南昌市6种棕榈科植物低温胁迫的生理响应
2013-08-27徐维杰杨寅桂喻苏琴
徐维杰,袁 婷,杨寅桂,喻苏琴
(1.江西农业大学 园林与艺术学院,江西 南昌 330045;2.江西农业大学 农学院,江西 南昌 330045)
棕榈科(Palmae)植物是热带地区重要的代表植物,目前全世界共存210余属2 800~3 000种,大多分布于热带及亚热带地区,以海岛及滨海热带雨林为主,是热带亚热带景观的基本标志之一[1]。棕榈科植物树形独特,树姿优美,在我国南方城市园林绿化中起着重要的作用。近年来,上海、四川等地通过引种驯化,在园林应用中收到了较好的效果[2-6]。南昌也在园林中应用棕榈科植物[7],并有进一步发展升温的趋势,但在实际应用中还存在一定的盲目性和盲从性,前几年南昌地区部分住宅区栽植的棕榈科植物,在2008年的低温寒害中部分冻死[8],寒害和冻害已成为长江流域棕榈科植物引种的重要限制因子[9-11],引起了人们的高度重视,一些学者对棕榈科植物的抗寒生理以及抗寒机制等方面展开了一些研究[12-19],旨在探索解决低温危害的根本方法。本研究以南昌市园林中栽培应用的6种棕榈科植物:蒲葵(Livistona chinensis)、华盛顿棕(Washingtonia robusta)、加拿利海枣(Phoenix canariensis)、银海枣(Phoenix sylvestris)、布迪椰子(Butia capitata)、美丽针葵(Phoenix roebelnii)为材料,研究其在低温胁迫下相对电导率、过氧化物酶(POD)、丙二醛(MDA)等生理生化指标变化,并配合Logistic方程求其半致死温度(LT50)[20],了解它们的抗寒能力,为“南棕北移”尤其是棕榈科植物在长江流域的应用提供参考。
1 材料与方法
1.1 南昌地区气候特点
南昌位于东经 115°38'~116°03',北纬 28°35'~28°55',属亚热带季风性气候,湿润温和,日照充足。年平均气温在17.1℃ ~17.8℃,极端最高气温达40.9℃,极端最低气温低于-10℃。年降雨量1 600~1 700 mm。年平均降雪日数8~10 d,积雪日数4~5 d,最大积雪深度20~40 cm。年无霜期251~272 d。冬季多偏北风,夏季多偏南风。
1.2 试验材料
蒲葵、加拿利海枣、银海枣和华盛顿棕采自南昌市蒋巷镇国鸿集团生态园,2003年栽植,树龄约10年;美丽针葵采自南昌市皇冠国际小区,2008年栽植,树龄约20年;布迪椰子采自江西农业大学花卉基地,2005年栽植,树龄约8年。
本试验采用叶片离体人工低温处理方法。2009年11月上旬采集叶片,选取植株中部叶片,每树种剪取3片叶,采后立即带回实验室,每树种剪取20片小裂片装入密封的塑料袋中,置于LRH-250CA型低温培养箱中进行低温处理,试验处理设置5个温度梯度,分别为:0,-3,-6,-9,-12℃,以南昌地区11月上旬平均气温15℃为对照温度,每两个温度间降温1 h,降至所需的温度后处理12 h,然后取出部分材料放置于室温下解冻12 h,进行各种生理指标的测定,剩下的材料继续下一梯度处理。
1.3 试验方法
1.3.1 相对电导率及半致死温度测定 参照邹琦[21]的方法测定相对电导率。结合Logistic方程对不同温度和相对电导率之间的关系进行拟合,求出曲线的拐点温度作为半致死温度[17]。
1.3.2 POD酶活性的测定 采用愈创木酚法[22]测定过氧化物酶(POD)活性。
1.3.3 MDA含量的测定 按高俊凤的方法[23]测定丙二醛(MDA)含量。
1.4 数据处理
采用Excel 2003对原始数据进行整理,应用SPSS 13.0统计分析软件对数据进行分析。
2 结果与分析
2.1 低温胁迫对相对电导率的影响
经低温处理后6种棕榈科植物相对电导率的变化趋势大体相同,均随温度降低呈现增大趋势(图1)。从对照温度到0℃降温过程中,各种棕榈的相对电导率都有所上升,但变化幅度不大。随着温度继续下降,各种棕榈的相对电导率变化有较大差异。0~-3℃,美丽针葵的相对电导率上升幅度最大,由16.71%上升至61.14%, 上升了44.43%;-6 ℃时,蒲葵、银海枣、华盛顿棕的相对电导率明显上升;-6~-9℃,布迪椰子与加拿利海枣的相对电导率急剧升高;-12℃时各种棕榈的相对电导率达到最大值,其中相对电导率增长幅度最大的是美丽针葵,蒲葵次之,增长幅度最小的是布迪椰子。
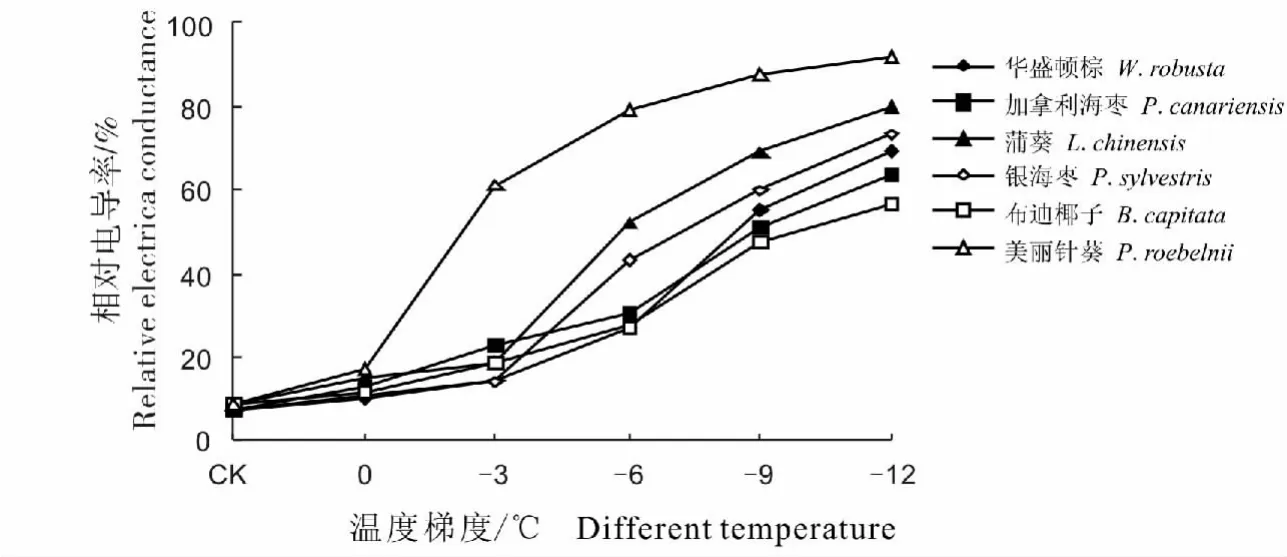
图1 不同低温处理对6种棕榈科植物相对电导率的影响Fig.1 The effect of relative conductivity on six palm plants under low temperature treatment
根据6种棕榈科植物相对电导率的变化趋势,再配合Logistic方程,求出各棕榈科植物的低温半致死温度(LT50)值。几种棕榈科植物经不同低温处理后,相对电导率符合Logistic函数,方程具有较高的拟合度,并达到极显著差异水平(表1)。其中布迪椰子的 LT50是6种棕榈科植物中最低的,可达-10.4℃,说明它的抗寒性好于其余5种;美丽针葵的LT50为-2.3℃,在6种棕榈科植物中最不耐寒。

表1 6种棕榈科植物相对电导率的Logistic方程及半致死温度LT50 Tab.1 The parameters of Logistic and LT50of six palm plants
2.2 低温胁迫对POD酶活性的影响
经低温处理后6种棕榈植物的POD酶活性变化总体呈先升高后降低或先升高后降低再升高的趋势(图2)。各种棕榈在温度下降初期,POD酶活性均表现出一定的上升趋势,这能对细胞膜起到较好的保护作用。美丽针葵在-3℃时POD酶活性达到最高,为53.16 U/gFW,是对照温度的2.97倍,随后酶活性呈现出迅速下降趋势,但仍高于对照;-6℃时,银海枣与蒲葵、华盛顿棕的POD酶活性达到最高,分别为63.17 U/gFW、63.14 U/gFW、79.66 U/gFW,分别是对照温度的 2.94 倍、2.13 倍、2.26 倍;当温度降至-9℃时,布迪椰子与加拿利海枣POD酶活性也达到最高,分别为91.50 U/gFW、83.11 U/gFW,分别为对照温度的1.64倍、1.94倍。
2.3 低温胁迫对MDA含量的影响
经低温处理后6种棕榈科植物的MDA含量随温度的下降而呈现出明显的上升趋势,且随着温度的降低其变化幅度增大(图3)。当温度下降至0℃时,各种棕榈的MDA含量略呈上升趋势,可能是因为植物对低温有一定的防御反应,能减缓膜脂过氧化作用,从0~-3℃降温过程中,美丽针葵与银海枣的MDA含量较其他棕榈增长幅度大,细胞膜受到了的伤害较大,到达-3℃时MDA含量也比其他棕榈都高,含量分别为0.35 μmol/g、0.33 μmol/g。当温度降低到-6 ℃时,美丽针葵的 MDA 含量达到了最大值,比对照温度增加了1.05倍。达-9℃时,蒲葵、华盛顿棕与布迪椰子的MDA含量均达到了最大值,其中蒲葵的增长幅度最大,布迪椰子的增长幅度最小。从整体来看,美丽针葵、银海枣的变化幅度相对较大,低温胁迫时积累的MDA较多,细胞膜脂过氧化程度较高,其抗寒性较差;而布迪椰子表现最为稳定,经低温处理后MDA积累得较少,细胞膜脂过氧化程度较低,其抗寒性较强。
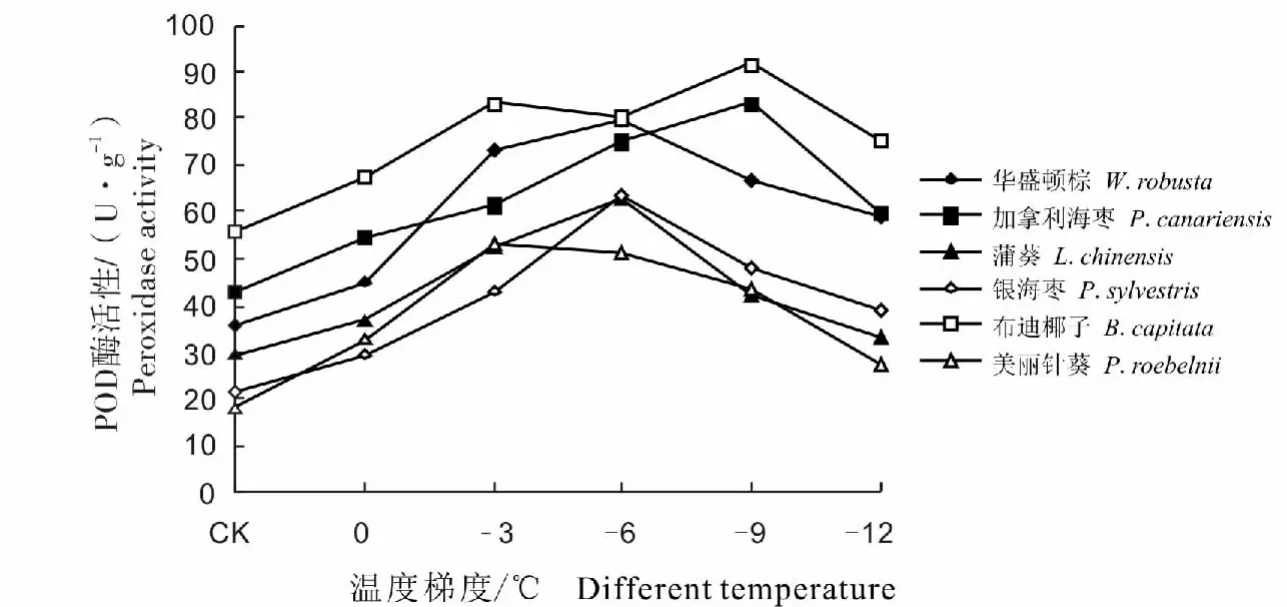
图2 不同低温处理对6种棕榈科植物POD酶活性的影响Fig.2 The effect on POD activities in six palm plants under different low temperature treatment
3 结论与讨论
植物的相对电导率变化是反映植物受逆境胁迫后细胞膜透性变化的重要生理指标,与植物的抗寒性呈负相关[12]。细胞膜透性的损伤是低温伤害的本质之一,直接反映植物膜系统受伤的程度,而相对电导率大小反映了质膜透性的大小[24]。抗寒性较强的植物,膜透性增大程度较小,相对电导率较小;抗寒性较差的植物,膜透性增大程度较大,相对电导率较大。本实验中,随处理温度的降低,6种棕榈科植物的相对电导率逐渐升高,规律较明显,温度越低对细胞膜造成的损伤越大,细胞膜的透性增大,导致细胞内相对电导率也增大。根据电导率变化配合Logistic方程求出植物低温半致死温度,布迪椰子、加拿利海枣、华盛顿棕、银海枣、蒲葵、美丽针葵这6种棕榈科植物的半致死温度分别为:-10.4,-9.5,-8.8,-7.5,-6.2,-2.3 ℃。
POD是植物体内普遍存在活性较高的一种酶,它在保护酶系统中主要是起到酶促降解H2O2的作用,避免细胞膜的过氧化伤害,从而提高植物的抗寒性[25]。本试验中6种棕榈科植物经低温处理后POD酶活性的变化大致相同,且与抗寒性呈正相关关系,在人工低温胁迫下,布迪椰子POD酶活性较高,抗寒性较强;美丽针葵POD酶活性较低,抗寒性较弱。
MDA是膜脂过氧化指标,表示细胞膜脂过氧化程度和植物对逆境条件反应的强弱。植物在低温逆境条件下遭受伤害往往发生膜脂过氧化作用,MDA是膜脂过氧化作用的最终产物,其在组织中含量的多少是膜脂质过氧化作用强弱的一个重要指标,能有效地反映细胞膜系统遭受逆境伤害的程度。MDA含量高说明植物膜脂过氧化程度高,反之,则膜脂过氧化程度低[26]。通过本试验可以看出,低温胁迫下6种棕榈科植物的MDA含量均有不同程度的增加,但增长幅度不同。布迪椰子MDA积累较少,细胞膜脂过氧化程度较低,其抗寒能力较强。美丽针葵、银海枣MDA积累较多,细胞膜脂过氧化程度较高,抗寒能力较差。
综上所述,6种棕榈科植物抗寒性与细胞质膜透性、保护酶活性之间存在较密切的关系,在低温胁迫下6种棕榈科植物呈现出保护性反应,保护酶(POD)活性上升,相对电导率缓慢上升,MDA含量略有升高,之后出现了伤害性反应,POD酶活性下降,相对电导率大幅度上升,MDA含量明显增多。通过本试验得出结论,6种棕榈科植物的抗寒性由强至弱依次为:布迪椰子、加拿利海枣、华盛顿棕、银海枣、蒲葵、美丽针葵。布迪椰子、加拿利海枣抗寒能力强,可以在南昌园林绿地中应用和栽培;银海枣、华盛顿棕、蒲葵抗寒能力也较强,可在南昌地区适量应用,但寒潮来临时应采取防冻措施;美丽针葵抗寒性较差,在南昌园林绿地中应慎用。
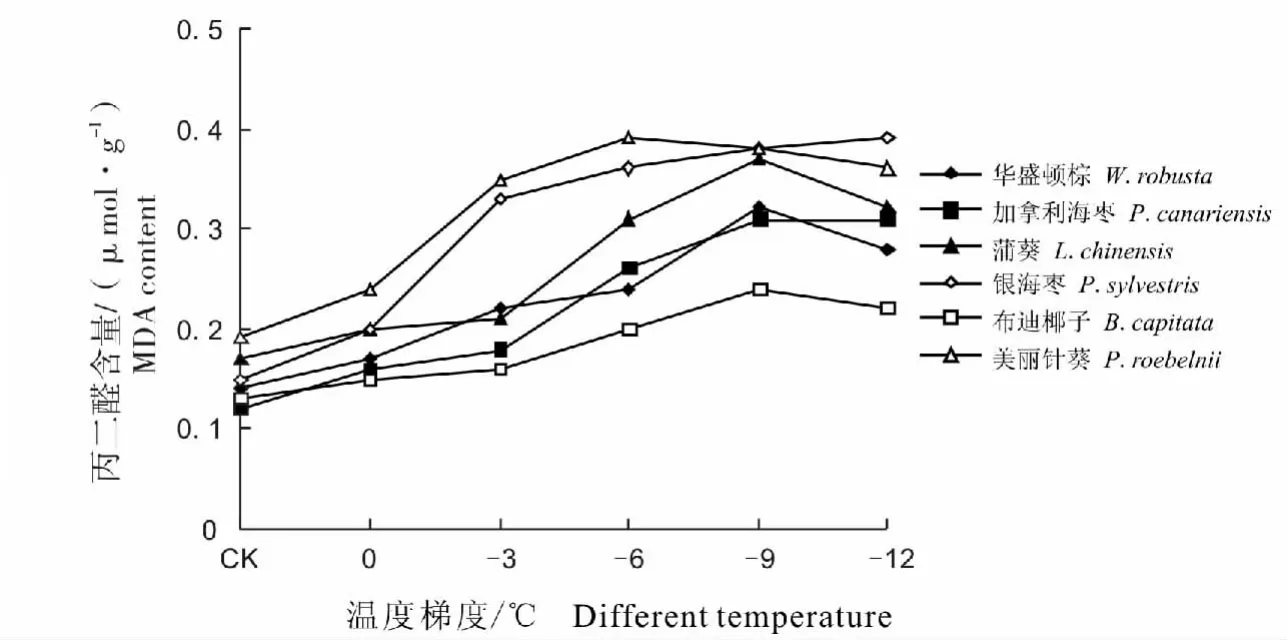
图3 不同低温处理对6种棕榈科植物丙二醛含量的影响Fig.3 The effect on the MDA content in six palm plants under different low temperature treatment
[1]钟如松,何洁英,伍有声,等.引种棕榈图谱[M].合肥:安徽科学技术出版社,2004.
[2]张锡九,代仕高.成都市棕榈科植物引种试验研究[J].四川林业科技,2007,6:14-17.
[3]臧小平,徐雪荣.湛江南亚热带植物园棕榈科观赏植物引种初报[J].亚热带植物科学,2003,32(4):48-51.
[4]何平,陈建雄.攀西地区耐寒棕榈植物的引种研究[J].西昌学院学报:自然科学版,2008(1):16-20.
[5]郑建灿.棕榈科植物在厦门的引种栽培及其应用[J].防护林科技,2003,3:52-53.
[6]张庆费,吴海萍,许东新.上海引种的7种棕榈科植物冻害状况分析[J].浙江林学院学报,2007,24(1):110-114.
[7]袁婷,李阳春,徐维杰.棕榈科植物在南昌园林中应用现状及分析[C].2008年年会暨园艺产业防灾减灾学术研讨会论文集,2009:188-192.
[8]袁婷,徐清峰,徐维杰.南昌市棕榈植物冻害调查与分析[J].现代园艺,2009,3:4-5.
[9]林应枢,黄华庆,徐炜.福州市棕搁科植物寒害调查初报[J].广东园林,1998(1):29-32.
[10]陈振东,林秀香.福建省棕桐科植物冻害调查初报[J].福建热作科技,2001,26(1):33-35.
[11]周昆华.昆明世界园艺博览园棕榈科植物的耐寒性调查[J].热带农业科技,2004,27(1):21-23.
[12]阮志平,廖启炓,丁印龙.4种棕榈科植物在厦门越冬的生理指标比较[J].浙江林学院学报,2007,24(1):115-118.
[13]林秀香,陈振东.我国棕榈科植物的研究进展[J].热带作物学报,2007,3:115-119.
[14]陈星,李俊全,王君玲,等.低温下棕榈某些生理变化及低温锻炼对棕榈耐寒性的影响[J].北京师范大学学报,1999,35(2):257-260.
[15]廖启蚪,丁印龙,杨盛昌,等.低温胁迫下加拿利海枣膜脂过氧化及保护酶活性的变化[J].厦门大学学报,2002,41(5):570—573.
[16]杨盛昌,谢潮添,张平,等.冷锻炼对低温胁迫下夏威夷椰子膜脂过氧化及保护酶活性的影响[J].植物资源与环境学报,2002,11(4):25-28.
[17]杨希,隆学武,曾华浩,等.美丽针葵耐寒性试验[J].福建林业科技,2002,29(2):56-57.
[18]阮志平,向平,李振基.布迪椰子、沼地棕和油棕的耐寒性研究[J].北京林业大学学报,2008(4):77-81.
[19]廖启蚪.低温胁迫下袖珍椰子幼苗的某些生理学变化[J],中国农学通报,2009,25(19):298-302.
[20]朱根海.应用Logistic方程确定植物组织低温半致死温度的研究[J].南京农业大学学报,1986(3):11-16.
[21]邹琦.植物生理学试验指导[M].北京:中国农业出版社,2000.
[22]郝再彬,苍晶,徐仲.植物生理实验[M].哈尔滨:哈尔滨工业大学出版社,2004.
[23]高俊凤.植物生理学实验技术[M].北京:世界图书出版社,2000.
[24]Bluma A,Ebercon A.Cell membrane stability as an measure of drought and heat tolerance in wheat[J].Crop Science,1981,(21):43-47.
[25]孙昌祖,刘家琪.低温胁迫对青杨叶片O2、MDA、膜透性、叶水势及保护酶的影响[J].内蒙古林学院学报,1998,20(3):32-36.
[26]Lirf,Wang L X,Liang Y R.Recent progress in the study of cold resistance of grapes[J].Journal of Inner Mongolia Insititute of Agriculture,1997(6):24-26.
