小兴安岭阔叶红松混交林林隙特征
2013-09-07刘少冲王敬华段文标陈立新王丽霞赵健慧
刘少冲,王敬华,段文标,* ,陈立新,王丽霞,杜 珊,赵健慧
(1.东北林业大学林学院,哈尔滨 150040;2.中国农村技术开发中心,北京 100045)
林隙是指森林群落中老龄树死亡或因偶然因素(如干旱、台风、火灾等)导致成熟阶段优势树种的死亡,从而在林冠层造成空隙的现象[1]。美国森林生态学家Runkle把林隙分为两类:(1)冠空隙或者实际林隙:是指直接处于林冠层隙下的土地面积或空间。(2)扩展林隙指由冠空隙周围树木的树干所围成的土地面积或空间,包括实际林隙和其边缘到周围树木树干基部所围成的面积或空间两部分[2]。
林隙形成直接影响着森林的结构,林隙内的生态环境,显著改变了资源的可利用程度,为植物种子萌发、苗木定居、幼树生长等自然更新过程提供了有利条件[3]。对林隙特征的研究已成为探讨森林群落结构动态,物种多样性的维持以及进行森林可持续经营管理的重要理论基础,对其研究成为生态学研究的热点和重点之一[4-5]。国外对林隙的特征、干扰和生物多样性维持等方面进行了深入研究[6-9]。我国一些学者对温带、亚热带和热带不同森林生态系统的林隙特征、干扰状况、树种更新等[10-14]进行了相关的探索,对林隙边缘木的偏冠现象、边缘木的数量等特征[15-16]进行了探讨,但是应用模型进行边缘木结构分布拟合的研究还为数不多[17-20]。总的看来,对林隙时空格局及干扰规律进行了较为广泛的探索。但是,迄今对小兴安岭阔叶红松混交林林隙特征的系统研究并不多见。为此,本文采用样线调查法,对小兴安岭凉水国家级自然保护区原始阔叶红松混交林林隙基本特征进行了调查和分析,主要探讨以下几个问题:(1)林隙的干扰频率和大小结构;(2)林隙的形成方式;(3)林隙形成木的种类组成和结构特征;(4)林隙边缘木的结构及偏冠率特征。通过这几项调查,了解阔叶红松混交林的变化规律,为小兴安岭凉水国家级自然保护区可持续经营提供理论参考和数据支持。
1 研究地区自然概况与研究方法
1.1 研究地区自然概况
研究地点设在黑龙江省伊春市带岭区凉水国家级自然保护区(47°7'39″—47°14'22″N,128°48'30″—128°55'50″E)。试验地位于该保护区的第12号林班的阔叶红松混交林内,海拔约400m,坡度4°—18°,土壤为暗棕壤,年平均气温-0.3℃,年平均最高气温7.5℃,年平均最低气温-6.6℃。年平均降水量676.0 mm,全年降水日数120—150 d,积雪期130—150 d,年平均相对湿度78%—96%,年平均蒸发量805.4 mm。年日照时数1850 h。年平均地温1.2℃,冻土约2.0 m。河流结冰期长达6个月,无霜期100—120 d。地带性植被是以红松(Pinus koraiensis)为主的针阔混交林,属温带针阔叶混交林地带北部亚地带植被。其组成以红松为主,伴生着紫椴(Tilia amurensis)、枫桦(Betula costata)、大青杨(Populus ussuriensis)、裂叶榆(Ulmus laciniata Mayr.)、五角槭(Acer mono)和花楷槭(Acer ukurunduense)等20余种阔叶树种,主要下木有刺五加(Acanthopanax senticosus)、毛榛子(Corylus mandshurica)、珍珠梅 (Sorbaria sorbifolia)、东北山梅花(Philadelphus schrenkii)等,草本植物主要有苔草(Carex spp.)、猴腿蹄盖蕨(Athyrium brevifrons Nakaiex Kitag)和小叶芹 (Aegopodium alpestra Ledeb)等。藤本植物有山葡萄(Vitis amurensis Rupr.)、五味子(Schisandra chinensis(Turez.)Baill.)、狗枣猕猴桃(Actinidia kolomikta)等。
1.2 研究方法
在阔叶红松混交林试验地内,采用样线调查法,随机设置2条相隔100m的样线。根据罗盘仪所指的方向,由西北向东南方向穿行,第1条样线为490m,第2条样线为800m,对样线中出现的每个林隙进行调查。用测绳实际测量走过的距离,测定始终由同一人完成。在每个林隙中心按照罗盘所指的北、东北、东、东南、南、西南、西、西北8个方向记录林隙的长度(分别按冠空隙和扩展林隙记载),确定林隙形成木(GM)的种类、数量、状态(分干中折断、干基折断、掘根风倒和枯立4类)、直径、树高、腐烂程度以及林隙边缘木的种类、胸径和高度。林隙年龄的确定采用查取轮枝法、生长锥法和估测法相结合的方法进行。对于有腐朽倒木且其上存在更新的幼苗、幼树和大树时,采用查取轮枝法、生长锥法确定林隙的年龄。对于已彻底分解且无法辨认倒木的林隙,采用估测法确定其年龄,即根据文献记载凉水自然保护区的开发年份和本地有经验的林业调查者综合判断。
1.3 数据处理
1.3.1 林隙面积及形状指数的计算
林隙大小的测定方法可归纳为:椭圆法、等角多边形法、三角形法、平面相片法、航片法、树冠投影的估算法以及把研究样地划分成一定大小的样方后,判断小样方是不是林隙的网格方法[6,8-9]。但由于各自的缺陷,没有一种方法被广泛采用。目前,等角八边形法仍然是估计林隙面积的主要方法。计算冠空隙(简称CG)和扩展林隙(简称EG)的面积采用等角多边形计算公式:A=0.5in(2π/n),式中ln+1等于l1(i=n+1也i1是第一个点,i=1)[21]。A为林隙面积(m2)。林隙周长的计算采用等角八边形的方法,得到其近似值。林隙形状指数计算公式[21]:SI=P/(2),式中SI为林隙形状指数;P为林隙周长(m);A为林隙面积(m2)。偏冠现象用偏冠率(简称RCI)来度量。它是指在林隙中心与边缘木的连线上,林隙内的最大枝条长度与该连线上树冠宽度的比值[15]。
1.3.2 Weibull分布模型
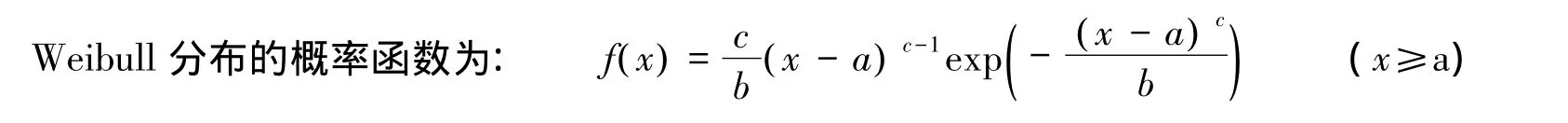
Weibull分布有3个参数,b>0为尺度参数,c>0为形状参数,a为位置参数。当c<1.0,Weibull分布呈倒J形;当1.0<c<3.6,呈正偏山状分布;c=3.6,近似于正态分布;c>3.6,转向负偏山状分布。因此,形状参数能较好地反映分布情况,是一个具有生态学意义的Weibull分布参数。Weibull分布参数求解方法很多,有最大似然估计、矩法、回归法、改进单纯形法[22-25]。本文采用三参数最大似然估计法,并用柯尔莫哥洛夫检验法评价威布尔分布模型对林隙冠空隙和扩展林隙面积以及边缘木径级和高度级拟合的适应程度。计算过程为:首先求出各组的理论频数,计算出累积概率F(xi),同时求出各组的实际频数ni,及其相应的实际累积频率Fn(xi),将F(xi)与Fn(xi)进行比较,以Δ=|F(xi)-Fn(xi)|及|F(xi)-Fn(xi-1)|反映理论分布与实际分布的差异程度,求算Dn=supΔ=sup{|F(xi)-Fn(xi)|,|F(xi)-Fn(xi-1)|},再找出Dn的临界值,通过理论的分布函数F(x)与样本累积频率分布函数Fn(x)的比较作出结论[17]。
1.3.3 正态分布

式中,¯x为随机变量x的数学期望;σ为随机变量x的标准差。
2 结果分析
2.1 林隙的干扰频率
运用样线调查法,沿2条样线穿行的总距离为1290m,线路上出现的林隙数总计41个,即平均每穿行100m,会遇到3.18个林隙,林隙的线状密度为31.78个/km。在调查样线上,处于扩展林隙中的线段总计397m,也就是说扩展林隙在整个林分中所占的比例平均为397/1290=30.78%,这个比例可以近似地理解为扩展林隙面积在林分面积中所占的百分比[5]。在所调查的41个林隙中,冠空隙的平均面积ACG=153.37m2,而扩展林隙的平均面积AEG=300.44m2,冠空隙和扩展林隙的相对比例平均为ACG/AEG=51.05%。所以,冠空隙在整个林分中所占的平均比例为30.78%×51.05%=15.71%。
在1290m的样线上,遇到的41个林隙中最老的林隙是70a,最年轻的林隙是1a,林隙的形成速率平均为41/1.290/(70-1)=0.46个·km-1·a-1。其中,冠空隙的形成速率为4.46×51.05%=2.28m·km-1·a-1,扩展林隙的形成速率为397/1.290/(70-1)=4.46m·km-1·a-1。以上分析表明:在阔叶红松混交林中,平均每年约有397/1290/(70-1)×100%=0.45%的面积转化为扩展林隙,约有0.45×51.05%×100%=0.23%的林冠面积转化为冠空隙,平均每年每公里上约有0.46个林隙出现,其中每年处于冠空隙中的线段长度平均每公里约有2.28m,而处于扩展林隙中的线段长度平均每公里有4.46m。照此计算,林隙干扰的周期为1/0.23%=434.8a。
2.2 林隙的大小结构
林隙的大小不仅可反映林隙内微环境因子的变动,而且还可提供林隙内更新所能利用的空间资源。根据林隙的两种不同定义,以50 m2为级别,采用上限排外法,统计各级内出现的林隙数和相对百分比以及各级面积总和所占百分比(表1,表2)。由表1可知,小兴安岭阔叶红松混交林冠空隙的面积以<200m2的居多,占总冠空隙数量的82.93%,>300 m2的冠空隙很少,占总林隙数量的4.88%。从面积百分比来看,100—200 m2的冠空隙面积最大,占57.19%,<50m2的所占面积最小,仅占0.67%。扩展林隙面积大多数在200—400m2之间,占扩展林隙数量的73.17%,小于100m2和大于500m2的扩展林隙较少,仅占扩展林隙数量的7.32%(表2)。

表1 小兴安岭阔叶红松混交林林冠空隙(CG)大小结构Table1 Size structure of CG in the mixed broad-leaved Korean pine forest in Xiaoxing'an Mountains
在所调查的41个林隙中,冠空隙的最小面积为42.12m2,最大为372.52m2,平均为153.37m2,而扩展林隙的最小面积为98.65m2,最大面积为633.10m2,平均为300.44m2。冠空隙平均面积约为扩展林隙平均面积的51.05%。根据冠空隙划分的6个级别来计算,每级平均林隙数为6.8333个,每m2有冠林隙0.0446个,即每100 m2有4.5个林隙,平均每个冠空隙占据22.44 m2;根据扩展林隙划分的9个级别来计算,每级平均林隙数为4.5556个,每m2有0.0152个扩展林隙,即每100 m2有1.5个扩展林隙,平均每个扩展林隙占据65.95 m2。综上所述,小兴安岭阔叶红松混交林林隙属中小林隙型,是森林更新演替的必要过程,进一步说明了小兴安岭阔叶红松混交林是比较稳定的森林群落。
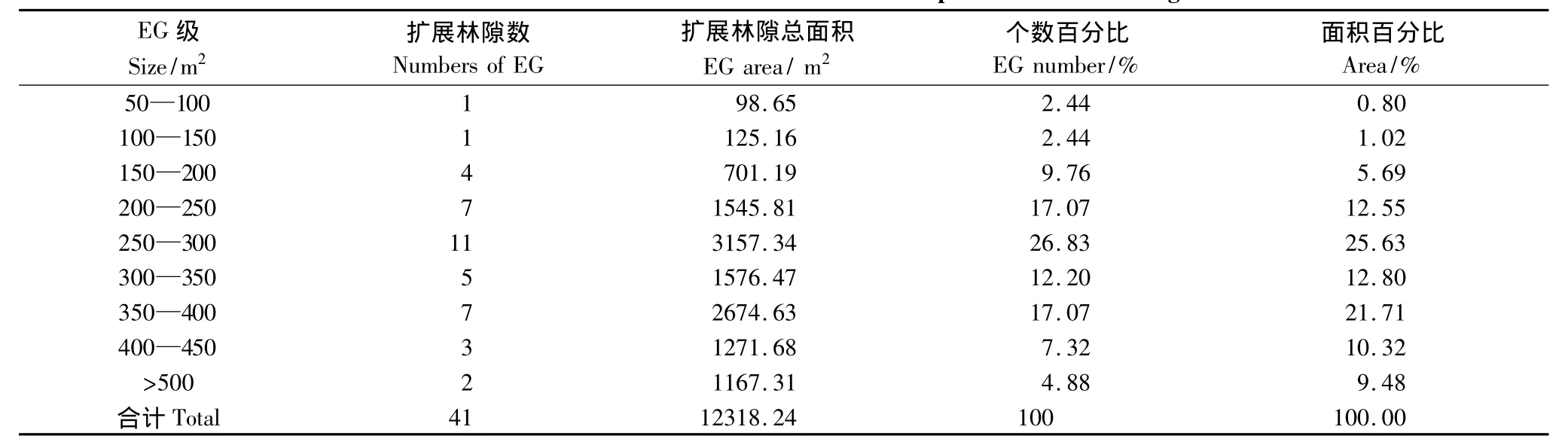
表2 小兴安岭阔叶红松混交林扩展林隙(EG)大小结构Table2 Size structure of EG in the mixed broad-leaved Korean pine forest in Xiaoxing'an Mountains
一些学者应用正态分布、对数正态分布、指数分布及Weibull分布模型对不同地域、不同林型的林隙进行了探讨,普遍认为Weibull分布在研究森林林隙面积大小分布规律时适用性更广,是一个理想的理论模型[17-19]。本文在数据预处理中采用了Weibull模型,高斯分布、对数正态和二阶对数拟合林隙大小结构,结果表明高斯分布、对数正态分布和二阶对数分布都无法对其进行模拟,因此予以剔除。应用Weibull分布模型对阔叶红松混交林林隙面积大小结构进行拟合与检验。结果表明,无论是林隙的冠空隙还是扩展林隙,面积分布均符合Weibull分布(表3)。

表3 林隙面积的Weibull分布拟合结果Table3 The simulated results of gap area by Weibull distribution
2.3 林隙的形成方式
林隙是树冠干扰的产物,林冠层在空间上形成不连续就表现为一定尺度的空隙。在调查的41个林隙中,最常见的林隙形成方式有以下4种:(1)主林层干中折断(即折断的高度在离开树基1.5m以上),大多是由于强风引起的,部分是由胸径较大的掘根倒木压断引起;(2)干基折断,一方面由风力产生,另一方面是因为树木衰老,基部强度减弱,风或重力等外力引起;(3)掘根风倒,是由于风力作用而使树木连根拔起,形成丘和坑,并且连根拔起的树木还会导致沿倾倒方向的树木大枝折断,这种林隙形成方式产生的林隙面积较大;(4)枯立,指树木已死亡,树枝全部或部分折断,枝叶完全脱落,主干直立于林中,产生的林隙面积较小。在调查的204株形成木中,干中折断的为43株(占21.08%),干基折断的为72株(占35.29%),掘根风倒的为58株(占28.43%),枯立的有31株(占15.20%)。从表4可以看出,小兴安岭阔叶红松混交林林隙是一种或几种方式共同形成的,主要方式是树干(干基和干中)折断,其次为掘根风倒,再次为枯立。
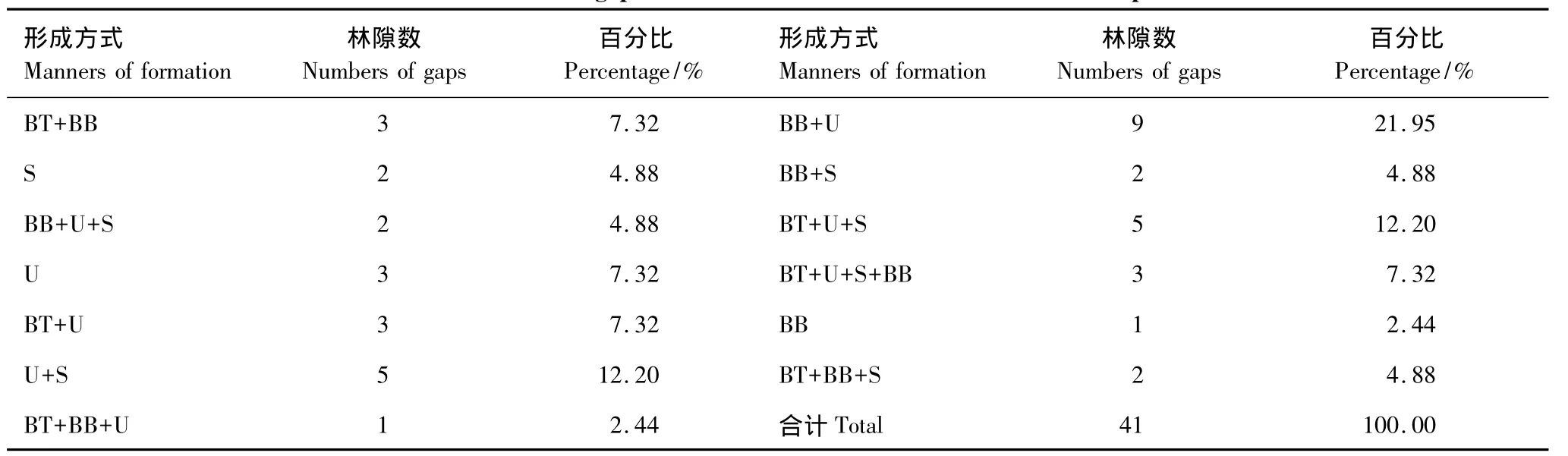
表4 阔叶红松混交林林隙的形成方式Table4 The manners of gap formation in the mixed broad-leaved Korean pine forest
2.4 单个林隙形成木的数量特征
森林中的林隙,有时由一株形成木形成,有时由多株形成木共同形成。对小兴安岭阔叶红松混交林林隙形成木的数量特征进行了统计(表5)。结果表明:有5株形成木形成的林隙最多,占24.39%;由3株、4株和6株形成的林隙共占51.21%;其它株数形成的林隙数量较少,占24.40%。可见,在小兴安岭阔叶红松混交林中,绝大多数的林隙是由3—6株形成木所形成的。在所调查的41个林隙中,共有204株形成木,平均每个林隙形成木为4.98株,每株形成木所能形成的冠空隙面积(CG)为30.82 m2,形成的扩展林隙面积(EG)为60.38 m2。
2.5 林隙形成木的特征
林隙形成木的特征是指林隙形成木的组成结构、径级结构和高度结构,其直接或间接地影响着林隙的环境特征及树种的更新。所以进行阔叶红松混交林林隙形成木特征的研究对全面认识阔叶红松混交林生态系统动态过程具有重要意义。
2.5.1 林隙形成木的组成结构
林隙形成木的组成结构是指形成林隙的树种及其比例。林隙形成木的组成与林隙优势树种的组成密切相关,一般林分中较大的优势木构成形成木的比例大。把在小兴安岭阔叶红松混交林林隙中所调查的41个林隙作为一个总体,林隙形成木的组成统计结果见表6。从表6可以看出,构成林隙形成木的主要树种是红松、白桦、冷杉、枫桦,腐烂程度较严重的,无法判别其树种组成,在表6树种一栏里将其列为“无法确定的树种”。
2.5.2 林隙形成木的径级结构
林隙形成木的径级结构是指形成林隙的各树种在不同径级上的株数分配比例。形成木的大小直接影响着林隙的大小,树木只有达到一定的径级并到达主林层后,才有可能在死亡后形成林隙。以10cm作为一个径级单位(上限排外法),各树种在不同的径级内的株数统计结果见表7。从表7可以看出,各林隙形成木在不同径级中的株数分配比例不同,但它们都以50cm以下居多,占75.98%。调查发现,在小兴安岭阔叶红松混交林林隙内,主林层的主要树种在胸径达到20cm以上(占59.80%)才开始倾倒,20cm以下(占28.92%)的林隙形成木是由于主林层树木倒下时的受害者。由大径级(胸径50cm以上)的树木形成的林隙占(占7.84%)。
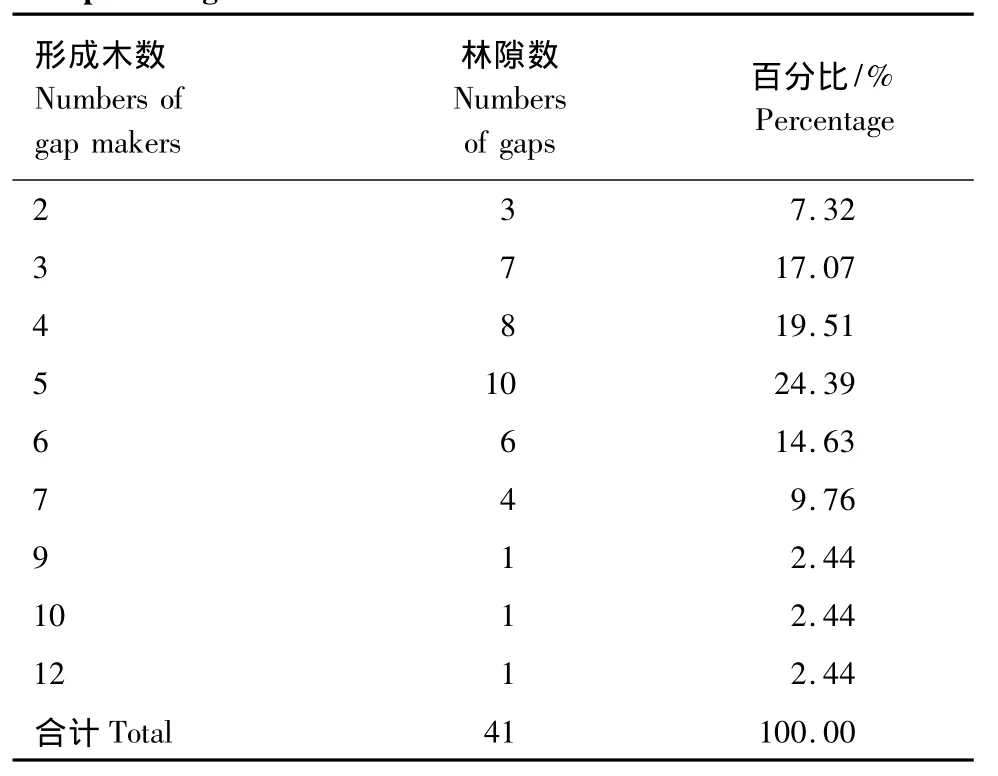
表5 单个林隙形成木的数量及其百分比Table5 Numbers of gap makers in each individual gap and the percentage
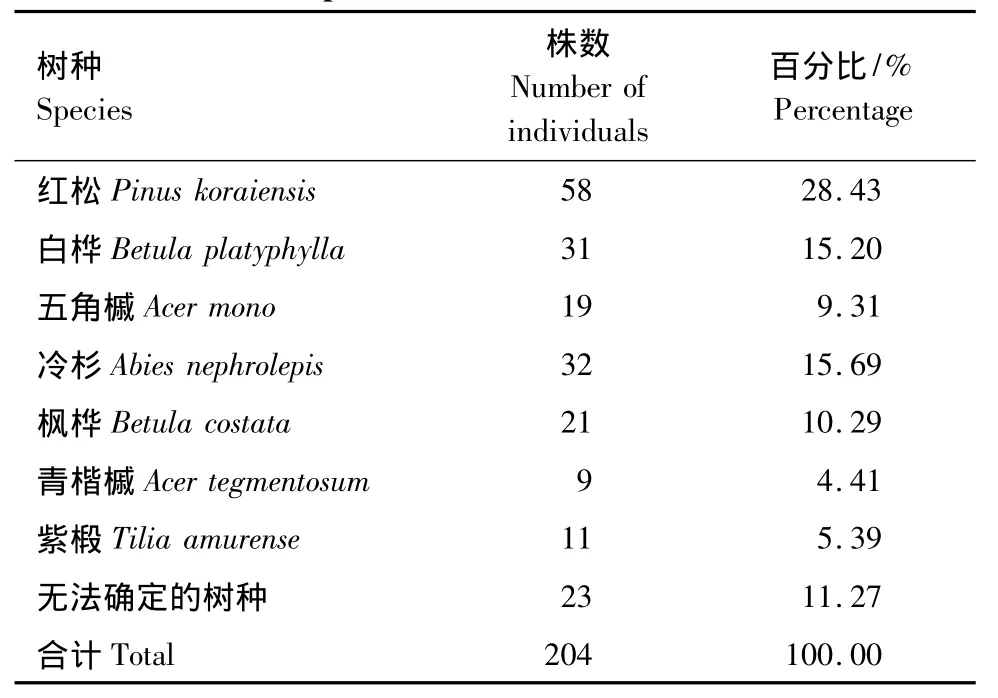
表6 阔叶红松混交林林隙形成木的组成结构Table6 Species composition structure of gap makers in the mixed broad-leaved Korean pine forest
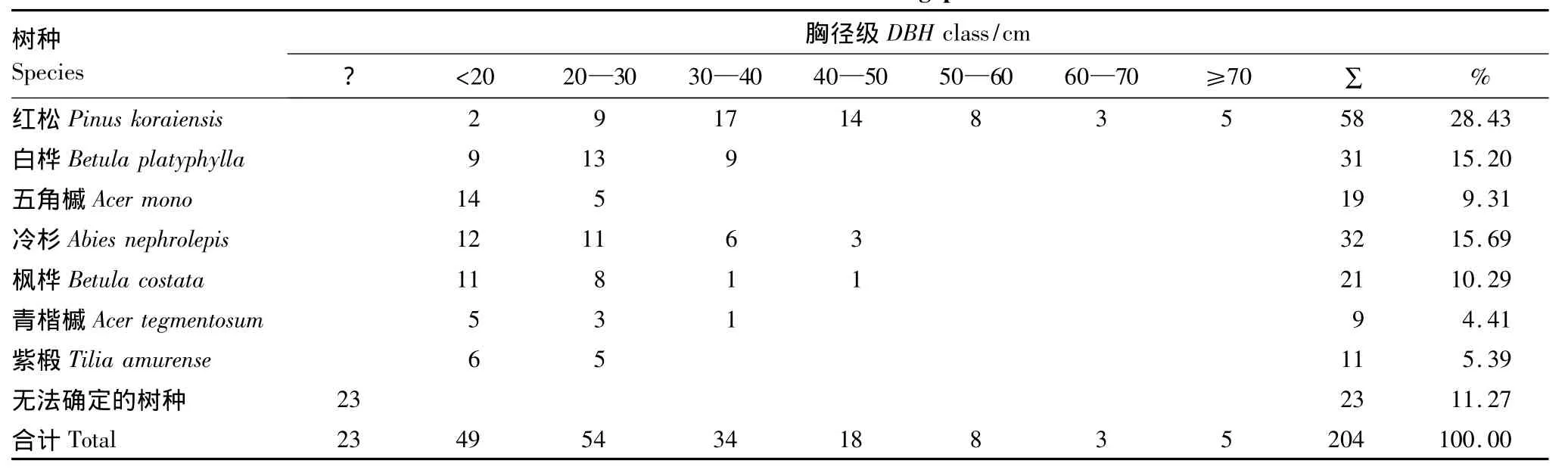
表7 林隙形成木的径级结构Table7 Diameter class structure of gap makers
2.5.3 林隙形成木的高度结构
林隙形成木的高度结构是指形成林隙的各树种在不同高度级内的株数分布情况。将所调查的41个林隙作为总体,统计各树种在各高度级内的株数(表8)。从表8可以看出,树种形成林隙时高度在15—30m以上的占80.39%。一般来说,上层的林隙形成木是林隙的真正制造者,下层部分林隙形成木是由于上层形成木折倒时受机械损伤或小气候的剧烈改变以及竞争中的自然淘汰而引起的。
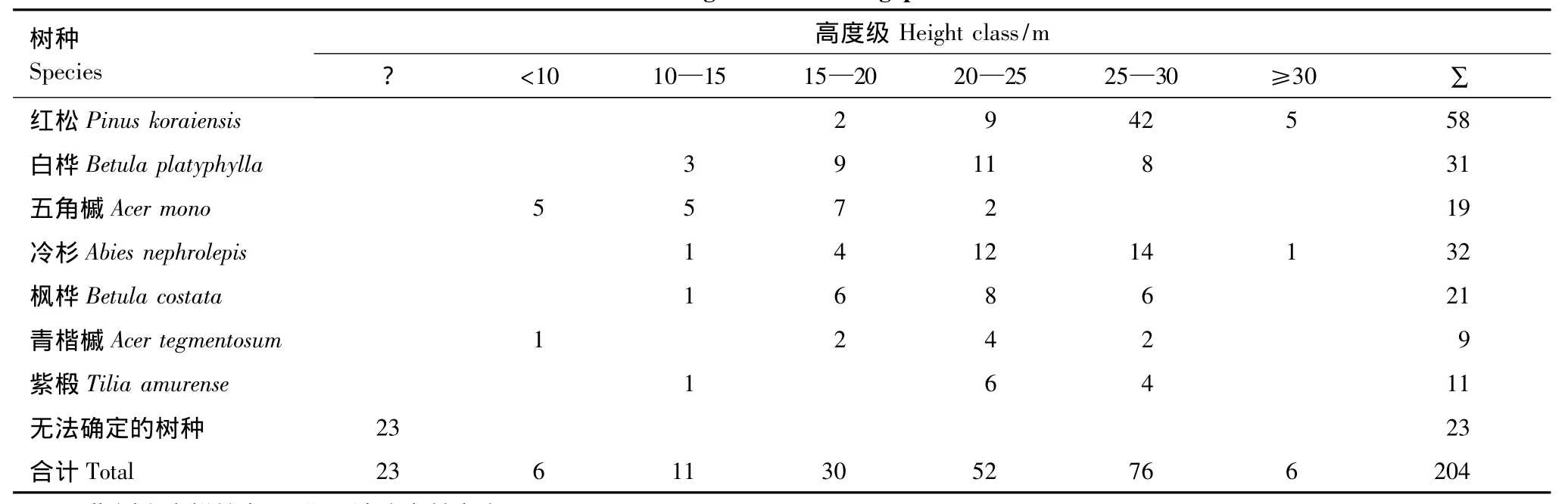
表8 林隙形成木的高度结构Table8 Height structure of gap makers
2.6 林隙边缘木特征
群落中主林层的乔木死亡形成林隙后,其周围的高度达到林冠层的树木称为林隙边缘木(简称GBT),它是衡量林隙特征的一个重要指标。林隙边缘木构成了林隙的大小和形状的边界,影响着林隙的填充方式、物种传播和环境因子的变化[15]。当林隙面积较小时,林隙的填充方式主要以边缘木的侧生长来进行填充,而当林隙面积较大时,林隙内的幼苗能够迅速生长从而可以填充林隙。另外,边缘木高度和林冠厚度或向内冠幅影响着进入到林隙下层的光照,较高的边缘木能维持林隙内部环境的稳定性。
2.6.1 林隙边缘木的径级和高度结构特征
在41个林隙中,共调查到410株边缘木,其中胸径最大的为90.7cm,最小的为10.0cm,平均胸径为32.92cm。根据胸径的测定结果,把胸径分为7个等级(表9),由表9可以看出,各径级所包含的个体数以及占总体的比例从大到小排列为:Ⅱ级>Ⅰ级>Ⅲ级>Ⅳ级>Ⅴ级>Ⅵ级>Ⅶ级。Ⅱ级所占比例最大,为129株,占总体31.46%;胸径>70 cm的个体数有18株,占总体4.39%。

表9 林隙边缘木的径级结构Table9 Diameter class structure of gap border trees
在调查的410株边缘木中,最高的为31.0m,最矮的为8.0m,平均高度为19.6m,与该群落乔木层中主要树木的高度相当。把边缘木高度分为6级(表10),由表10可以看出,15—20m之间的树木数量最多,有148株,占36.10%;其次为20—25m之间的,有113株,占27.56%,≥30m的最少,有19株,占4.63%。
用威布尔分布模型拟合林隙边缘木的胸径级分布。拟合结果采用柯儿莫哥洛夫临界值检验:Dn=0.5834<D0.05,410,林隙边缘木的胸径级多度分布遵循威布尔分布。用正态分布模型拟合其胸径级分布,拟合结果采用卡方值检验:∑X2=70.5827>=0.0000,林隙边缘木的胸径级多度分布不符合正态分布。对林隙边缘木的高度级分布拟合结果为:威布尔分布,Dn=0.5973<D0.05,410,林隙高度级符合威布尔分布;用正态分布拟合结果为∑X2=13.0607>=0.0045,林隙边缘木的高度级多度分布也不符合正态分布(表11)。

表10 林隙边缘木的高度结构Table10 Height structure of gap border trees
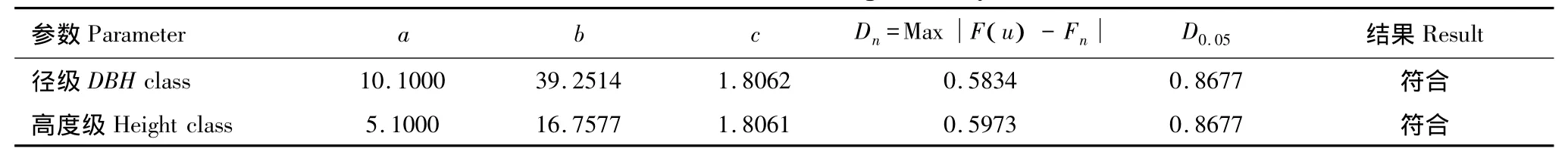
表11 径级和高度级的Weibull分布拟合结果Table 11 The simulated results of DBH class and height class by Weibull distribution model
林隙边缘木胸径和高度的平均值与该群落乔木层主要树木的平均值基本一致,这表明林隙形成木死亡时,其周围大多数树木的高度已达到林冠层,少数树木是在林隙形成后到达林冠层的。随着林隙的不断更新和填充,边缘木的状态也处于动态变化中,对林隙边缘木高度、径级分布的研究,有助于揭示林隙结构与动态和更新所处的状态,了解其微生境的异质性特征。
2.6.2 林隙边缘木的偏冠率分布特征
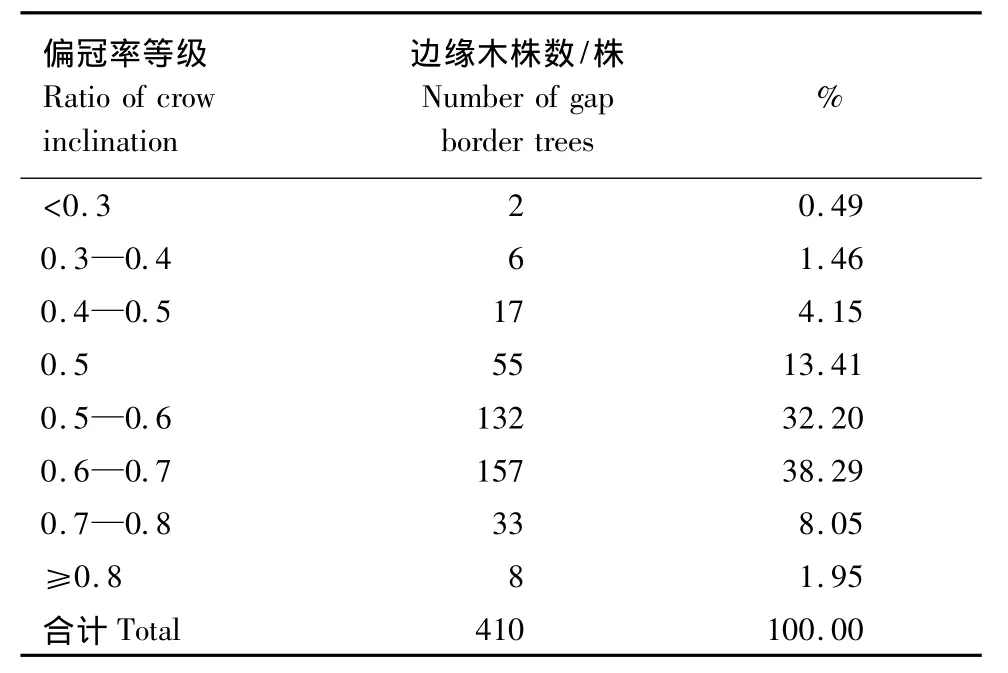
表12 林隙边缘木的偏冠率分布Table 12 Distribution of ratio of crow inclination of gap border trees
林隙形成后,林隙边缘木朝向林隙内的树枝生长速度比与其相反方向的树枝生长速度快,即出现偏冠现象。对41个林隙410株边缘木的偏冠率调查结果表明(表12),有55株(占13.41%)未出现偏冠现象,绝大多数边缘木的偏冠率在0.5—0.7之间(共289株,占70.49%),偏冠率在0.7—0.8的有33株,占8.05%,偏冠率在0.4—0.5的有17株,占4.15%,偏冠率≥0.8和<0.4的株数很少,分别占1.95%和1.95%(表12)。说明林隙的形成导致边缘木出现偏冠,这与对川西亚高山针叶林林隙边缘木和茂兰喀斯特林隙边缘木的研究结果基本一致[15]。林隙中出现偏冠率不均情况的原因是树木两侧都是林隙,先形成林隙的方向利于树枝生长,该侧就会出现偏冠现象。
2.7 林隙形状
林隙形状是林隙重要特征因子,在决定资源可利用性方面起着重要作用。林隙的形状是不规则的。从水平面看,林隙有圆形、椭圆形、多边形和浑圆形等;从垂直剖面看,林隙像一个倒圆锥体,越往上开敞度越大,向下则逐渐变小;填充阶段的林隙表现为不规则的分支脉状,立体观测为不规则的分枝脉状凹陷[21]。林隙形状越不规则或越复杂,边缘上的环境因子变化越显著,同时由于树木对地上(光)与地下(水)的竞争,林隙内的边缘效应就越明显。从图1可知,小兴安岭阔叶红松混交林林隙形状指数变动在0.98—1.82之间,1—1.45之间的分布占56.09%。林隙形状变异较大,多为不规则的多边形,与对天山云杉林林隙特征的研究结果一致[26]。
林冠空隙直径与周围林木算术平均高的比值,表征了林隙相对于周围林分的开敞程度。随着径高比的不同,林隙内的光照、温度、湿度、土壤养分等相应地发生变化。由于林隙径高比对林隙内生境产生影响,从而进一步影响林隙的更新,因此,研究林隙径高比(D/H)具有实际意义[27]。由图2可以看出,冠空隙的直径与高度的比值(用 DCG/H表示)呈单峰型变化,比值在0.30—0.45的分布最多,占36.59%;扩展林隙的直径与高度的比值(用DEG/H表示)呈双峰型变化,比值在0.75—0.90和 1.05—1.15的分布较多,分别占21.95%和24.39%(图3)。根据臧润国等的研究结果,林冠空隙内的光照随D/H比值的增大而增加。当D/H的值大约为2时,光照达到最大[10]。由此可见,该研究区林冠空隙内的光照都没有到达潜在的最大光照值。
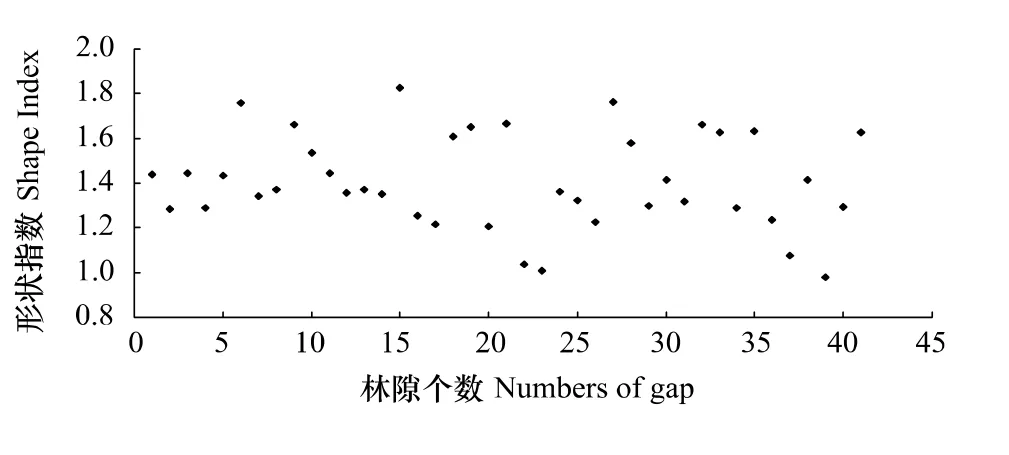
图1 小兴安岭阔叶红松混交林林隙形状指数Fig.1 Shape index of gap in the mixed broad-leaved korean pine forest in Xiaoxing'an Mountains
3 结论与讨论
(1)对小兴安岭阔叶红松混交林林隙的干扰频率进行了计算。结果表明,冠空隙的形成速率为2.28m·km-1·a-1,扩展林隙的形成速率为4.46m·km-1·a-1。林隙干扰的周期为434.8a。长白山红松阔叶林林冠空隙干扰的周转期为751a[12],长白山暗针叶林林隙干扰周期为416.7a[5],天山云杉林干扰返回期间隔为763a[26],在中亚热带常绿阔叶林林隙干扰返回间隔期为357a[13],在塔里木荒漠河岸林林隙的周转率为164a[28],海南岛热带山地雨林林隙干扰的返回间隔期为159a[11]。从以往研究结果来看,从温带到热带林隙干扰间隔期逐步减少,森林循环速率不断增加。
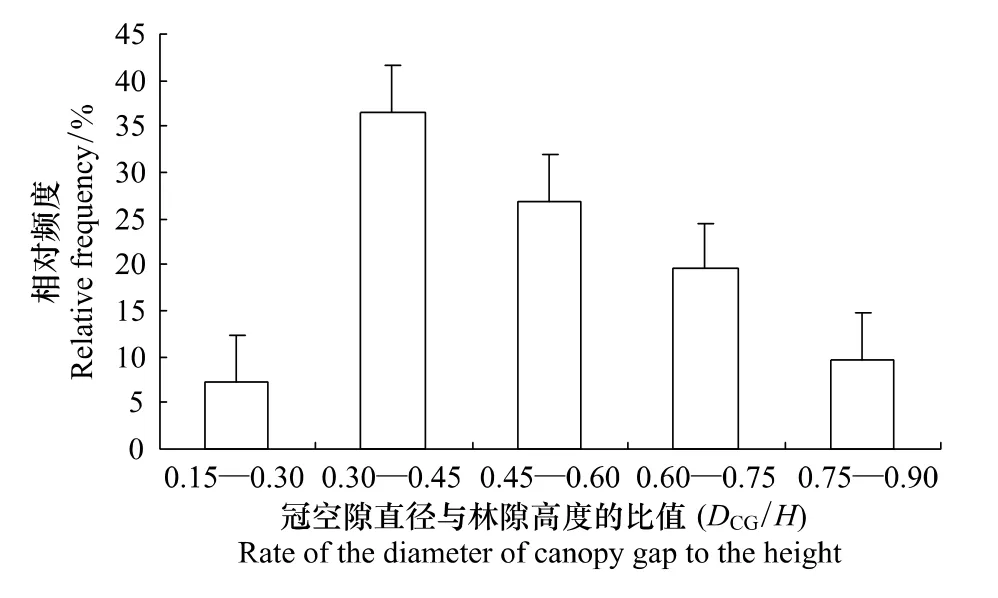
图2 林冠空隙直径与林隙高度比值(D CG/H)的分布Fig.2 Distribution of D CG/H in the mixed broad-leaved korean pine forest
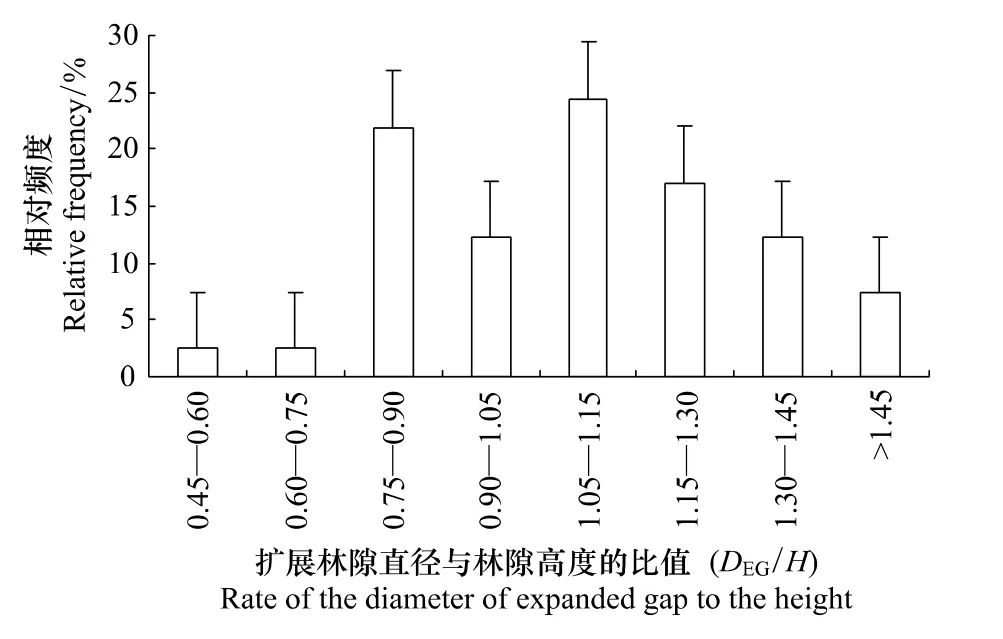
图3 扩展林隙直径与林隙高度的比值(D EG/H)的分布Fig.3 Distribution of D EG/H in the mixed broad-leaved korean pine forest
(2)在所调查的41个林隙中,冠空隙的平均面积为153.37m2,而扩展林隙的平均面积为300.44m2,冠空隙和扩展林隙的相对比例为51.05%。平均每个冠空隙占据22.44 m2,平均每个扩展林隙占据65.95 m2。反映出小兴安岭阔叶红松混交林林隙属中小林隙型,是森林更新演替的必要过程,进一步说明了阔叶红松混交林是比较稳定的森林群落,也反映出林隙的形成速率和林隙填充速率相对平衡。应用Weibull分布模型拟合了阔叶红松混交林林隙大小分布特征。结果表明,冠空隙和扩展林隙面积的大小分布均符合Weibull分布。闫淑君等[17]对中亚热带常绿阔叶林林隙面积进行的Weibull模型拟合结果,与本文的研究结果相一致。赵秀海等[19]采用几种模型对长白山阔叶红松混交林林隙大小结构进行了研究,符利勇等[18]对关帝山天然次生针叶林林隙大小利用3种模型进行了拟合,结果均为对数正态分布模型,与本研究结论不太一致。产生差异的原因可能是不同地域的气候条件、植被组成和结构、林隙干扰类型以及林隙微生境条件等不同,从而使分布模型对林隙大小的拟合精度不同。
(3)小兴安岭阔叶红松混交林林隙是由一种或几种方式共同形成的。主要方式是树干(干中和干基)折断(占56.37%)。在调查中发现,干基折断和掘根风倒的林隙形成木几乎都已发生心腐,以这两种方式形成的林隙处于坡度较大、土壤潮湿或立地条件较差的地段,易受强风影响,与暗针叶林研究结果一致[14]。本研究还发现一些林隙形成木是因为主林层林隙形成木折倒时受到机械损伤及自然淘汰产生的。一般树高和胸径较小的林隙形成木对林隙的生境改善作用较小。这与对长白山暗针叶林林隙干扰的研究结果一致。因此,在以后的调查中,应区分真正的林隙形成木和受害木,使林隙干扰状况的研究更为准确[5]。此外,病虫害、雪压、雷击等因素也有可能形成林隙,其相关性有待于进一步研究。
(4)在小兴安岭阔叶红松混交林中,由5株形成木形成的林隙最多,占24.39%;由3株、4株和6株形成木形成的林隙共占51.21%。可见,绝大多数的林隙是由3—6株形成木所形成的,其他林隙形成木多是上层倒木的受害者。在所调查的41个林隙中,共有204株形成木,平均每个林隙的形成木为4.98株,每株形成木所能形成的冠空隙面积(CG)和扩展林隙面积(EG)分别为30.82 m2和60.38 m2。本文研究结果与其他学者对热带山地雨林[11]、长白山暗针叶林[5]、中亚热带常绿阔叶林[13]、亚高山暗针叶林[14]、天山云杉[26]和荒漠河岸林[28]等林隙研究结果相似之处在于林隙是由多株形成木产生,同时也反映出小兴安岭阔叶红松混交林树种间关系较紧密,一旦有树木发生倾倒,周围树木就会产生倾倒现象,从而形成多株形成木的林隙。
(5)林隙边缘木生长发育过程中高度、冠长和冠形的变化使林隙面积和形状发生变化,增加了林隙微环境的时空异质性。研究结果得出,林隙的边缘木径级分布和高度分布符合Weibull分布,但不符合正态分布。这与杨晓伟等对中亚热带台湾相思次生林林隙边缘木高径分布特征研究结果不一致[20],与闫淑君等在中亚热带常绿阔叶林林隙边缘木高径分布特征的研究结果不尽一致[16]。产生这种差异的原因可能是与区组划分尺度及区组个数有关,也可能与气候条件、植被组成、林隙结构和林隙干扰类型等因子有关,具体原因有待深究。林隙形状指数变动在0.98—1.82之间,1—1.45之间的分布较多。对林隙径高比(D/H)的研究得出,冠空隙的直径与高度的比值呈单峰型变化,比值在0.30—0.45的分布最多;扩展林隙的直径与高度的比值呈双峰型变化,比值在0.75—0.90和1.05—1.15的分布较多,说明此研究区林冠空隙内的光照都没有到达潜在的最大光照值。
(6)林隙形成是推动植被更新替代的重要驱动力,林隙干扰增加了森林环境的异质性,在群落动态和结构维持等方面起着重要的作用。本文对小兴安岭阔叶红松混交林林隙特征及一般干扰状况进行了研究,目的在于为未来深入研究林隙干扰与植被响应关系及森林群落动态奠定基础和积累数据。
[1] Watt A S.Pattern and process in the plant community.Journal of Ecology,1947,35(1/2):1-22.
[2] Runkle JR.Gap regeneration in some old-growth forests of the eastern United States.Ecology,1981,62(4):1041-1051.
[3] Liang X D,Ye W H.Advances in study on forest gaps.Journal of Tropical and Subtropical Botany,2001,9(4):355-364.
[4] Jiang G H,Wu Z M,Shen H H,Zhang L.Formation and characteristics of gaps in Castanopsis eyrei stands in Zhawan Natural Reserve of Anhui Province.Journal of Anhui Agricultural University,2008,35(3):385-389.
[5] Yang X.Gap characteristics and disturbance regime in a dark coniferous forest in Changbai mountain areas.Acta Ecologica Sinica,2002,22(11):1825-1831.
[6] Gagnon T L,Jokela E J,Moser W K,Huber D A.Characteristics of gaps and natural regeneration in mature longleaf pine flatwoods ecosystems.Forest Ecology and Management,2004,187(2/3):373-380.
[7] de Lima R A F,de Moura L C.Gap disturbance regime and composition in the Atlantic Montane rain forest:the influence of topography.Plant Ecology,2008,197(2):239-253.
[8] Kakamess R,Zobel Z.The role of the seed bank in gap regeneration in a calcareous grassland community.Ecology,2002,83(4):1017-1025.
[9] Jin G Z,Tian Y Y,Zhao F X,Kim J H.The Pattern of natural regeneration by canopy gap size in the mixed broadleaved-korean pine forest of Xiaoxing'an mountains,China.Journal of Korean Forestry Society,2007,96(2):227-234.
[10] Zang R G,Liu T,Guo Z L,Gao W T.Gap disturbance regime in a broadleaved Korean pine forest in the Changbai Mountain Natural Reserve.Acta Phytoecologica Sinica,1998,22(2):135-142.
[11] Zang R G,Yang Y C,Liu J Y,Yu S X,Yang X S.Gaps and their natural disturbance regimes in the tropical mountain rain forest of Hainanisland.Scientia Silvae Sinicae,1999,35(l):1-8.
[12] Wu G.Characteristics of gap in Korean pine broad-leaved forests in Changbai Mountain.Chinese Journal of Applied Ecology,1997,8(4):360-364.
[13] Yan SJ,Hong W,Wu C Z,Bi X L,Wang X G,Feng L.Gaps and their natural disturbance characteristics in mid-subtropical evergreen broadleaved forest in Wanmulin.Chinese Journal of Applied Ecology,2004,15(7):1126-1130.
[14] Wang W,Tao J P,Li Z F,Zhang W Y,Ding Y.Gap features of subalpine dark coniferous forest in Wolong Nature Reserve.Chinese Journal of Applied Ecology,2004,15(11):1989-1993.
[15] Xian JR,Hu T X,Wang K Y,Zhang Y B,Zhu X M,Xu JW.Studies on the characteristics of gap border trees in the Subalpine coniferous forest in Western Sichuan.Forest Research,2004,17(5):636-640.
[16] Yan SJ,Hong W,Wu C Z.Distribution characteristic of DBH and tree height for gap edge-trees in mid-subtropical evergreen broad-leaved forest in Wanmulin.Journal of Fujian College of Forestry,2003,23(1):5-8.
[17] Yan S J,Hong W,Wu C Z.A study on Weibull model for GAP area in mid-subtropical evergreen broad-leaved forest in Wanmulin.Acta Agriculturae Universitatis Jiangxiensis:Natural Sciences Edition,2002,24(6):802-805.
[18] Fu L Y,He Z,Liu Y A.Study of the gap size model in a secondary coniferous forest of Guandi Mountain.Journal of Nanjing Forestry University:Natural Science Edition,2010,34(5):51-54.
[19] Zhang CY,Zhao X H,Zheng JM.Size structure of canopy gaps in broadleaved Korean pine forests in the Changbai Mountains.Journal of Beijing Forestry University,2006,28(4):34-38.
[20] Yang X W,Hong W,Wu C Z.Distribution characteristic of DBH and tree height for gap edge trees of acacia confuse in Mid-subtropical Zone.Journal of Southwest Forestry College,2010,30(6):25-28.
[21] Zhu J J,Liu SR.Ecological Research on Forest Disturbances.Beijing:Forestry Press of China,2007:166-182.
[22] Wu C Z,Hong W.Study on Weibull Model for species abundance distribution of Tsoongiodendron odorum forest.Journal of Fujian College of Forestry,1997,17(1):20-24.
[23] Bi X L,Hong W,Wu C Z,Yan SJ.The application of modified simplex method in the study on stand structure.Acta Agriculturae Universitatis Jiangxiensis,2002,24(1):95-99.
[24] Ding ST.The Weibull distribution and fitting.Acta Agriculturae Universitatis Jiangxiensis,1985,24(3):45-52.
[25] Wu CZ,Hong W.A Study on Weibull D.B.H.distribution of Chinese fir plantation and its optimal fitting.Acta Agriculturae Universitatis Jiangxiensis,1998,20(1):86-90.
[26] Liu C L,Pan C D,Shi R F.Characteristics of gap in Picea schrenkiana-Dryopteris filix-mas stand.Arid Land Geography,2007,30(2):209-216.
[27] Fu LY,Tang SZ,Liu Y A.The forest gap diameter height ratio in a secondary coniferous forest of Guan Di Mountain.Acta Ecologica Sinica,2011,31(5):1260-1268.
[28] Han L,Wang H Z,Chen JL,Yu J.Disturbance regimes and gaps characteristicsof the desert riparian forest at the middle reaches of Tarim River.Acta Ecologica Sinica,2011,31(16):4699-4708.
参考文献:
[3] 梁晓东,叶万辉.林窗研究进展(综述).热带亚热带植物学报,2001,9(4):355-364.
[4] 江国华,吴泽民,沈后海,张磊.安徽查湾自然保护区甜槠林林隙形成及特征研究.安徽农业大学学报,2008,35(3):385-389.
[5] 杨修.长白山暗针叶林林隙一般特征及干扰状况.生态学报,2002,22(11):1825-1831.
[10] 臧润国,刘涛,郭忠凌,高文韬.长白山自然保护区阔叶红松混交林林隙干扰状况的研究.植物生态学报,1998,22(2):135-142.
[11] 臧润国,杨彦承,刘静艳,余世孝,杨秀森.海南岛热带山地雨林林隙及其自然干扰特征.林业科学,1999,35(l):1-8.
[12] 吴刚.长白山红松阔叶林林冠空隙特征的研究.应用生态学报,1997,8(4):360-364.
[13] 闫淑君,洪伟,吴承祯,毕晓丽,王新功,封磊.中亚热带常绿阔叶林林隙及其自然干扰特征的研究.应用生态学报,2004,15(7):1126-1130.
[14] 王微,陶建平,李宗峰,张炜银,丁易.卧龙自然保护区亚高山暗针叶林林隙特征研究.应用生态学报,2004,15(11):1989-1993.
[15] 鲜骏仁,胡庭兴,王开运,张远彬,朱雪梅,徐精文.川西亚高山针叶林林窗边界木特征的研究.林业科学研究,2004,17(5):636-640.
[16] 闫淑君,洪伟,吴承祯.中亚热带常绿阔叶林林隙边缘木高径分布特征.福建林学院学报,2003,23(1):5-8.
[17] 闫淑君,洪伟,吴承祯.中亚热带常绿阔叶林林隙面积的Weibull分布模型研究.江西农业大学学报:自然科学版,2002,24(6):802-805.
[18] 符利勇,何铮,刘应安.关帝山天然次生针叶林林隙大小模型研究.南京林业大学学报:自然科学版,2010,34(5):51-54.
[19] 张春雨,赵秀海,郑景明.长白山阔叶红松混交林林隙大小结构研究.北京林业大学学报,2006,28(4):34-38.
[20] 杨晓伟,洪伟,吴承祯.中亚热带台湾相思次生林林隙边缘木高径分布特征.西南林学院学报,2010,30(6):25-28.
[21] 朱教君,刘世荣.森林干扰生态研究.北京:中国林业出版社,2007:166-182.
[22] 吴承祯,洪伟.观光木群落物种多度分布的Weibull模型研究.福建林学院学报,1997,17(1):20-24.
[23] 毕晓丽,洪伟,吴承祯,闫淑君.改进单纯形法在林分结构规律研究中的应用.江西农业大学学报,2002,24(1):95-99.
[24] 丁思统.威布尔(Weibull)分布及其拟合.江西农业大学学报,1985,24(3):45-52.
[25] 吴承祯,洪伟.杉木人工林胸径的Weibull分布及其最优拟合研究.江西农业大学学报,1998,20(1):86-90.
[26] 刘翠玲,潘存德,师瑞峰.鳞毛蕨天山云杉林林冠空隙特征.干旱区地理,2007,30(2):209-216.
[27] 符利勇,唐守正,刘应安.关帝山天然次生针叶林林隙径高比.生态学报,2011,31(5):1260-1268.
[28] 韩路,王海珍,陈加利,于军.塔里木荒漠河岸林干扰状况与林隙特征.生态学报,2011,31(16):4699-4708.
