重金属Cd2+对日本楤木营养品质和抗氧化酶活性的影响1)
2013-09-18李德生李海茹茹丽叶
孟 丽 李德生 李海茹 茹丽叶 王 璠
(天津理工大学,天津,300384)
吴 强
(天津市宁河县林业局)
Cd2+是剧毒重金属元素之一,极易在水体、土壤、动植物体内迁移和富集,具有一定的致癌性[1]。目前,有关Cd2+对土壤系统的研究已有很多,重金属Cd2+通过植物吸收迁移被运输到地上部分,对植物的生长造成一定伤害。研究表明,重金属Cd2+大量积累于植物体内导致蛋白质失活,影响植物的正常代谢[2]。在重金属 Cd2+胁迫下,龙葵(Solanum nigrum L.)叶片中苹果酸、酒石酸等有机酸的质量分数变化显著,且各个生长期变化规律不一致[3]。此外,植物受到重金属Cd2+胁迫时,会产生大量活性氧自由基,为适应环境其体内形成了特定的抗氧化性酶系统,通过合成POD、SOD和CAT等酶来清除体内的活性氧自由基,使植物免受伤害。
近年来,有关重金属Cd2+对蔬菜和大田作物等草本植物影响的研究非常多[4-7],但有关 Cd2+对木本蔬菜代谢和抗性研究较少。日本楤木(Arlia elata var.inermis)属森林、木本蔬菜类,具有很高的营养价值和药用价值,在日本有“天下第一珍”的美誉。文中以1年生日本楤木为材料,探究不同质量分数重金属Cd2+处理下日本楤木叶片中可溶性蛋白、可溶性糖、Vc、叶绿素等品质指标的变化,总结重金属Cd2+污染对日本楤木抗氧化酶活性的影响规律,以期为日本楤木的栽培和生产提供一定的理论依据。
1 材料与方法
试验于2011年10月份至2012年9月份在天津理工大学气候室进行,选用长势一致的1年生日本楤木苗木为供试材料。土壤采自理工大学,W(校园土)∶W(营养土)=25∶1,混匀过2 mm筛,装入30 cm(径)×20 cm(高)塑料盆中,每盆装土 3.5 kg。供试土壤基本理化性质为:有机质、全氮、全磷、总钾质量分数分别为53.07、8.46、0.96、0.71 g·kg-1,土壤pH值为7.57。
苗木移植1周后进行重金属胁迫处理,重金属Cd2+以CdCl2·2.5H2O配成的水溶液形式施入土壤,质量分数分别为 0、1、2、5、10、20 mg·kg-1,分别记为 CK、Cd-1、Cd-2、Cd-5、Cd-10、Cd-20,每个处理设置3组重复,采用3次测定结果的平均值进行分析。
土壤理化性质的测定参照《土壤学实验指导》的方法[8];可溶性蛋白质量分数测定采用考马斯亮蓝 G-250 法[9];Vc质量分数的测定采用 2,6-二氯酚靛酚滴定法[10];可溶性糖测定利用蒽酮比色法[11];POD和CAT的活性测定参照《植物生理学实验技术》的方法[12]。
该疾病使用抗生素治疗取得了理想的治疗效果,常用药物包括四环素、土霉素等,其中这2种药物作用于病猪时需要按照猪每千克体重,不少于30 mg、控制在40 mg的标准进行治疗,每天治疗次数为2次,方式为肌肉注射,持续治疗1周;还可以使用卡那霉素药物,依照猪体重进行每天2次的20 mg药物注射治疗,治疗时间不超过5 d。
2 结果与分析
2.1 Cd2+污染对日本楤木叶片叶绿素质量分数的影响
Vc是植物体内重要的小分子抗氧化物质,能够直接清除因氧代谢、光合作用及环境胁迫等产生的活性氧[20],使植物免受伤害。在Cd2+处理情况下,日本楤木叶片内Vc质量分数呈下降趋势,Vc质量分数的变化范围为0.99 ~1.45 mg·g-1。当处理质量分数为20 mg·kg-1时,日本楤木Vc的质量分数比对照降低了31.7%,说明重金属Cd2+对Vc的合成有一定的阻碍作用,但尚未达到显著性水平(P>0.05)。
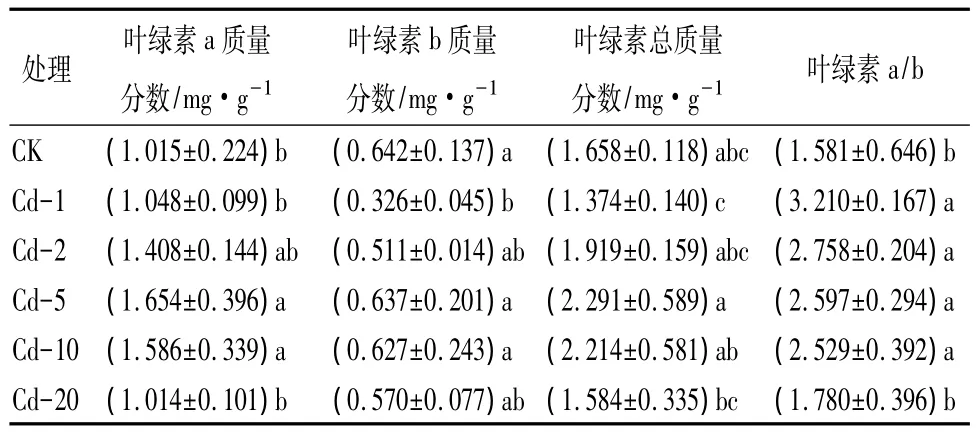
表1 Cd2+污染对日本楤木叶片叶绿素质量分数的影响
2.2 Cd2+污染对日本楤木营养品质的影响
2.2.3 Cd2+污染对日本楤木叶片Vc质量分数的影响
如表2所示,可溶性蛋白与植物体内细胞渗透势的调节有关,蛋白质质量分数的高低可间接反映作物进行各种代谢活动的强弱[16]。如表2所示,随着Cd2+处理质量分数的增加,可溶性蛋白质量分数呈现先上升后缓慢下降的趋势,与对照相比变化显著(P<0.05)。当 Cd2+质量分数为1 mg·kg-1时,可溶性蛋白质量分数达到最大值,比对照增加3.9%,可能是由于低质量分数Cd2+能够迅速刺激蛋白质合成,产生新的蛋白质,蛋白质总量增加;当Cd2+质量分数超过2 mg·kg-1时,重金属离子可能与-SH基结合导致代谢酶失活,可溶性蛋白合成受到抑制。
虚与实。园林无水不活,中国传统绘画中更是注重虚实的对比。没有水则没有空间感,在余荫山房中水景占据着重要的地位,建筑是实的,而水则是柔和的,一刚一柔拉开了实景间的距离。《道德经》在“有之以为利,无以为用”的虚实关系的阐释中透射出老子对于空间哲理的思考。在余荫山房中,粉墙为实、廊桥为虚;建筑为实,水庭为虚;廊柱栏杆为实、镂空部分为虚,在以实写虚的对比中产生“留白”空间。
2.2.2 Cd2+污染对日本楤木叶片可溶性糖质量分数的影响
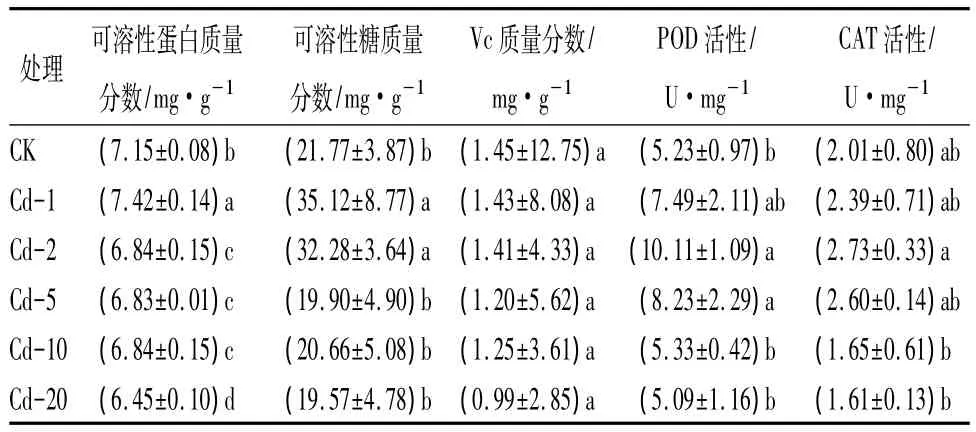
表2 Cd2+污染对日本楤木叶片品质及酶活性的影响
此次《通知》专门对高品位步行街进行了“画像”:区位优越,商业资源丰富;环境优美,公共设施健全;功能完善,名品名店集聚;特色鲜明,文化底蕴深厚;消费吸引力和辐射带动力明显,在国内国外享有较高知名度和美誉度。据了解,商务部将组织制定高品位步行街评价指标,对重点培育步行街进行跟踪指导、定期评价。
可溶性糖是植物体内碳素营养状况及农产品品质性状的重要指标之一[17],当植物受到重金属胁迫时,可溶性糖能够调节体内渗透压,降低植物伤害。重金属Cd2+通过影响植物体内碳水化合物的代谢过程,进而影响可溶性糖的质量分数[18]。表2中数据表明,当Cd2+处理质量分数低于5 mg·kg-1时,可溶性糖质量分数高于对照组,当质量分数达到1 mg·kg-1时,可溶性糖质量分数达到最大值,与对照相比显著增加61.3%(P<0.05),可能是由于大分子的多糖分解转化为单糖,也有可能是光合作用的产物直接转化为低分子的蔗糖,而不是淀粉[19]。随后,在高质量分数(Cd2+≥10 mg·kg-1)处理下,日本楤木可溶性糖质量分数下降,可能是由于高质量分数的Cd2+抑制了可溶性糖合成的酶的活性,导致植物碳水化合物代谢活动受阻,可溶性糖质量分数降低。
试验原始数据的处理和制图采用Excel软件,处理组与对照之间的差异显著性检验分析均由SPSS统计软件完成。
2.2.1 Cd2+污染对日本楤木叶片可溶性蛋白质量分数的影响
叶绿素是植物光合作用的重要组成成分,其质量分数高低与作物的产量和品质具有一定相关性[13],也是反映植物叶片光合作用强弱的重要指标之一,能够表征逆境胁迫下植物组织、器官的衰老状况[14]。从表1可以看出,在 Cd2+处理情况下,随着Cd2+处理质量分数增加,叶绿素a质量分数呈现先升高后降低的趋势,当质量分数达到5 mg·kg-1时,叶绿素a质量分数与对照相比显著增加62.9%(P<0.05);叶绿素b整体呈现下降的趋势,但未达到显著性水平(P>0.05)。在高质量分数的Cd2+胁迫下,叶绿素总质量分数、叶绿素a和叶绿素b质量分数均低于对照,原因可能是细胞内的Cd2+与叶绿素生成途径的酶的肽链中富含-SH的部分结合,改变了酶的正常构型,抑制了酶的活性,阻碍了叶绿素的合成[15];或者是重金属离子提高了叶绿素酶的活性,使得叶绿素降解加快。
模袋布块与块之间搭接时,须把前一块布的搭接反滤布平铺好后再铺设下一块模袋布,采用8号铁丝做成U形环将模袋下边沿与反滤布连接,插入土中,上边沿(模袋布表面)采用手工缝制对接,连缝平均不大于2 cm,并保证块与块之间的密实度达到设计厚度的50%以上。依次将缝制对接好的模袋布拉入水中定位。
2.3 Cd2+污染对日本楤木叶片抗氧化酶活性的影响
抗氧化酶是植物体活性氧清除系统的重要组成成分,包括POD、CAT和SOD等,这些酶能够促进活性氧自由基的清除,使细胞免受伤害[21]。表2反映了重金属Cd2+胁迫下POD和CAT的活性变化,在Cd2+胁迫时,日本楤木叶片内POD和CAT活性随处理质量分数增高呈现先增加后下降趋势;当Cd2+质量分数为2 mg·kg-1时,POD与CAT活性均达到最大值,POD活性与对照相比显著增加(P<0.05),CAT活性变化不显著(P>0.05),说明日本楤木为保护细胞免受氧化胁迫的伤害而提高POD的活性,维持自身正常代谢;当处理质量分数为20 mg·kg-1时,POD和CAT活性均低于对照,可能由于高质量分数Cd2+胁迫超过了POD和CAT的承受极限,破坏了它们的功能,抗氧化酶活性丧失。或者是由于重金属Cd2+在植物体内累积,可能会取代重金属蛋白中的必需元素(Ca2+、Mg2+、Fe2+、Zn2+),导致生物大分子构象改变,造成酶的活性下降。
再者,学校应积极举办体育比赛、体育项目表演、校园体育文化节等活动,增强校园体育文化氛围,丰富学生的课外活动,使学生劳逸结合,在学习的同时释放压力、振奋精神。
3 结束语
土壤中Cd2+污染对日本楤木叶片的营养物质有一定影响,主要表现为对可溶性蛋白、可溶性糖、Vc、叶绿素质量分数的影响。可溶性蛋白、可溶性糖和叶绿素总质量分数随Cd2+质量分数升高呈现先增加后下降的趋势,分别在Cd2+质量分数为1 mg·kg-1和5 mg·kg-1时达到最大值,表明日本楤木对低质量分数的Cd2+有一定的抵抗和适应能力。随着Cd2+质量分数的增加,日本楤木叶片内Vc合成受阻,Vc质量分数呈下降趋势。因此,在Cd2+质量分数较高的土壤中种植日本楤木会抑制其生长和营养品质。
在重金属胁迫下,植物体内活性氧增加,一旦超过抗氧化系统的防御能力,就可能对植物产生伤害。抗氧化酶(POD、SOD、CAT)是植物抗氧化系统的重要组成成分,POD和CAT能够在逆境或衰老初期表达时,清除过氧化体中产生的H2O2,起到保护植物的作用[22]。在本试验中,POD和 CAT活性随着Cd2+处理质量分数的增大呈现先增高后降低的趋势,当质量分数达到2、5 mg·kg-1时,POD 活性显著高于对照,说明日本楤木为保护细胞免受氧化胁迫的伤害而提高POD活性。
[1]Nawrot T,Plusquin M,Hogervorst J,et al.Environmental exposure to cadmium and risk of cancer:A prospective population based study[J].The Lancet Oncology,2006,7(2):119-126.
[2]Stohs S J,Bagchi D,Hassoun E,et al.Oxidative mechanisms in the toxicity of chromium and cadmium ions[J].Journal of Environmental Pathology,Toxicology and Oncology,2000,19(3):201-213.
[3]孙瑞莲,周启星,王新.镉超积累植物龙葵叶片中镉的积累与有机酸含量的关系[J].环境科学,2006,27(4):765-769.
[4]郭智,黄苏珍,原海燕.Cd胁迫对马蔺和鸢尾幼苗生长、Cd积累及微量元素吸收的影响[J].生态环境,2008,17(2):651-656.
[5]方晓航,曾晓雯,于方明,等.Cd胁迫对白菜生理特征及元素吸收的影响研究[J].农业环境科学学报,2006,25(1):25-29.
[6]陈瑛,李廷强,杨肖娥,等.不同品种小白菜对镉的吸收积累差异[J].应用生态学报,2009,20(3):736-740.
[7]罗红艳,张银龙.Cd、Pb及其相互作用对三种木本植物幼树生理生化特性的影响[J].化工之友,2006(8):49-50.
[8]林大仪.土壤学实验指导[M].北京:中国林业出版社,2004.
[9]江行玉,赵可夫.植物重金属伤害及其抗性机理[J].应用与环境生物学报,2001,7(1):92-99.
[10]郝建军.植物生理学实验技术[M].北京:化学工业出版社,2007:183-185.
[11]赵海泉.基础生物学实验指导[M].北京:中国农业大学出版社,2008.
[12]高俊凤.植物生理学实验技术[M].西安:世界图书出版公司,2000:192-199.
[13]王建湘,周杰良.不同有机肥种类对小白菜品质及产量的影响[J].上海蔬菜,2007,1(1):63-64.
[14]Jennifer C,Nora G.Cleaning up contaminants with plants[J].Biocycle,2005,46(S):30-32.
[15]Somaehekaraiah B V,Padmaja K,Prasad A R K.Phytotoxicity of cadmium ions on germination seedling of mung bean(Phaseolus vulgarize):involvement of lipid peroxides in chlorophyll degradation[J].Physiology Plant,1992,85(1):85-89.
[16]张雅倩,林琪,张玉,等.干旱胁迫对不同小麦品种花后旗叶生理特性的影响[J].干旱地区农业研究,2010,28(6):158-164.
[17]谢建治,张书廷,刘树庆,等.潮褐土重金属Cd污染对小白菜营养品质指标的影响[J].农业环境科学学报,2004,23(4):678-682.
[18]熊愈辉,杨肖娥.镉对植物毒害与植物耐镉机理研究进展[J].安徽农业科学,2006,34(13):2969-2971.
[19]邱海杰,熊治廷,费利西泰.水杨酸对Cd胁迫下两种大白菜主要营养品质的影响[J].农业环境科学学报,2006,25(4):871-874.
[20]Padh H.Cellular functions of ascorbic acid[J].Biochemistry and Cell Biology,1990,68(10):1166-1173.
[21]宇克莉,邹婧,邹金华.镉胁迫对玉米幼苗抗氧化酶系统及矿质元素吸收的影响[J].农业环境科学学报,2010,29(6):1050-1056.
[22]尹永强,胡建斌,邓明军.植物叶片抗氧化系统及其对逆境胁迫的响应研究进展[J].中国农学通报,2007,23(1):105-110.
