高寒退化草地狼毒与赖草种群空间格局及竞争关系
2013-09-11赵成章
任 珩,赵成章
(西北师范大学地理与环境科学学院甘肃省湿地资源保护与产业发展工程研究中心,兰州 730070)
空间格局是植物种群的基本特征,研究植物种群的空间格局有助于认识它们的生态过程以及与生境的相互关系[1]。其中,判定种群的空间分布类型和空间关联性是空间格局研究的两个主要内容[2],二者不仅反映植物的空间分布特点、利用环境资源的状况和种群在群落中的作用与地位,而且可以揭示植物个体之间相互依存和相互制约的复杂关系[2-4],通常被认为是研究物种间相互作用以及植物与环境间关系的一种重要手段[2,5]。竞争是自然群落中普遍存在的现象,是塑造植物形态、生活史以及植物群落结构和动态特征的主要动力之一[6]。在过度放牧和气候变迁影响下,天然草地严重退化,植物群落优势种群发生更替,改变了系统物质与能量的收支平衡和群落资源的分配格局[7],对于植物群落环境和物种的竞争能力产生影响,导致植物种群或群落形成不同的空间格局[8-9]。因此研究植物种群空间格局及竞争关系,对于揭示草地生态系统的退化机理,制定草地生态系统恢复措施具有重要意义。
在祁连山北坡阿尔泰针茅草原,狼毒(Stellera chamaejasme)和赖草(Aneurolepidium dasystachys)是原生地带性植被群落的常见种和零星分布物种[10-11]。过度放牧影响下,建群种阿尔泰针茅(Stipa krylovii)更新能力受到限制,逐渐丧失了群落中的优势地位[12],群落环境和资源利用状态随之发生改变。与此同时,狼毒凭借对干旱环境和放牧干扰的适应能力,种群规模逐步扩大并替代阿尔泰针茅成为退化草地群落的优势种群,赖草借助株高优势以及根状茎的生理整合能力[11,13],逐步成为退化草地群落的伴生种。退化草地群落狼毒和赖草地位的变化必然引起种群的分布格局和关联性发生变化,是认识干旱半干旱区草原植物对干扰和竞争响应特征的重要途径。目前,学界在退化草地中狼毒种群分布格局[14-15]和狼毒与阿尔泰针茅种群之间的关联性[16]、赖草分株种群对干旱胁迫和土壤资源异质性的响应[11,17]等方面进行了深入研究,但是草地退化过程中狼毒和赖草种群空间格局的尺度效应,及种内、种间竞争关系的转换机理尚不明晰。鉴于此,本文利用群落调查法与空间点格局分析法相结合,研究了祁连山北坡山地草原中狼毒与赖草种群的空间分布格局及关联性,旨在认识物种在异质环境中空间格局及竞争关系的转换特征,以及退化草地植物种群应对放牧干扰的更新途径。
1 研究方法
1.1 研究区概况
东祁连山北坡地处河西走廊与青藏高原的过渡带,具有大陆性气候和山地垂直气候特征,生态环境脆弱,植被以荒漠和草原为主,在特定的地形地貌和气候条件下主要发育并形成了荒漠草原(1900—2450 m)、典型草原(2450—2800 m)和草甸草原(2800—3800 m),植物以旱生和中生草本以及灌木为主。研究区位于甘肃省肃南县马场滩草原(38°47'22.1″—38°52'11″N,99°45'11.7″—99°57'41.2″E),海拔 2610—2632 m,年均温1—2℃,≥0℃的年积温为2450℃,相对无霜期80d,年降水量270—350 mm,年蒸发量1480—1620 mm,相对湿度65%,土壤以山地栗钙土为主。原生植被属于典型草原亚型的阿尔泰针茅草原,主要植物有:阿尔泰针茅、狼毒、冷蒿(Artemisia frigida)、赖草、甘肃臭草(Melica przewalskyi)、扁穗冰草(Agropyron cristatum)、星毛委陵菜(Potentilla acaulis)等。
1.2 样地设置与调查分析
根据2001—2010年对祁连山北坡“毒杂草型”退化草地群落演替过程的观察研究[10,12],在地貌相对一致、地形平缓的冬春草场,结合草地放牧管理和草地群落学调查,根据草地群落的优势种、物种丰富度,以及群落主要毒杂草物种狼毒的平均高度、盖度和密度等指标选取了4户牧民的围栏草地作为实验样地,分别为:未退化草地(Ⅰ)、轻度退化草地(Ⅱ)、中度退化草地(Ⅲ)和重度退化草地(Ⅳ),各样地概况和群落特征详见表1。供试草地每年6—11月休牧,其余时间正常放牧。

表1 供试样地的主要特征指标Table 1 The chief characteristics of Plots
试验在2010年8月植物生长盛期进行,在每个围栏样地内随机设置3个2 m×2 m的样方,共设置12个样方,用1 m×1 m的样方框(样方框内小格子的面积为2 cm×2 cm)按从左到右,从上到下的顺序依次放置(样方框按正南正北方向放置)4次,进行草地群落调查、狼毒与赖草种群的空间格局及关联性调查[18]。实测并记录每个样方(2 m×2 m)内的植物种类、密度、株高和地上生物量;以样方框左下角顶点为基点,记录样方(2 m×2 m)中每个狼毒与赖草株丛的地面投影和相对位置,用坐标值直接表示距离[19]。在室内对每一个样方(2 m×2 m)按照取样时样方框放置的顺序进行拼接、合并,确定各群落优势种。
1.3 数据分析
1.3.1 点格局分析
位于研究区域内二维空间的点构成点事件,用k(d)函数来反映点事件空间布局的依赖程度[20],其定义如下:
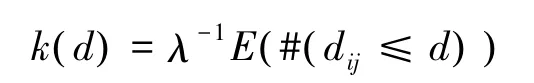
式中,λ表示研究区域内每单位面积点事件的密度,E表示在一定距离尺度下点事件数量的期望,#表示数量,i、j是两个具有相同特征的点事件,dij表示点i到点j的距离,d为距离尺度。如果一定距离尺度下点事件随机分布,则应有:
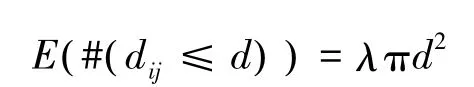
此时,k(d)=πd2;在距离尺度较小的情况下,若点事件为聚集分布,k(d)的观测值则将大于πd2。估计式为:

利用L(d)函数的估计值判断在此尺度下点事件的分布类型。距离尺度d下,若L(d)>0,为聚集分布;若L(d)=0,为随机分布;若L(d)<0,为均匀分布。

利用Monte Carlo方法求解空间分布的上下包迹线,若研究对象实际值落在上下包迹线的区间内,则此距离尺度下点事件为随机分布;如果实际值落在上包迹线之上,则为聚集分布;如果实际值落在下包迹线之下,则为均匀分布。
1.3.2 空间关联性分析
狼毒和赖草之间的空间关联性分析可表达为二者的点格局分析。将第一个种群k(d)写成k11(d),将第二个种群写成k22(d)。两个种群个体在距离(尺度)d内的数目为k12(d),其定义和计算原理与单种格局相近[21]。不难证明k12(d)可以用下式估计:
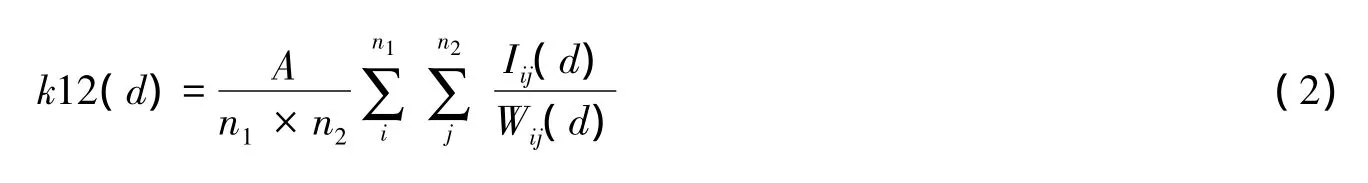
式中,n1和n2分别为种群1和种群2的个体数,A、Iij(d)和Wij(d)含义同(1)式,不同的是i和j分别代表种群1和2的个体,同样计算:

通常,当L12(d)=0表明两个种群间在d尺度无关联性,L12(d)>0为正关联,L12(d)<0为负关联。
利用Monte Carlo检验拟合包迹线,检验种群空间关联的显著性。若实际值落在上下包迹线的区间内,则此距离尺度下二者空间关联性不显著;若落在上包迹线之上,则为显著正关联;若落在下包迹线之下,则为显著负关联。本文中采用的步长为2 cm,Monte Carlo随机模拟的数目为20次,得到上下两条包迹线围成的95%置信区间。
1.3.3 种群领地密度计算
通常,种群领地密度是在确定种群空斑面积的基础上完成的。本文参考王鑫厅等对羊草(Leymus chinensis)种群空斑面积和种群领地密度的计算方法[22],若在单位方格中(2 cm×2 cm)没有种群株丛的地上投影出现,则称之为1个单位面积的空斑,以确定种群空斑面积的大小。将取样面积内除去种群空斑面积余下的区域称为种群领地,单位领地面积内的种群个体数称为种群领地密度。即种群领地=取样面积-种群空斑总面积,种群领地密度=种群个体数/种群领地面积。
2 结果与分析
2.1 狼毒和赖草种群特征分析
狼毒和赖草种群特征在不同退化草地中存在差异(表2)。狼毒地上生物量和种群领地密度均有显著的差异(F=18.77,P<0.01;F=13.43,P<0.01),随着草地退化,地上生物量和种群领地密度持续增加;赖草种群高度、地上生物量和领地密度也存在显著差异(F=15.87,P<0.01;F=10.02,P<0.01;F=18.97,P<0.01),随着草地退化,高度和种群领地密度逐渐降低,地上生物量先增大后减小。

表2 不同退化梯度狼毒和赖草种群特征Table 2 Ppulation characteristics of Stellera chamaejasme and Aneurolepidium dasystachys in different degradation gradient type
2.2 狼毒种群的空间分布格局
图1表明,不同退化梯度中,狼毒种群在不同尺度上空间点格局特征存在明显差异。未退化草地中,53—58 cm尺度上呈聚集分布;轻度退化草地中,聚集格局尺度转换为63—100 cm;中度退化草地中,在46—50 cm和64—85 cm呈聚集分布;重度退化草地中,在22—41 cm尺度上呈均匀分布。狼毒种群的分布格局统计上是极其显著的(P<0.01)。

图1 狼毒种群的点格局分析Fig.1 Point pattern analysis of Stellera chamaejasme population
2.3 赖草种群的空间分布格局
赖草种群空间格局以聚集分布和随机分布为主,且聚集分布格局在不同退化梯度上具有明显的尺度转换效应(图2)。未退化草地中,赖草种群在10—51 cm尺度上呈聚集分布格局;轻度退化草地中,聚集格局转换为4—9 cm和37—100 cm;中度退化草地中,8—36 cm尺度上表现为聚集分布;重度退化草地中,聚集格局仅在18—34 cm尺度上分布。赖草种群的分布格局统计上是极其显著的(P<0.01)。
2.4 狼毒与赖草种群的空间关联性
不同退化梯度中,狼毒与赖草的空间关联性随空间尺度变化表现出不同的关联类型(图3)。狼毒和赖草种群空间关联性在未退化至中度退化草地中,以正相关和不相关为主:未退化草地中在8—20 cm尺度上呈现正相关,轻度退化草地中仅在66—71 cm出现正相关关联性,中度退化草地在0—100 cm尺度上表现为关联性不显著;重度退化草地以负相关和不相关为主,在16—28 cm尺度上表现为显著负相关。狼毒与赖草的空间关联统计上是显著的(P<0.01)。
3 结论与讨论
干扰影响下特定植物群落中,微景观格局破碎化通常会造成群落资源与空间容量下降,引起具备较强更新能力的种群个体倾向于分享相邻个体的基础资源与空间部分,加剧了植物种群空间格局及竞争关系的不确定性[7]。本文选取的草地类型同狼毒与阿尔泰针茅种群空间关联研究草地一致,基本上反映了研究区“毒杂草型”退化草地的形成过程[16]。草地退化过程中,建群种阿尔泰针茅株丛死亡和草毡表层破碎,释放了部分资源和空间,狼毒与赖草等物种利用群落中剩余资源拓殖,一方面与阿尔泰针茅等建群物种竞争资源,另一方面,需要相互排斥实现对剩余资源的支配,从而植物竞争关系在特定阶段表现出不同的特征,空间格局发生尺度转换。
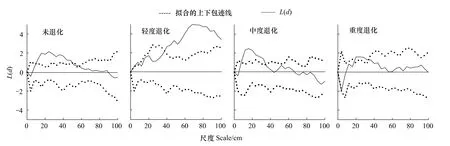
图2 赖草种群的点格局分析Fig.2 Point pattern analysis of Aneurolepidium dasystachys population

图3 狼毒和赖草种群空间关联性分析Fig.3 Spatial association analysis of Stellera chamaejasme and Aneurolepidium dasystachys population
种群分布格局是植物种群生物学特性对环境条件长期适应和选择的结果[23-24]。由于植物与环境之间存在着不间断的相互作用、相互影响,植物种群的分布格局总是随着群落环境的变化而发生变化[9,24]。未退化草地中,阿尔泰针茅形成的草毡层有助于加强对群落资源的控制作用[16],以幼株为主的狼毒种群,对群落资源的竞争相对较弱,为了抵御种间竞争和提高存活率,他们需要相互庇护形成团体竞争资源[14,16],从而狼毒种群在53—58 cm呈聚集分布。轻度退化草地中,家畜的过度践踏和啃食破坏了阿尔泰针茅草毡层,狼毒趁机突破限制,在过剩资源的驱动下迅速拓殖,并且凭借家畜不采食特性占据资源竞争的优势地位,种群规模迅速扩大形成聚块,引起聚集尺度区间转换为63—100 cm,狼毒种群地上生物量变化从16.43 g/m2增大至45.42 g/m2,也表明了种群对群落过剩资源的迅速利用状况。中度退化草地聚集尺度进一步转换为46—50 cm和64—85 cm,主要是随着狼毒逐渐成为群落优势种,资源的利用方式也从以争夺过剩资源为主转变为重新分配为主,狼毒能够进入较大级别的株丛的数量减少,聚块开始扩散[14-15],以及狼毒在地下10—30 cm土层形成密集的根系层[10],引起种群竞争关系由以种间竞争为主转向种内竞争,造成聚集尺度区间减小;且随草地进一步退化,狼毒种内竞争加剧引起重度退化草地22—41 cm尺度上出现均匀分布格局。
退化初期赖草种群以聚集格局为主且聚集区间增大,主要是因为赖草植株叶片宽大、茎叶稍硬[13],家畜首先选择采食适口性较好的阿尔泰针茅,赖草在拓殖能力驱动下,通过产生分株占领阿尔泰针茅退化所释放的资源空间。但伴随着狼毒种群的崛起,群落资源在狼毒种群控制下重新分配[14],赖草对资源的利用限于狼毒未充分利用部分,与此同时家畜开始大量采食导致植株的高度从67.11 cm下降至30.95 cm,降低了种群竞争光照资源的能力,赖草可利用资源受到限制,赖草地上生物量从18.51 g/m2减小至11.43 g/m2,种内个体更多的表现出对有限资源的争夺,引起聚集尺度在中度至重度退化草地中由8—36 cm转换为18—34 cm。赖草根茎克隆特性可能也对种群聚集区间转换产生了一定的影响,在土壤水分有效性较低条件下,通过根茎延伸分株的定植范围,在较大空间内置放分株,以便于种群尽快越过不利生境斑块,提高种群个体的存活概率[17],这可能也是退化草地中赖草非均匀分布的主要原因,需要进一步从种群无性繁殖特性深入研究。
种间关联理论认为,正相关是由于一个物种依赖于另一个物种,或者异质环境中几个物种对环境条件有相似的适应和反应引起的;负相关则是二者在竞争资源中互相排斥,或者环境需求的不相似性综合作用的结果[4,16]。狼毒与赖草种间空间关联性的动态特征可能反映了优势种更替过程中物种间竞争与依存的复杂关系。未退化和轻度退化草地关联性以正相关和不相关为主,正相关尺度从8—20 cm转换为66—71 cm,随后在中度和重度退化草地中转变为不相关和负相关,可能是因为草地退化初期,狼毒和赖草种群的主要竞争对象是阿尔泰针茅,在自身所控制的有限空间内,相互协作的伴生种间关系有助于共同抵御优势种的竞争压力,提高个体存活率,但随着狼毒逐渐成为群落优势种,阻碍了这种种间协作关系的延续,在种群拓殖能力的驱动下,植物个体间更多的表现为对阿尔泰针茅株丛破碎、死亡所留下的空斑和资源的争夺,同时阿尔泰针茅草毡层破碎[12],土壤水分蒸散速率加快使群落环境趋于干旱化,赖草无性系稳长深扎的适应对策加剧了二者根系生态位的重叠程度[11],使种间关系由协作关系逐渐转向竞争关系,随着退化程度加剧,种间竞争激烈程度增大,从而空间关联性转变为负相关。值得一提的是中度至重度退化草地狼毒种群同时表现出强烈的种内和种间竞争关系,可能是因为种内竞争有助于狼毒种群整合资源实现种群扩散,种间竞争则可能是狼毒排斥具有相似资源生态位需求物种的途径。草地退化过程中,狼毒种群领地密度从12株/m2增长至41.67株/m2,赖草从701.42株/m2减小至253.78株/m2的变化规律,也验证了二者非对称性竞争关系,由此推测这种同步提高种内、种间竞争强度的竞争关系,可能有助于狼毒种群排除其他竞争物种,提升群落中的地位。
综上所述,狼毒和赖草的空间格局随物种在群落中的地位更替表现出尺度转换效应,反映了植物种间、种内个体间竞争关系的转化过程。对于逐渐演变为群落优势种的狼毒种群,同步提升种内、种间竞争强度的更新途径,可能更有利于提升种群在群落中的地位和排除具有相似资源生态位需求的竞争物种;而赖草在非对称性资源竞争影响下,种群可利用资源受到限制,种群成为群落伴生物种。因此,干扰影响下,植物群落演替过程中物种的空间格局及竞争关系的转变可能是草地退化的关键因素,反映了退化草地植物种群应对放牧干扰的繁殖与更新的适应性途径。
[1] Yang X H,Zhang J T,Wu B,Li X S,Zhang Y Y.Point pattern analysis of Artemisia Ordosica population in the Mu Us sandy land.Chinese Journal of Plant Ecology,2006,30(4):563-570.
[2] Zhang J T.Quantitative Ecology.Beijing:Science Press,2004.
[3] Condit R,Ashton P S,Baker P,Bunyavejchewin S,Gunatilleke S,Gunatilleke N,Hubbell S P,Foster R B,Itoh A,LaFrankie J V,Lee H S,Losos E,Manokaran N,Sukumar R,Yamakura T.Spatial patterns in the distribution of tropical tree species.Science,2000,288(5470):1414-1418.
[4] Liu J H,Gao H,Zhang L H,Chen L P,Zhao N X,Gao Y B.Comparative analysis of inter-specific association within the Stipa grandis-S.krylovii communityin typical steppe of Inner Mongolia,China.Chinese Journal of Plant Ecology,2010,34(11):1016-1024.
[5] Greig-Smith P.Quantitative Plant Ecology.London:Blackwell Scientific Publications,1983.
[6] Li B,Chen J K,Andrew R W.A literature review on plant competition.Chinese Bulletin of Botany,1998,15(4):18-29.
[7] Zhao D L,Liu Z L,Yang G X,Xin X P,Liang C Z,Wang W,Wang L X.Grazing impact on distribution pattern of the plant communities and populations in Stipa krylovii steppe.Acta Prataculturae Sinica,2010,19(3):6-13.
[8] Niu K C,Luo Y J,Choler P,Du G Z.The role of biomass allocation strategy in diversity loss due to fertilization.Basic and Applied Ecology,2008,9(5):485-493.
[9] Wang Y G,Yang X H,Ci L J.Analysis of spatial pattern and competition relationship of shrub community in arid desert of Western Ordos Plateau.Journal of Plant Resources and Environment,2010,19(2):8-14.
[10] Zhao C Z,Fan S Y,Yin C Q,He X B.Study on vegetation community's structure of degraded grassland of noxious and miscellaneous grass type.Journal of Desert Research,2004,24(4):507-512.
[11] Ren A Z,Gao Y B,Liang Y,Chen S P,Liu S,Liu N.Effect of drought stress on clonal growth of Pennisetum Centrasiaticum and Leymus Secalinus.Journal of Desert Research,1999,19(Suppl 1):30-34.
[12] Zhao C Z,Ren H,Sheng Y P,Gao F Y,Shi F X.Fine-scale spatial point patterns of Stipa krylovii population in different alpine degraded grasslands.Acta Ecologica Sinica,2011,31(21):6388-6395.
[13] Institute of Botany,the Chinese Academy of Sciences.Iconographia Cormophytorum Sinicorum(tomus V).Beijing:Science Press,1976:87.
[14] Zhao C Z,Gao F Y,Wang X P,Sheng Y P,Shi F X.Fine-scale spatial patterns of Stellera chamaejasme population in degraded alpine grassland in upper reaches of Heihe,China.Chinese Journal of Plant Ecology,2010,34(11):1319-1326.
[15] Xing F,Song R.Population distribution pattern and dynamics of poisonous Stellera chamaejasme on grassland.Pratacultural Science,2002,19(1):16-19.
[16] Zhao C Z,Ren H.Fine-scale spatial associations of Stipa krylovii and Stellera chamaejasme population in alpine degraded grassland.Acta Ecologica Sinica,2011,31(20):6080-6087.
[17] Zhu X W,Liu H D,Liang S C,Ye Y Z,Dong M.Hetergeneityies of Leymus secalinus ramet population and it's soil resources in the Otindag Sandland.Acta Ecologica Sinica,2004,24(7):1459-1464.
[18] Chen B R,Yang G X,Zhang H B,Duan Q W,Xin X P.Spatial pattern analysis of Leymus chinensis population under different disturbances.Acta Ecologica Sinica,2010,30(21):5868-5874.
[19] Ripley B D.Spatial Statistics.New York:Wiley,1981:10-200.
[20] Diggle P.Statistical Analysis of Spatial Point Patterns.New York:Academic Press,1983.
[21] Manly B F J.Randomization and Monte Carlo Methods in Biology.New York:Chapman and Hal,1991.
[22] Wang X T,Wang W,Liang C Z.Changes in the population spatial distribution pattern of Leymus chinensis in degraded steppe community during restorative succession in Inner Mongolia,China.Chinese Journal of Plant Ecology,2009,33(1):63-70.
[23] Gittins R.Canonical Analysis;a Review With Applications in Ecology.Berlin:Sprinter Verlag,1985.
[24] Chao Z G,Wang S P,Xu G P,Hu Y G,Zhang Z H.Effect of fencing on community structure and distribution patterns of main populations in degraded Kobresia humilis Meadow.Acta Botanica Boreali-Occidentalia Sinica,2008,28(11):2320-2326.
参考文献:
[1] 杨洪晓,张金屯,吴波,李晓松,张友炎.毛乌素沙地油蒿种群点格局分析.植物生态学报,2006,30(4):563-570.
[2] 张金屯.数量生态学.北京:科学出版,2004.
[4] 刘珏宏,高慧,张丽红,陈丽萍,赵念席,高玉葆.内蒙古锡林郭勒草原大针茅-克氏针茅群落的种间关联特征分析.植物生态学报,2010,34(11):1016-1024.
[6] 李博,陈家宽,A.R.沃金森.植物竞争研究进展.植物学通报,1998,15(4):18-29.
[7] 赵登亮,刘钟龄,杨桂霞,辛小平,梁存柱,王炜,王立新.放牧对克氏针茅草原植物群落与种群格局的影响.草业学报,2010,19(3):6-13.
[9] 王彦阁,杨晓晖,慈龙骏.西鄂尔多斯高原干旱荒漠灌木群落空间分布格局及其竞争关系分析.植物资源与环境学报,2010,19(2):8-14.
[10] 赵成章,樊胜岳,殷翠琴,贺学斌.毒杂草型退化草地植被群落特征的研究.中国沙漠,2004,24(4):507-512.
[11] 任安芝,玉葆,梁宇,陈世苹,刘爽,刘宁.白草和赖草无性系生长对干旱胁迫的反应.中国沙漠,1999,19(增刊1):30-34.
[12] 赵成章,任珩,盛亚萍,高福元,石福习.不同高寒退化草地阿尔泰针茅种群的小尺度点格局.生态学报,2011,31(21):6388-6395.
[13] 中国科学院北京植物研究所.中国高等植物图鉴(第五册).北京:科学出版,1976:87-87.
[14] 赵成章,高福元,王小鹏,盛亚萍,石福习.黑河上游高寒退化草地狼毒种群小尺度点格局分析.植物生态学报,2010,34(11):1319-1326.
[15] 邢福,宋日.草地有毒植物狼毒种群分布格局及动态.草业科学,2002,19(1):16-19.
[16] 赵成章,任珩.退化草地阿尔泰针茅与狼毒种群的小尺度种间空间关联.生态学报,2011,31(20):6080-6087.
[17] 朱选伟,刘海东,梁士楚,叶永忠,董鸣.浑善达克沙地赖草分株种群与土壤资源异质性分析.生态学报,2004,24(7):1459-1464.
[18] 陈宝瑞,杨桂霞,张宏斌,段庆伟,辛晓平.不同干扰类型下羊草种群的空间格局.生态学报,2010,30(21):5868-5874.
[22] 王鑫厅,王炜,梁存柱.典型草原退化群落不同恢复演替阶段羊草种群空间格局的比较.植物生态学报,2009,33(1):63-70.
[24] 晁增国,汪诗平,徐广平,胡宜刚,张振华.围封对退化矮嵩草草甸群落结构和主要种群空间分布格局的影响.西北植物学报,2008,28(11):2320-2326.
