甘蓝一代杂种S单元型的分布
2013-09-03苗雯雯刘玉梅杨丽梅张扬勇方智远
苗雯雯 田 磊 庄 木 刘玉梅 杨丽梅 张扬勇 方智远
(中国农业科学院蔬菜花卉研究所, 北京 100081)
当前,甘蓝(Brassica oleraceaL. var.capitataL.)的商业栽培品种中几乎都是利用自交不亲和系或雄性不育系配制的一代杂种。在甘蓝育种实践中,一代杂种的结实性对新品种的示范推广进程具有重要的影响,且与亲本材料的自交不亲和性(self-incompatibility,SI)存在着密切的联系。甘蓝属于典型的孢子体型自交不亲和体系,在遗传上由一个具有多个复等位基因的S位点控制,又称S单元型(S haplotype)(Nasrallah & Nasrallah,1993)。目前已鉴定的涉及自交不亲和识别反应的重要S基因包括:S位点糖蛋白基因(S-locus glycoprotein,SLG)、S位点受体激酶基因(S-locus receptor kinase,SRK)、S位点富半胱氨酸蛋白基因(S-locus cysteinerich protein,SCR)或S位点蛋白11基因((S-locus protein 11,SP11),其中SRK是雌蕊柱头中表达的雌性决定因子(Kusaba et al.,2000),而SCR/SP11是花粉中的雄性决定因子(Schopfer et al.,1999 )。在甘蓝类作物(包括结球甘蓝、羽衣甘蓝、抱子甘蓝、球茎甘蓝、花椰菜等)中,前人研究已鉴定出存在50多个S单元型(Ockendon,2000)。明确甘蓝一代杂种中S单元型,有助于了解育成品种中优势S单元型的分布,并可为引入新的S单元型以及配制新组合提供参考依据。因此,在田磊等(2011)的研究基础上,本试验利用一套特异引物对SRK基因进行PCR扩增、克隆测序及Blast分析序列信息,鉴定了23份甘蓝一代杂种的S单元型,为甘蓝新品种选育提供参考。
1 材料与方法
1.1 试验材料
供试材料为中国农业科学院蔬菜花卉研究所育成的23份甘蓝一代杂种,包括京丰、庆丰等20个通过全国(或北京市)审定(或鉴定)的不同类型的商业品种,以及06-28、06-88等4个新组合(表1)。所有材料均于2012年在本所科研基地种植,并进行统一的栽培管理。
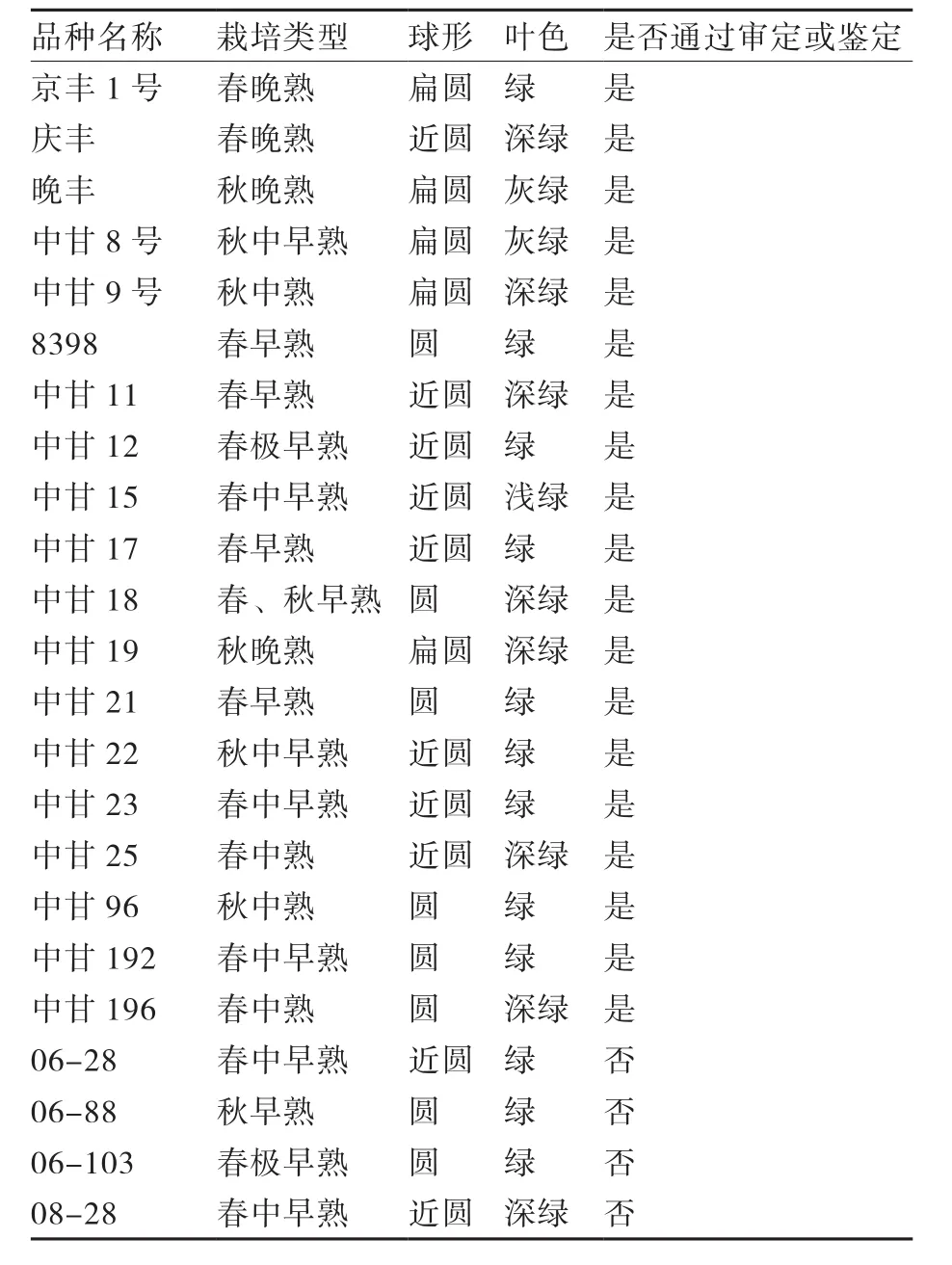
表1 供试甘蓝一代杂种的特征特性
1.2 试验方法
1.2.1 基因组DNA的提取 取适量(约2 g)供试材料幼嫩的叶片,液氮研磨后利用改良的CTAB法(Doyle & Doyle,1990)提取DNA。用1%琼脂糖凝胶电泳检测DNA浓度和纯度;根据浓度大小分别稀释到50 ng·μL-1,作为PCR扩增的模板。
1.2.2 基因组DNA的PCR扩增SRK基因激酶区的DNA片段由Ⅰ类特异引物PK1/PK4(Nishio et al.,1997)和Ⅱ类特异引物KD4/KD7(Park et al.,2002)进行扩增;若这2对引物均无目标扩增片段,则利用SRK基因激酶区的通用引物DF/DR进行扩增(表2)。Ⅰ类特异引物PK1/PK4扩增片段为SRK基因激酶区2~5外显子序列,Ⅱ类特异引物KD4/KD7扩增片段为SRK基因激酶区4~7外显子序列,引物DF/DR的扩增片段为SRK基因激酶区5~7外显子序列。
以基因组DNA为模板,利用以上3对引物进行PCR扩增。PK1/PK4的PCR扩增程序为:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火1 min,72 ℃延伸1.5 min,35个循环;72 ℃延伸10 min,4 ℃保存。KD4/KD7的PCR扩增程序为:94 ℃预变性5 min;94 ℃变性30 s,60℃退火1 min,72 ℃延伸1.5 min,34个循环;72 ℃延伸10 min,4 ℃保存。DF/DR的PCR扩增程序为:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸1 min,35个循环;72℃延伸7 min,4 ℃保存。PCR扩增产物在1%琼脂糖凝胶中进行电泳,恒压130 V。利用紫外凝胶成像仪系统进行拍照。PCR反应体积为20 μL:模板DNA 4.0 μL,dNTPs(2.5 mmol·L-1)1.6 μL,10×Buffer( 含 Mg2+)2.0 μL,Taq酶(2.5 U·μL-1)0.4 μL, 上 游 引 物(5 mmol·L-1)1.0 μL,下游引物(5 mmol·L-1)1.0μL,ddH2O 10μL。
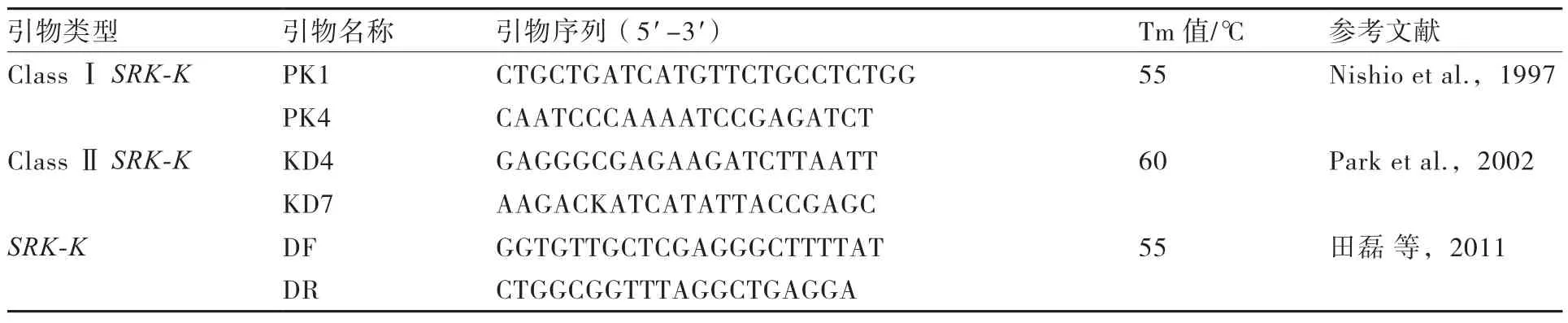
表2 SRK基因激酶区特异引物
1.2.3 目的片段的回收克隆及序列分析 利用EasyPure Quick Gel Extraction Kit(北京全 式金生物技术有限公司)回收目标片段,连接到pGEM-T Easy(Promega公司)载体上,4 ℃过夜。连接产物利用Trans1-T1 Phage Resistant感受态细胞(北京全式金生物技术有限公司)转化,进行蓝白斑筛选。挑取阳性克隆进行PCR扩增鉴定,每个目标片段的转化样品选3个阳性克隆由生工生物工程(上海)有限公司测序。利用DNAStar软件进行序列拼接后,Blast以及BlastX在NCBI(http:// www.ncbi.nlm.nih.gov/)在线进行,比对的Organism为Brassica oleracea(taxid:3712)。
2 结果与分析
2.1 甘蓝一代杂种S单元型的鉴定
SRK激酶区引物PK1/PK4的扩增片段大小约为950 bp(图1),引物KD4/KD7的扩增片段大小约为1 100 bp(图2)。PK1/PK4仅能扩增Ⅰ类甘蓝材料,在18份材料中有目标片段的扩增;KD4/KD7仅能扩增Ⅱ类甘蓝材料,在15份材料中有目标片段的扩增。因此,在23份供试甘蓝一代杂种中,初步推断庆丰、中甘15、8398等10份材料属于Ⅰ类和Ⅱ类杂合体,京丰1号、中甘8号、中甘9号等8 份材料属于Ⅰ类杂合体,中甘11、中甘12、中甘22等5份材料属于Ⅱ类杂合体。

图1 特异引物PK1/PK4的PCR扩增结果
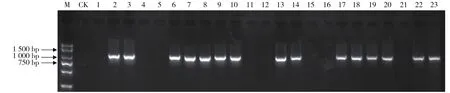
图2 特异引物KD4/KD7的PCR扩增结果
对于属Ⅰ类和Ⅱ类杂合体的庆丰、中甘15、8398等,利用以上两对引物分别扩增出的激酶区片段,进行回收、克隆及测序分析确定其S单元型。中甘15、中甘17、中甘21和8398的母本具有相同的来源,它们的PK1/PK4扩增片段克隆测序结果完全相同(918 bp),Blast比对表明,与Brassica oleracea SRK28基因(GenBank:AB0190355.1)在序列覆盖率为100%下核苷酸序列相似性为100%(E值为0)。因此,这4份甘蓝一代杂种均含有S28单元型。8398和中甘21的父本虽然来源不同,但它们的KD4/KD7扩增片段克隆测序结果完全相同(1 043 bp),BlastX比对表明,可能的氨基酸序列均与Brassica oleracea SRK5(GenBank:CAB41878.1)的氨基酸序列相似性为99%(序列覆盖率98%,E值为0),仅415~417 bp处的氨基酸F(苯丙氨酸)与SRK5蛋白的氨基酸L(亮氨酸)不同,二者均属于非极性疏水氨基酸。因此,推测8398和中甘21含有S5单元型。采用同样的方法,鉴定了其余属Ⅰ类和Ⅱ类杂合体的甘蓝品种的S单元型(表3)。其中,庆丰的PK1/PK4扩增片段克隆测序未能确定其具体的Ⅰ类S单元型,将其暂命名为Sn。
本试验中,供试甘蓝一代杂种的亲本材料均为高代自交系或雄性不育系,理论上是同一S单元型的纯合体,因此,杂种的S单元型同时含有双亲的S单元型。以上推测在属Ⅰ类和Ⅱ类杂合体的庆丰、中甘15、8398等S单元型测定中得到验证。对于同属Ⅰ类(或Ⅱ类)杂合类型的材料,扩增条带为单一条带,少量的克隆测序难以区分其中含有不同的S单元型,在未知S单元型的情况下设计仅扩增杂合体中的每个Ⅰ类S单元型的特异引物比较困难。因此Ⅰ类杂合体的京丰1号、中甘8号等,以及属于Ⅱ类杂合体的中甘11、中甘12等材料,通过对其父、母本纯合体的S单元型进行鉴定,推测配制的一代杂种的S单元型(表3)。
晚丰仅在KD4/KD7扩增得到目标条带,起初推测其为Ⅱ类杂合体。利用其父母本鉴定其S单元型时,首先确定其中一个亲本为S2,而另一个亲本在PK1/PK4和KD4/KD7中均没有扩增产物,由此推测可能是该亲本的S单元型中SRK基因的激酶区不能用以上两对引物扩增得到。因此,利用在Ⅰ类和Ⅱ类S单元型中均有扩增产物的引物DF/DR进行扩增(图3),扩增片段大小为750 bp,测序分析鉴定该材料的S单元型为S68,因此属Ⅰ类和Ⅱ类杂合类型(S68S2)。

表3 23个甘蓝一代杂种S单元型的鉴定结果
2.2 S单元型在甘蓝一代杂种中的分布
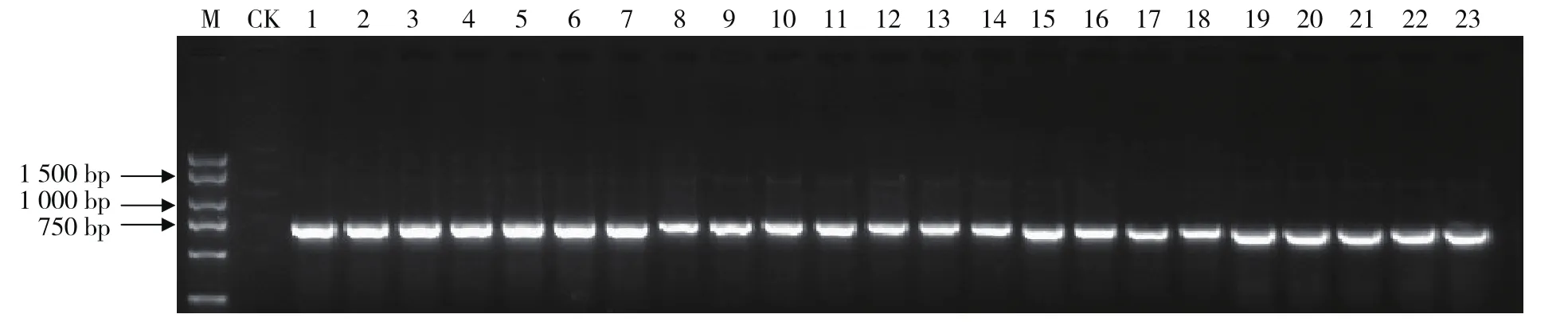
图3 利用特异引物DF/DR对甘蓝一代杂种的PCR扩增
通过序列分析鉴定S单元型,供试23个甘蓝一代杂种中有8个Ⅰ类S单元型S7、S16、S28、S35、S45、S57、S68和未知的Sn,Ⅱ类S单元型有S2、S5、S15。其中Ⅰ类和Ⅱ类杂合体所占的比例最大,约占47.8%;其次为Ⅰ类杂合体,约占34.8%;而Ⅱ类杂合体,约占17.4%。S单元型中出现频率最高的是Ⅱ类的S5,达到19.6%;其次为Ⅰ类的S28,约占17.4%。由此可知,Ⅱ类S5单元型是组成供试甘蓝一代杂种的主要S单元型,这可能与骨干亲本的应用频率较高有关。总体来说,与甘蓝类作物已鉴定的50多个S单元型相比,供试品种中出现的S单元型的数目不多,为了避免配制的杂交组合出现杂交不结实现象,在今后的甘蓝遗传育种中引进和利用更多的S单元型是十分重要的。
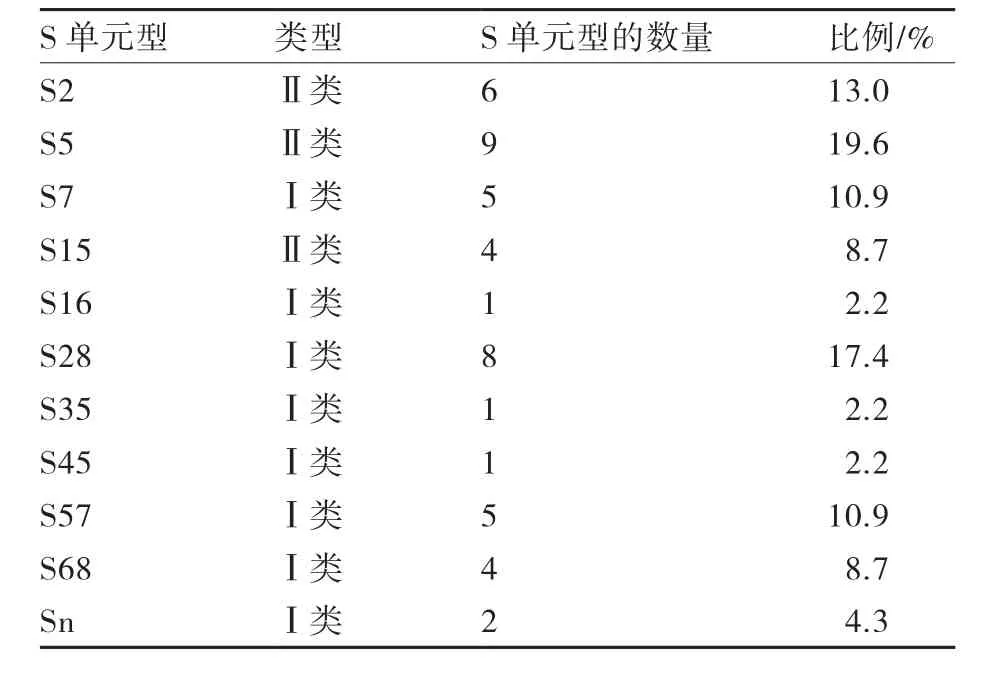
表4 23个甘蓝一代杂种S单元型的分布
3 结论与讨论
在本试验中,根据庆丰的PK1/PK4扩增片段序列未能确定该品种的Ⅰ类S单元型(序列已登录到GenBank:JX840774),表明该S单元型(暂命名为Sn)可能是已鉴定的S单元型中未公开序列的,也可能是甘蓝中尚未被鉴定的新S单元型,进一步测定其SRK的S区、SCR/SP11、SLG等基因序列的分析鉴定工作正在进行中。
本试验中,大部分甘蓝一代杂种的亲本为自交不亲和系,如中甘18,其亲本为S28和S57,都属于Ⅰ类S单元型,表现为强自交不亲和性;而有的亲本材料则为自交亲和系,如中甘21的父本,属于Ⅱ类的S5单元型,表现为强自交亲和性。以上结果验证了Ⅰ类S单元型具有较强的SI性,而Ⅱ类S单元型的SI性相对较弱,与Ockendon(1982)提出的甘蓝中S5和S15纯合体的自交不亲和性更弱,更容易出现自交亲和材料的结论基本一致。
在选育甘蓝自交系时,需要经过自交分离以及定向选择的过程,自交多代后才能获得具有稳定的、优良性状的自交系,这就容易造成一些S单元型的丢失。方智远等(1983)利用花期杂交授粉的方法研究金早生、黑叶小平头等15份自交不亲和系间的亲和关系,指出这15份材料中涉及到13个不同的S单元型。Ockendon(1982)利用荧光显微法对197份甘蓝类材料(包括紫甘蓝、皱叶甘蓝)的S单元型进行鉴定,共发现了31个S单元型,其中在甘蓝中发现了21个S单元型。Sakamoto等(2000)对31个甘蓝一代杂种的SLG基因进行PCR-RFLP分析,鉴定出15个S单元型,但比Ockendon(1982)多了4个新的S单元型(S33、S35、S51、S57)。在甘蓝类作物中已鉴定出的S单元型有50多个,而甘蓝栽培品种中仅约占一半,表明在甘蓝栽培品种的选育过程中S单元型的丢失十分严重。本试验中甘蓝一代杂种中涉及的S单元型仅有11个,可能与在培育新品种时利用骨干亲本的频率比其他材料的频率高有关。育种材料中S单元型类型少,会增加配制杂种的亲本间杂交不亲和的发生频率。06-88是由两个来源不同的优良高代自交不亲和系配制而成的优良品种,但在利用蜜蜂繁殖一代杂种时结实率很低,致使繁种不成功。本试验中S单元型鉴定结果揭示,这是由于双亲S单元型相同引起的严重杂交不亲和所致。因此,开展甘蓝材料的S单元型鉴定研究,并在今后的甘蓝遗传育种中引进和利用更多的S单元型是十分重要的。
方智远,孙培田,刘玉梅.1983.甘蓝杂种优势利用和自交不亲和系选育的几个问题.中国农业科学,(3):51-62.
田磊,庄木,刘玉梅,杨丽梅,张扬勇,方智远.2011.两个甘蓝自交系自交不亲和S单元型的初步鉴定.中国蔬菜,(4):8-12.
Doyle J J,Doyle J L.1990.Isolation of plant DNA from fresh tissue.Focus,12:13-15.
Kusaba M,Matsushita M,Okazaki K,Satta Y,Nishio T.2000.Sequence and structural diversity of the S locus genes from different lines with the same self-recognition specificities inBrassica oleracea.Genetics,154:413-420.
Nasrallah J B,Nasrallah M E.1993.Pollen-stigma signalling in the sporophytic self-incompatibility response.Plant Cell,5:1325-1335.
Nishio T,Kusaba M,Sakamoto K,Ockendon D J.1997.Polymorphism of the kinase domain of the S-locus receptor kinase gene (SRK)inBrassica oleraceaL. Theoretical and Applied Genetics,95:335-342.
Ockendon D J.1982.An S-allele survey of cabbage (Brassica oleraceavar.capitata).Euphytica,31:325-331.
Ockendon D J.2000.The S-allele collection ofBrassica oleracea.Acta Horticultural,539:25-30.
Park J I,Lee S S,Watanabe M,Takahata Y,Nou I S.2002.Identification of S-alleles using polymerase chain reaction-cleaved amplified polymorphic sequence of the S-locus receptor kinase in inbreeding lines ofBrassica oleracea.Plant Breeding,121:192-197.
Sakamoto K,Kusaba M,Nishio T.2000.Single-seed PCR-RFLP analysis for the identification of S haplotypes in commercial F1hybrid cultivars of broccoli and cabbage.Plant Cell Reports,19:400-406.
Schopfer C R,Nasrallah M E,Nasrallah J B.1999.The male determinant of self-incompatibility inBrassica.Science,286:1697-1700.
