互花米草与短叶茳芏枯落物分解过程中碳氮磷化学计量学特征
2013-09-11欧阳林梅王维奇
欧阳林梅,王 纯,王维奇,2,* ,仝 川,2
(1.福建师范大学亚热带湿地研究中心,福州 350007;2.福建师范大学地理研究所,福州 350007)
生态化学计量学为研究碳、氮、磷等主要元素的生物地球化学循环和生态学过程提供了一种新思路,是当前生态学研究的前沿领域之一[1-2]。近年来,生态学家对该领域的关注日益增强,并取得了丰硕的成果[3-6],所涉及的领域包括对植物组织、土壤和枯落物元素生态化学计量学的研究[7-11],主要是对陆地生态系统和水生生态系统元素生态化学计量学开展了相关研究[3-11],对介于陆地和水生生态系统过渡带的湿地生态系统元素生态化学计量学的研究还十分有限[12-13],湿地枯落物分解过程是湿地生物地球化学循环过程中的关键一环,是植物生长所需养分的主要来源[14],其元素生态化学计量学的研究对揭示枯落物分解速率以及养分的积累和释放机制具有重要意义[5]。
短叶茳芏(Cyperus malaccensis var.brevifolius)是闽江河口鳝鱼滩湿地的主要优势植物,近年来,互花米草(Spartina alterniflora)迅速入侵,并占据从低潮滩到高潮滩各种生境,目前,这两种植物已成为闽江河口鳝鱼滩湿地最为主要的湿地植物群落。作者在探讨了闽江河口湿地不同环境条件下土壤、植被及其所产生的枯落物碳氮磷生态化学计量学特征的基础上[13,15-16],对互花米草和短叶茳芏枯落物分解过程中的生态化学计量学特征的变化规律及其对分解速率、养分和能量动态的指示作用等问题作一探讨,对完善生态化学计量学理论具有重要意义。
1 材料与方法
1.1 研究区概况
选取闽江河口鳝鱼滩湿地为研究区域,区内气候暖热湿润,年均温19.3℃,年降水量1346 mm。天然植被主要有芦苇(Phragmites australis)、短叶茳芏、藨草(Scirpus triqueter)和近几年外来入侵的互花米草等[17],在近潮沟生境湿地受潮汐的影响除小潮日外均淹水(高淹水频率),远潮沟生境则仅在大潮日淹水(低淹水频率),远近潮沟生境之间样地相距约500 m,互花米草和为短叶茳芏在两种生境均有分布,这为我们探讨不同生境枯落物分解过程中碳、氮、磷的生态化学计量学特征及其指示意义研究提供了理想的实验地。2007年5—12月潮水盐度平均值为4.2,12月份潮水中硫酸盐含量为714 mg/L[18]。
1.2 土壤样品处理及测定
2007年1月在闽江河口鳝鱼滩,按照生境差异设置两个样点,一个为高淹水频率的近潮沟生境,另一个是低淹水频率的远潮沟生境,为了保证实验所用枯落物来源的一致性,在两种生境样地之间,短叶茳芏和互花米草分别占绝对优势的地段,剪取立枯体,带回实验室用自来水冲洗立枯体表面污泥,然后剪成10 cm左右小段,风干,在70℃ 下烘干至恒重,装袋(孔径0.2 mm,规格为20 cm×25 cm),每袋称重25 g。2007年1月7日将系有尼龙绳的分解袋栓到事先分别在两种生境样地埋设好的木桩上,木桩分别埋设在两种生境样地两种植物占优势的群落内,各埋设3个木桩。分解袋随机分布在以木桩为圆心、约1.5 m为半径的范围内。分解袋投放后的20、67、97、127、157、186、218、249、280 d 分别取回 12 袋(每种植物每个样地分别取 3 袋),回实验室后捡去枯落物中的杂质,清洗干净枯落物表面,放烘箱中在70℃下烘干至恒重并称重,粉碎,过100目孔筛。植物碳、氮元素含量采用碳氮元素分析仪(Vario EL III,德国生产)测定,磷采用钼锑抗比色法(UV-2450,日本生产)测定,热值采用微电脑数显两用热量计(WGR-WSR,中国生产)测定。
1.3 数据处理
应用Excel 2003和SPSS 13.0统计分析软件对测定数据进行整理。原始数据的处理采用Excel 2003,应用SPSS13.0统计分析软件中的成对样本T检验对不同生境互花米草和短叶茳芏枯落物分解过程中的碳、氮、磷含量、热值以及C/N、C/P、N/P进行差异性检验。枯落物C/N、C/P、N/P采用的是物质的量之比(mol/mol)。
2 结果与分析
2.1 互花米草和短叶茳芏枯落物分解速率
整个观测期内(280d),在近潮沟生境,互花米草枯落物和短叶茳芏枯落物的干物质损失量分别为初始量的71.1%和75.2%,分解速率分别为0.00414/d和0.00528/d;远潮沟生境,互花米草枯落物和短叶茳芏枯落物的干物质损失量分别为初始量的67.3%和70.7%,分解速率分别为0.00368/d和0.00413/d。互花米草和短叶茳芏分解速率表现为在近潮沟生境大于在远潮沟生境。两种生境均表现为互花米草枯落物分解速率低于短叶茳芏枯落物分解速率。
2.2 互花米草和短叶茳芏枯落物分解过程中碳、氮、磷含量变化
在近潮沟生境和远潮沟生境,互花米草枯落物和短叶茳芏枯落物在280 d的分解期内平均C、N、P含量均表现为C>N>P(P<0.01);两种生境均表现为互花米草枯落物分解过程中的N、P含量低于短叶茳芏(P<0.05),C含量差异不显著;对于同一种植物枯落物分解过程而言,在近潮沟生境与远潮沟生境其分解过程中C、N、P含量差异不显著(表1)。

表1 枯落物分解过程中C、N、P含量特征Table 1 C,N,P content during litter decomposition
2.3 互花米草和短叶茳芏枯落物分解过程中热值变化
在近潮沟生境互花米草枯落物和短叶茳芏枯落物在280 d的分解期内平均热值变化为15.7—18.0 kJ/g和15.0—18.9kJ/g,平均值分别为16.8kJ/g和16.5kJ/g,互花米草枯落物分解过程中的热值和短叶茳芏枯落物分解过程中的热值差异不显著;远潮沟生境,互花米草枯落物和短叶茳芏枯落物在280 d的分解期内平均热值变化为16.2—18.8kJ/g 和14.7—17.1kJ/g,平均值分别为17.4kJ/g 和15.4kJ/g,互花米草枯落物分解过程中的热值极显著高于短叶茳芏枯落物分解过程中的热值(P<0.01);对于同一种植物枯落物分解过程而言,在近潮沟生境与远潮沟生境其分解过程中热值差异不显著。
2.4 互花米草和短叶茳芏枯落物分解过程中C/N、C/P和N/P动态
从枯落物分解的整个观测期的均值看,近潮沟生境和远潮沟生境互花米草枯落物和短叶茳芏枯落物分解过程中的C/P明显高于C/N和N/P(P<0.01)。两种生境均表现为互花米草枯落物分解过程中的C/N、C/P高于短叶茳芏(P<0.01),N/P差异不显著。对于同一种植物枯落物分解过程而言,在近潮沟生境与远潮沟生境其分解过程中C/N、C/P和N/P差异不显著(图1)。此外,在近潮沟生境和远潮沟生境,互花米草枯落物和短叶茳芏枯落物分解过程中C/N、C/P和N/P的变化趋势看,两种植物枯落物分解过程中C/N均表现为整体下降的趋势,C/P和N/P均表现为先上升后下降的趋势(图1)。
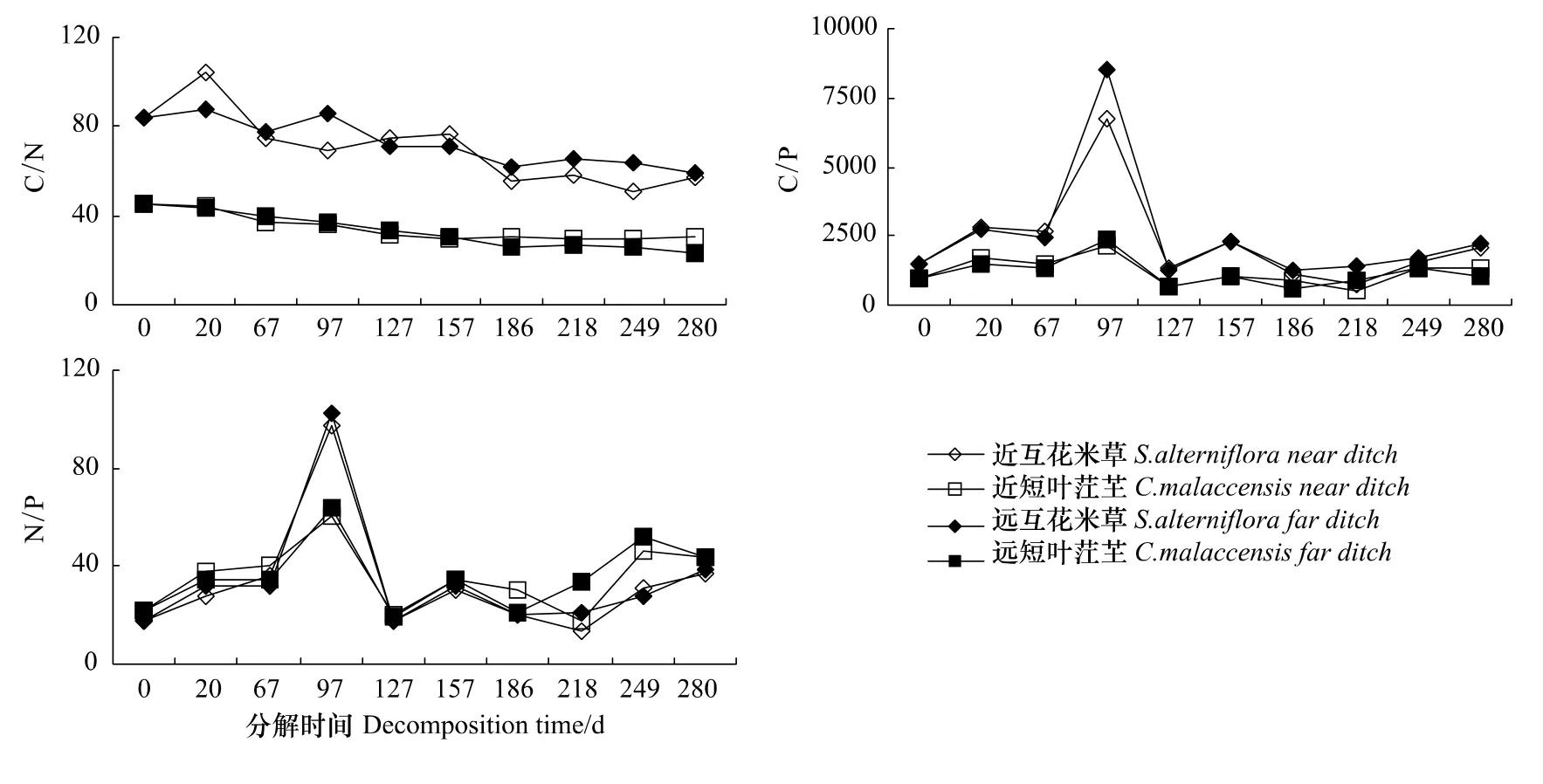
图1 枯落物分解过程中C/N、C/P、N/P动态Fig.1 Dynamics of C/N、C/P、N/P during litter decomposition
3 讨论
枯落物分解过程是以碳为主导的物质循环模式,随着分解过程的进行,残留枯落物的氮、磷等养分发生积聚或释放,进而改变着枯落物分解过程中的碳、氮、磷比[14]。本研究中,在近潮沟生境和远潮沟生境互花米草和短叶茳芏枯落物分解过程中C/N均表现为整体下降的趋势,C/P和N/P均表现为先上升后下降的趋势(图1)。C/N在分解过程中的变化主要与枯落物分解过程中碳含量的变化不大,氮含量发生了明显的富集有关。因此,氮是调节枯落物分解过程中C/N变化的关键因子。C/P和N/P主要与枯落物分解过程中磷含量的释放—吸收—释放的波动性变化模式有关,受碳、氮含量变化的影响不大。因此,磷是调节枯落物分解过程中C/P和N/P变化的关键因子。
在近潮沟生境和远潮沟生境,互花米草枯落物分解过程中的C/N和C/P高于短叶茳芏湿地枯落物,这一碳与养分生态化学计量比对枯落物分解过程的指示作用主要表现在以下几个方面:第一,互花米草枯落物的分解速率和碳与养分的生态化学计量比呈负相关,这与较低的C/N、C/P具有较高的碳利用效率密切相关[14];第二,互花米草枯落物的分解过程中养分的释放潜力与碳与养分的生态化学计量比呈负相关,主要是因为参与枯落物分解过程的微生物生长遵循着较为严格的养分生态化学计量比需求[10,19],其C/N需求为9.33—17.50,C/P 需求为 51.67—266.67[14],这些比值明显低于枯落物。因此,为了维持微生物的正常生长,枯落物中碳与养分的生态化学计量比越高,对养分的需求与束缚能力越强,从而释放养分的潜力越弱,实际上碳与养分的生态化学计量比对枯落物分解过程中养分释放的影响是一个极其复杂的过程,可能与控制枯落物磷元素积累和释放的生态化学计量比的阈值、分解者在枯落物分解过程中对枯落物质量的适应性[14,20-21]以及枯落物分解前期的淋溶作用有关[22];第三,互花米草枯落物的分解过程中能量的释放潜力与碳与养分的生态化学计量比呈负相关,根据Meehan[23]的研究结论,可能与较低的碳与养分比值可以降低枯落物分解酶物质生产过程中能量消耗水平,促使更多的能量释放到环境中有关。
综上所述,枯落物分解过程中的元素生态化学计量学特征及其指示作用的变化是十分复杂。因此,更为深入地的机理问题,包括量化控制枯落物中养分和能量释放的关键阈值及其微生物种群、不同分解者种群对碳的利用效率和碳与养分比值之间的关系等问题[4],都有待于深入研究。
[1] Güsewell S.N ∶P ratios in terrestrial plants:variation and functional significance.New Phytologist,2004,164(2):243-266.
[2] He J S,Han X G.Ecological stoichiometry:searching for unifying principles from individuals to ecosystems.Chinese Journal of Plant Ecology,2010,34(1):2-6.
[3] Elser J J,Bracken M E S,Cleland E E,Gruner D S,Harpole W S,Hillebrand H,Ngai J T,Seabloom E W,Shurin J B,Smith J E.Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater,marine and terrestrial ecosystems.Ecology Letters,2007,10(12):1135-1142.
[4] Keiblinger K M,Hall E K,Wanek W,Szukics U,Hämmerle I,Ellersdorfer G,Böck S,Strauss J,Sterflinger K,Richter A,Zechmeister-Boltenstern S.The effect of resource quantity and resource stoichiometry on microbial carbon-use-efficiency.FEMS Microbiology Ecology,2010,73(3):430-440.
[5] Marichal R,Mathieu J,Couteaux M M,Mora P,Roy J,Lavelle P.Earthworm and microbe response to litter and soils of tropical forest plantations with contrasting C ∶N ∶P stoichiometric ratios.Soil Biology and Biochemistry,2011,43(7):1528-1535.
[6] Ågren G I.Stoichiometry and nutrition of plant growth in nature communities.Annual Review of Ecology,Evolution,and Systematics,2008,39:153-170.
[7] Tessier J T,Raynal D J.Use of nitrogen to phosphorus ratios in plant tissue as indicator of nutrient limitation and nitrogen saturation.Journal of Applied Ecology,2003,40(3):523-534.
[8] Zhang L X,Bai Y F,Han X G.Differential responses of N:P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol.Acta Botanica Sinica,2004,46(3):259-270.
[9] Gao S P,Li J X,Xu M C,Chen X,Dai J.Leaf N and P stoichiometry of common species in successional stages of the evergreen broad-leaved forest in Tiantong National Forest Park,Zhejiang Province,China.Acta Ecologica Sinica,2007,27(3):947-952.
[10] Cleveland C C,Liptzin D.C:N:P stoichiometry in soil:is there a“Redfield ratio”for the microbial biomass?Biogeochemistry,2007,85(3):235-252.
[11] Tian H Q,Chen G S,Zhang C,Melillo J M,Hall C A S.Pattern and variation of C ∶N ∶P ratios in China's soils:a synthesis of observational data.Biogeochemistry,2010,98(1/3):139-151.
[12] Koerselman W,Meuleman A F M.The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation.Journal of Applied Ecology,1996,33(6):1441-1450.
[13] Wang W Q,Tong C,Jia R X,Zeng C S.Ecological stoichiometry characteristics of wetland soil carbon,nitrogen and phosphorus in different waterflooded frequency.Journal of Soil and Water Conservation,2010,24(3):238-242.
[14] Manzoni S,Trofymow J A,Jackson R B,Porporato A.Stoichiometric controls on carbon,nitrogen,and phosphorus dynamics in decomposing litter.Ecological Monographs,2010,80(1):89-106.
[15] Wang W Q,Tong C,Zeng C S.Stoichiometry characteristics of carbon,nitrogen,phosphorus and anaerobic carbon decomposition of wetland soil of different texture.China Environmental Science,2010,30(10):1130-1134.
[16] Wang W Q,Zeng C S,Zhong C Q,Tong C.Effect of human disturbance on ecological stoichiometry characteristics of soil carbon,nitrogen and phosphorus in Minjiang River estuarine wetland.Chinese Journal of Environmental Science,2010,31(10):2411-2416.
[17] Liu J Q,Zeng C S,Chen N.Research of Minjiang River Estuary Wetland.Beijing:Science Press,2006.
[18] Tong C,Liu B G.Litter decomposition and nutrient dynamics in different tidal water submergence environments of estuarine tidal wetland.Geographical Research,2009,28(1):118-128.
[19] Hättenschwiler S,Jørgensen H B.Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest.Journal of Ecology,2010,98(4):754-763.
[20] Bragazza L,Siffi C,Lacumin P,Gerdol R.Mass loss and nutrient release during litter decay in peatland:the role of microbial adaptability to litter chemistry.Soil Biology and Biochemistry,2007,39(1):257-267.
[21] Manzoni S,Jackson R B,Trofymow J A,Porporato A.The global stoichiometry of litter nitrogen mineralization.Science,2008,321(5889):684-686.
[22] Cleveland C C,Reed S C,Townsend A R.Nutrient regulation of organic matter decomposition in a tropical rain forest.Ecology,2006,87(2):492-503.
[23] Meehan T D.Energy use and animal abundance in litter and soil communities.Ecology,2006,87(7):1650-1658.
参考文献:
[2] 贺金生,韩兴国.生态化学计量学:探索从个体到生态系统的统一化理论.植物生态学报,2010,34(1):2-6.
[8] 张丽霞,白永飞,韩兴国.内蒙古典型草原生态系统中N素添加对羊草和黄囊苔草N∶P化学计量学特征的影响.植物学报,2004,46(3):259-270.
[9] 高三平,李俊祥,徐明策,陈熙,戴洁.天童常绿阔叶林不同演替阶段常见种叶片N、P化学计量学特征.生态学报,2007,27(3):947-952.
[13] 王维奇,仝川,贾瑞霞,曾从盛.不同淹水频率下湿地土壤碳氮磷生态化学计量学特征.水土保持学报,2010,24(3):238-242.
[15] 王维奇,仝川,曾从盛.不同质地湿地土壤碳、氮、磷计量学及厌氧碳分解特征.中国环境科学,2010,30(10):1130-1134.
[16] 王维奇,曾从盛,钟春棋,仝川.人类干扰对闽江河口湿地土壤碳、氮、磷生态化学计量学特征的影响.环境科学,2010,31(10):2411-2416.
[17] 刘剑秋,曾从盛,陈宁.闽江河口湿地研究.北京:科学出版社,2006.
[18] 仝川,刘白贵.不同水淹环境下河口感潮湿地枯落物分解及营养动态.地理研究,2009,28(1):118-128.
