水稻抗稻瘟病分子遗传研究进展
2013-09-19郑卓之戴良英
郑卓之,李 魏,戴良英*
(1湖南农业大学生物安全科学技术学院,长沙410128;2作物基因工程湖南省重点实验室,长沙410128)
稻瘟病是由子囊菌Magnaporthe oryzae引起的一种真菌性病害,在水稻的整个生育阶段皆可发生,是严重影响水稻生产的最主要病害之一。稻瘟病分布极广,世界上主要水稻种植区均有发生,如何防治稻瘟病早已成为世界性的重点研究课题。自上世纪中叶至今,关于稻瘟病的研究取得了许多突出成果,培育含有抗稻瘟基因的水稻品种成为防治稻瘟病的重要手段。由于稻瘟菌生理小种变异过快,各地稻瘟菌生理小种各异,水稻生长环境也差异较大,因此只含有单一抗稻瘟病基因的水稻品种很难持久、稳定地表现出稻瘟病抗性。所以,将广谱抗稻瘟病基因或者多个抗稻瘟病基因导入到一个水稻品种中,使之具有广谱而且持久的稻瘟病抗性成为水稻抗病育种研究的重要方向。
Harold H.Flor于20世纪早期提出了“基因对基因”假说[1],植物抗性基因编码的蛋白能与病原物无毒基因所编码的蛋白相互作用,进而引起植物的抗病反应。研究表明,水稻抗性基因与稻瘟病无毒基因的蛋白特异性相互作用符合“基因对基因”假说,所以对稻瘟菌无毒基因与水稻抗稻瘟菌基因的相互作用的研究,可以为水稻抗病育种中抗病基因的合理选择提供研究基础。同时,对于不同地区、不同环境选择何种水稻抗稻瘟病品种提供理论依据。
1 水稻抗稻瘟病基因的定位和克隆
迄今为止,已报道了水稻中至少85个主效抗稻瘟病基因,其中已克隆基因22个(表1)。这些主效抗稻瘟病基因成簇分布在水稻基因组除3号染色体以外的其他11条染色体上。其中以6号、11号和12号染色体分布的数量最多,分别为15、22和16个(http://www.ricedata.cn/gene/gene_pi.htm,2012-06-20)。Monosi等通过分析水稻基因组,发现水稻中约有500个基因含NBS-LRR结构,其中位于11号染色体的约占25%[2]。
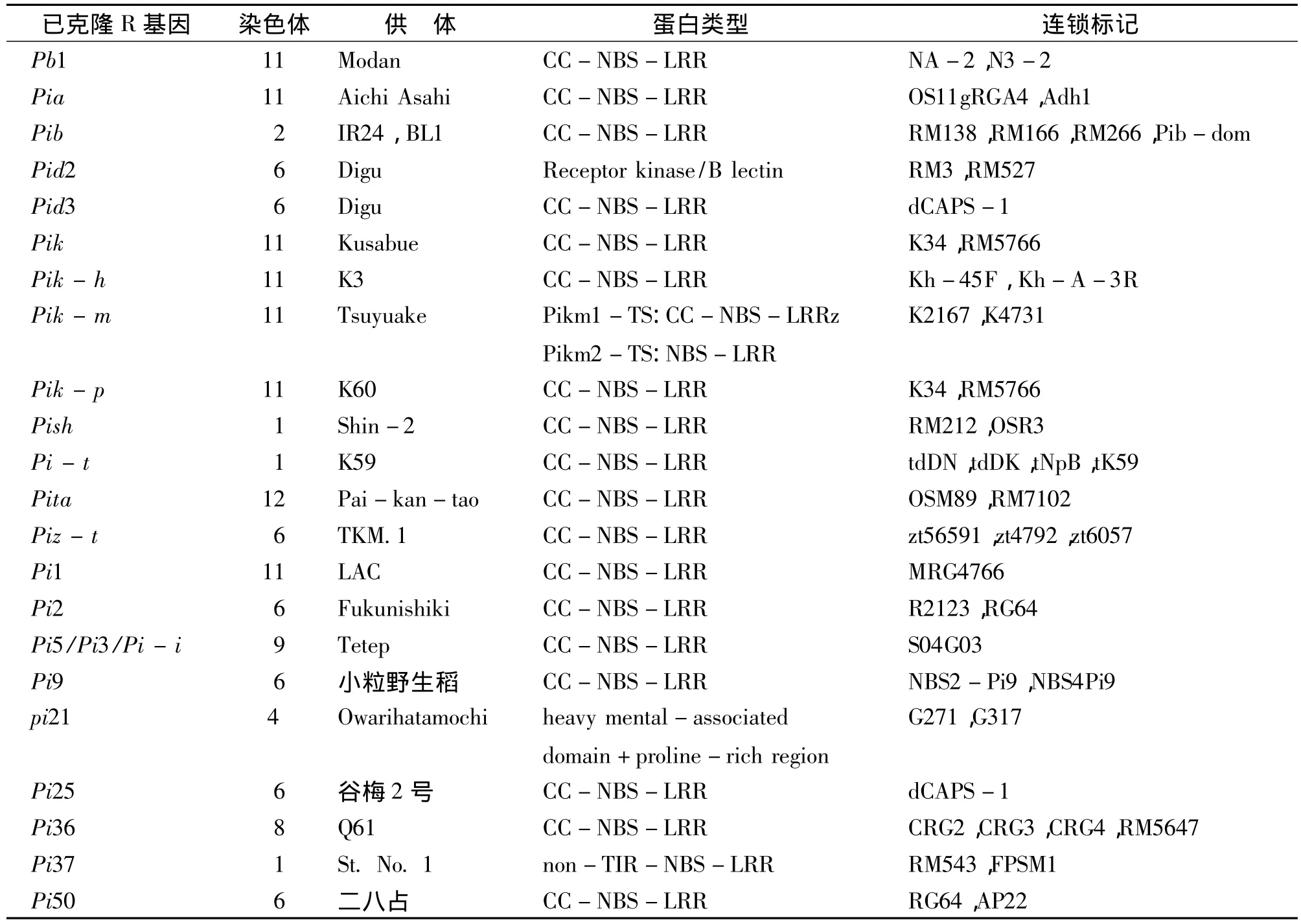
表1 已克隆的水稻抗稻瘟基因分子定位及结构特点Table 1 Cloned rice blast resistance genes molecular localization and structure characteristics
2 水稻抗稻瘟病基因的结构特点
通过对比所有植物中已克隆的70多个抗性基因,在一些结构域上它们表现出高度保守性和同源性。这些保守的功能性结构域包括:核苷酸结合位点(NBS结构域)、富含亮氨酸重复(LRR结构域)、Toll蛋白和白细胞介素1受体(TIR结构域)、无规则卷曲(CC结构域)、跨膜结构域(TM)、激酶(KIN)、亮氨酸拉链(LZ)等。植物抗性基因根据这些保守结构域大致可分为4类[3]:NBS-LRR类型、KIN类型、TM-LRR类型和TM-CC类型。也有少数已克隆的植物抗性基因不属于以上4类,如第一个克隆的植物抗性基因Hm1,它编码一个HC-毒素还原酶[4]。
NBS-LRR结构域中,N端NBS结构是最为保守的部分,能与核苷酸结合并参与调控细胞死亡,从而使植物表现出抗病反应[5]。C端的LRR结构域是一段多亮氨酸重复出现的结构。LRR结构域能识别病原物无毒基因产物,引发抗性反应的产生[6],是抗病专化性的基础[7]。例如,在 Pita/Avr-Pita的直接互作中,Pita表达产物与Avr-Pita表达产物的特异性结合与LRR区域有关[8]。植物在长时间与病原物共同进化后使得不同基因中的LRR区域具有明显的差异。NBS-LRR类型抗性基因在已克隆的70多个植物抗性基因中是最多的一种,占70%左右[9]。在已克隆的22个水稻抗稻瘟病基因中,除Pi21含重金属结合域(heavy mental-associated domain)和富含脯氨酸结构域(proline-rich region)[10],Pid -2 编码凝集素受体激酶(Receptor kinase/B lectin)外[11],其他抗稻瘟病基因均属于NBS-LRR类型(表1)。
3 稻瘟病Avr基因的鉴定和克隆
病原物Avr基因编码的蛋白能直接或间接地被寄主植物体内相应的R蛋白特异性识别并激活植物的抗病反应。近年来,对于真菌Avr基因的鉴定取得了较大的进展,为研究R基因与Avr基因的相互作用、共同进化以及植物抗病机制提供了理论基础。
至今已有40多个稻瘟病Avr基因被鉴定,其中9 个已被克隆:Pwl1、Pwl2、Avr - CO39、Avr- Pita、ACE1、AvrPiz - t、Avr - Pia、Avr - Pii和 Avr -Pik/km/kp(表2)。已经成对克隆出Avr基因和R基因的有:Avr-Pita与Pita,AvrPiz-t与Piz-t,Avr-Pia与Pia,以及Avr-Pik/km/kp与Pik/Pikm/Pik-p(表2)。此外,通过序列比对和遗传关联分析,鉴定了AvrPia、AvrPii和AvrPik/km/kp 3个无毒基因[12,13]。其中,AvrPii所编码的蛋白包含典型的锌指结构和 LXAR结构[13],前者是一种通过与DNA结合进而调控基因表达的保守结构域。另外一个已克隆的Avr基因ACE1编码一个含4 035个氨基酸的蛋白,该蛋白是已克隆的9个稻瘟病无毒基因中唯一一个非分泌蛋白(表2)。ACE1的N-端含有I型聚酮合酶(polyketide synthase,PKS),C-端则含与细菌非核糖体多肽合成酶(non-ribosomal peptide synthetases,NRPS)同源的特殊无毒结构[14]。该蛋白是一种聚酮化合物的生物合成中的关键调控因子[14]。有趣的是,ACE1只在成功侵染水稻叶片的稻瘟病成熟附着胞中表达。ACE1与其在水稻中对应的R基因Pi33并不是直接相互作用,而是ACE1在附着胞侵染过程中引发了Pi33介导的抗病反应[14]。Li等成功地从稻瘟病无毒菌株81278ZB15中分离到了Avr基因AvrPiz-t[15]。通过生物信息学手段分析表明,AvrPiz-t编码长度为108个氨基酸的分泌蛋白,但在其他真菌中却找不到同源的蛋白。测序分析发现毒性亲本Guy11菌株中的AvrPiz-t起始密码子上游462 bp处有一个Pot3转座子插入导致AvrPiz-t无毒功能的丧失。启动子功能验证实验表明,Pot3在位于起始密码子上游537 bp和736 bp处插入并不影响该菌株的无毒性,因而只有保证起始密码子上游大于462 bp的区域工作正常时,AvrPiz-t才能够发挥其无毒基因的功能[15]。AvrPiz-t表达产物在烟草叶片中能够抑制BAX诱导的细胞程序性死亡,证明了AvrPiz-t对稻瘟病的侵染致病有一定的作用[15]。
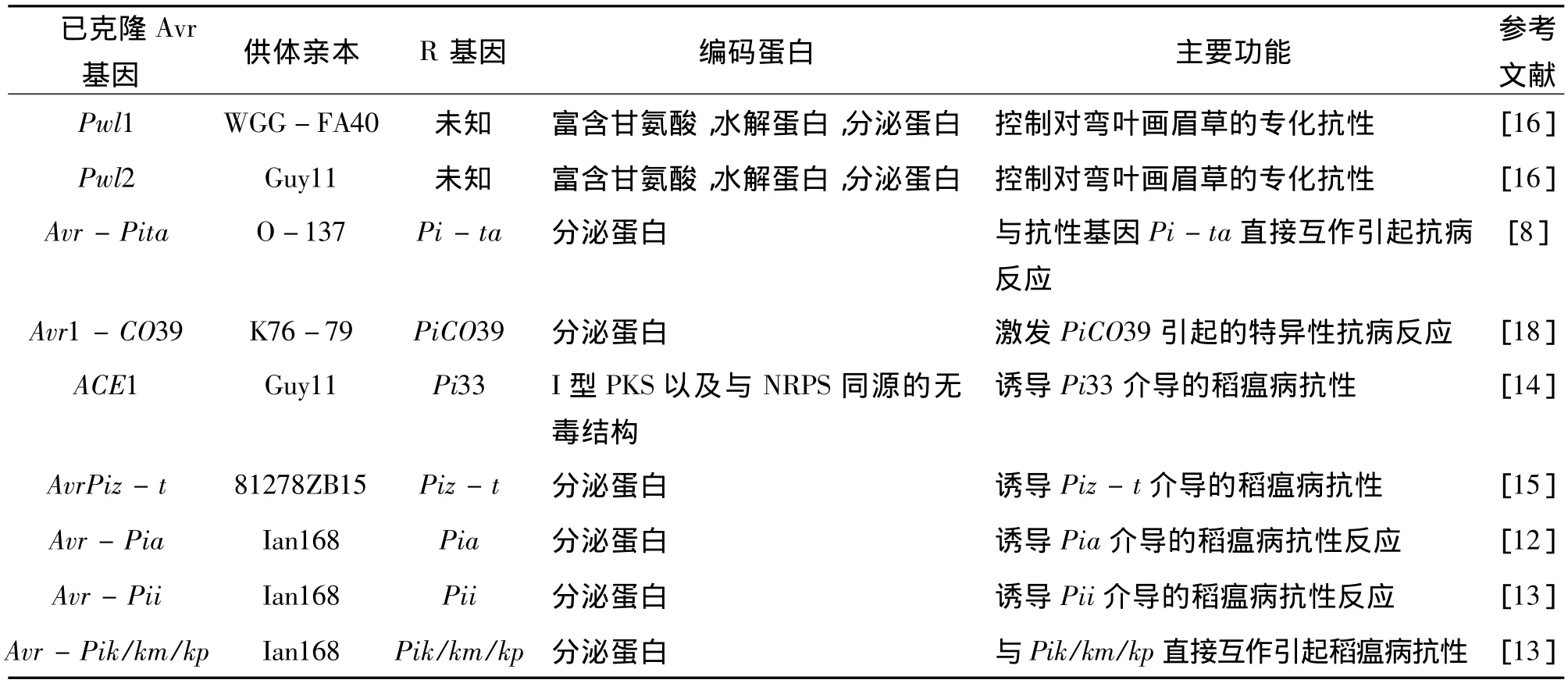
表2 已克隆的稻瘟菌无毒基因Table 2 Cloned Magnaporthe oryzae avirulence genes
4 水稻抗稻瘟病基因与稻瘟病Avr基因的互作机制
目前研究发现,大多数植物R基因与病原菌Avr基因所编码的蛋白并不能直接互作而是R蛋白通过寄主体内的辅助蛋白与Avr蛋白间接互作,即“保卫”模式(Guard Model)。例如病原细菌Pseudomonas syringae分泌的效应蛋白 AvrRpm1、AvrB和AvrRpt2只有在磷酸化或水解辅助蛋白RIN4后才能引起相应R蛋白RPM1和RPS2介导的抗病反应。在已定位和克隆的稻瘟病抗性基因中存在一些广谱抗性基因,如Pi1、Pi2、Pi-gm和Pihk1等。这种无毒蛋白对辅助蛋白RIN4修饰后的间接作用模式,可用以解释一种抗性蛋白识别多种无毒蛋白出现广谱抗性的原因。
除了间接作用外,植物R蛋白与病原菌Avr蛋白还有一种方式是直接相互作用。例如,水稻中抗稻瘟病R蛋白Pita与稻瘟病Avr蛋白Avr-Pita,Pik与Avr-Pik都属于直接作用。Avr-Pita C末端具的176个氨基酸(Avr-Pita176)包含一个金属蛋白酶功能域。Avr-Pita176能与Pita的LRR区域特异性结合,引发植物的抗病反应[8]。Pi-ta与其同源感病基因编码产物的区别仅仅是在第918个氨基酸由丙氨酸置换成丝氨酸[19]。Avr-Pita176或者Pita的LRR区域中任意一个氨基酸的改变都会破坏Avr-Pita与Pita的相互作用,从而使植物失去对稻瘟病的抗性[8]。Pik及其等位基因中都含有2个NBS-LRR基因,只有在2个NBS-LRR基因都存在的情况下才表现出抗病反应[20]。不同的Pik等位基因与对应的不同Avr-Pik基因的表达产物能够特异性结合,它们的结合方式与Pita不同,不是与LRR结构域有关而是与Pik-1的N-端的CC结构域相关。此外,需要2个NBS-LRR基因才能介导对同一个稻瘟病菌株抗性的还有 Pi5和Pia[22-23]。
对于稻瘟病菌来说最简单地避免被水稻R基因识别的方法就是失去Avr基因。Avr-Pia和Avr-Pii似乎采取的就是这种方式避免被水稻R基因识别,因为在对已克隆的Avr基因的DNA多态性分析中,Avr-Pia和Avr-Pii仅表现出存在缺失性多态[21]。然而在稻瘟病菌与水稻的协同进化过程中,带有Avr-Pik的稻瘟病菌株并不是通过失去Avr-Pik避免R基因的识别,而是通过氨基酸的置换形成不同的等位基因。那么有可能Avr-Pik的缺失比Avr-Pia和Avr-Pii的缺失对稻瘟菌而言有着更大的影响。要明白Avr基因在稻瘟病侵染过程中起到什么作用,就需要在其对应R基因缺失的情况下分析确认其与寄主植物的哪一个或几个靶标蛋白起作用。
5 展望
植物不像动物一样具有完善的获得性免疫系统,所以对于病原物的抗性只能通过抗性基因的表达来完成。虽然对于水稻抗性基因的研究已经取得了较大的进展,但是由于稻瘟病菌致病小种的易变性和多态性,水稻生产依然受到较大威胁。定位、克隆更多广谱、稳定的稻瘟病R基因对于水稻抗病育种具有重要意义。但是从长远来看,由于稻瘟病菌极强的适应能力和较快的小种变异速度,水稻R基因很难保持稳定、持续的抗性。所以,有必要对水稻R基因与稻瘟病菌Avr基因的互作机制进行更深入的研究,理解R基因与Avr基因的识别方式、水稻内在抗性机制以及水稻R基因与稻瘟病菌Avr基因的共同进化规律,找出能广谱、持续、稳定提高水稻对稻瘟病抗性的方法。
[1] Flor HH.Complementary genetic systems in flax and flax of recombination values in heredity [J].Adv Genet,1956,8:29-54.
[2] Monosi B,Wisser RJ,Pennill L,et al.Full-genome analysis of resistance gene homologues in rice[J].Theoretical and Applied Genetics,2004,109:1434-1447.
[3] Martin GB,Bogdanove AJ,Sessa G.Understanding the functions of plant disease resistance proteins[J].Annu Rev Plant Bio,2003,154:23-61.
[4] Johal GS,Briggs SP.Reduetase activity encoded by the HM1 disease resistance gene in maize [J].Science,1992,258:985-987.
[5] Traut TW.The function and consensus motifs of nine types of peptide segments that form different types of nucleiotide - binding sites[J].Eur J Biochem,1994,222:9-19.
[6] Ronald PC.The molecular basis of disease resistance in rice[J].Planr Mol Biol,1997,35:179 -186.
[7] Hulbert SH,Webb CA,Smith SM,et al.Resistance gene complexes:evolution and utilization[J].Annu Rev Phyropatho,2001,139:285-312.
[8] Jia Y,McAdams SA,Bryan GT,et al.Direct interaction of resistance gene and avirulence gene products confers rice blast resistance[J].EMBO J,2000,19:4004 -4014.
[9] Liu Jinling,Liu Xionglun,Dai Liangying,et al.Recent progress in elucidating the structure.Function and evolution of disease resistance genes in plant[J].Journal of Genetics and Genomics,2007,34:765-776.
[10] Fukuoka S,Saka N,Koga H,et al.Loss of function of a proline-containing protein confers durable disease resistance in rice[J].Science,2009,325:998 -1001.
[11] Chen XW,Shang JJ,Chen DX,et al.A B-lectin receptor kinase gene conferring rice blast resistance[J].The Plant Journal,2006,46(5):794 -804.
[12] Miki S,Matsui K,Kito H,et al.Molecular cloning and characterization of the AVR-Pia locus from a Japanese field isolate of Magnaporthe oryzae [J]. Mol Plant Pathol,2009,10:361 -374.
[13] Yoshida K,Saitoh H,Fujisawa S,et al.Association genetics reveals three novel avirulence genes from the rice blast fungal pathogen Magnaporthe oryzae [J].Plant Cell,2009,21:1573 -1591.
[14] Bhnert HU,Fudal I,Dioh W,et al.A putative polyketide synthase/peptide synthetase from Magnaporthe grisea signals pathogen attack to resistant rice[J].Plant Cell,2004,16:2499 -2513.
[15] Li W,Wang B,Wu J,et al.The Magnaporthe oryzae avirulence gene AvrPiz-t encodes a predicted secreted protein that triggers the immunity in rice mediated by the blast resistance gene Piz- t[J].Mol Plant- Microbe Interact,2009,22:411 -420.
[16] Sweigard JA,Carroll AM,Kang S,et al.Identification,cloning,and characterization of PWL2,a gene for host species specificity in the rice blast fungus[J].Plant Cell,1995,7:1221-1233.
[17] 张红生,吴云雨,鲍永美,等.水稻与稻瘟病菌互作机制研究进展[J].南京农业大学学报,2012,35:1-8.
[18]Leong SA.The ins and outs of host recognition of Magnaporthe oryzae[C].Genomics of Disease,2008:199 -216.
[19] Bryan GT,Wu KS,Farrall L,et al.A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance gene Pi - ta [J].Plant Cell,2000,12:2033-2046.
[20] Ashikawa I,Hayashi N,Abe F,et al.Characterization of the rice blast resistance gene Pik cloned from Kanto51[J].Mol Breeding,2012,30:485 -494.
[21] Kanzaki H,Yoshida K,Saitoh H,et al.Arms race coevolution of Magnaporthe oryzae AVR-Pik and rice Pik genes driven by their physical interactions[J].The Plant Journal,2012,72:894 -907.
[22] Lee SK,Song MY,Seo YS,et al.Rice Pi5-mediated resistance to Magnaporthe oryzae requires the presence of two coiled-coil-nucleotide-binding-leucine-rich repeat genes[J].Genetics,2009,181:1627 -1638.
[23] Okuyama Y,Kanzaki H,Abe A,et al.A multifaceted genomics approach allows the isolation of the rice Piablast resistance gene consisting of two adjacent NBSLRR protein genes[J].Plant J,2011,66:467 -479.
