029花芽分化及开花的影响[J].西北植物学报,2010,30(10):2074-2080.
[5]陈晓,李思远,吴连成,等.光周期影响植物花时的分子机制[J].西北植物学报,2006,26(7):1490-1499.
[6]马月萍,戴思兰.植物花芽分化机理研究进展[J].分子植物育种,2003,1(4):539-545.
[7]李申军,王红艳,孙雅娟.菊花花期调控技术[J].吉林林业科技,2011,40(2):51.
[8]刘慧琴,章心惠,王宏航,等.出口切花——日本“神马”花期调控和花梗长度控制[J].浙江农业科学,2006(6):642-643.
[9]邝琦,王文通,乔志钦,等 .21个切花菊品种的光周期与花期调控研究[J].安徽农业科学,2011,39(7):3861-3862,3866.
[10]宛敏渭,刘秀珍.中国物候观测方法[M].北京:科学出版社,1979:42-44,49-52.
[11]傅晓杰.寒冷地区秋菊短日照诱导花期及调控技术研究[D].北京:中国农业大学,2006.
[12]倪月河,汪觉先.菊花栽培与鉴赏[M].上海:上海科学技术出版社,2000:3-18.
[13]韩天富,盖钧镒,王金陵,等.大豆开花逆转现象的发现[J].作物学报,1998,24(2):168-171.
[14]王纪忠.龙眼(Dimocarpus longana Lour.)树体营养与成花逆转的关系[D].福建:福建农业大学,2004.
[15]Bagnard C.Florasion et reversion chez Sinapis alba(Ⅱ).Caracteres morphologiques de plantes porteuses de reversions[J].Can J Bot,1980,58:2335-2342.
[16]梅虎,谈锋.内源激素和核酸与紫苏花芽生理分化关系[J].西南农业大学学报,2002,24(2):118-120,150.
[17]Battey N H,Lyndon R F.Reversion of flowering[J].The BotRev,1990,56(2):162-189.
[18]张翠华.光周期诱导菊花成花及成花逆转机理研究[D].山东:山东农业大学,2009.
[19]Doyle M R,Davis S J,Bastow R M,et al.The ELF 4gene controls circadian rhythms and flowering time in Arabidopsis thaliana[J].Nature,2002,419(6902):74-77.
[20]田素波,郭春晓,郑成淑.光周期诱导植物成花的分子调控机制[J].园艺学报,2010,37(2):325-330.
[21]潘才博,张启翔,潘会堂,等.菊花FT 类似基因的克隆与表达分析[J].园艺学报,2010,37(5):769-776.
[22]姜丹,梁建丽,陈晓丽,等.拟南芥花期基因FT转化切花菊“神马”[J].园艺学报,2010,37(3):441-448.
[23]刘兵,王程,金剑,等.生殖生长期光富集和遮阴对大豆干物质分配及产量品质的影响[J].干旱地区农业研究,2009,27(2):103-107.
[24]刘建敏,刘鑫军,李晓趁.花卉花期调控常见问题分析[J].北方园艺,2008(1):139-140.
