扁浒苔(Ulva compressa)的分子鉴定及生活史的初步研究
2013-08-14蔡永超马家海高嵩杜晶王丁晶
蔡永超,马家海,高嵩,杜晶,王丁晶
(上海海洋大学 水产与生命学院,上海 201306)
扁浒苔隶属于绿藻门,绿藻纲,石莼目,石莼科,石莼属。藻体亮绿色或暗绿色,管状,常扁压,高10~30 cm,甚至可达40 cm,有许多分枝,基部分枝较密,上部渐疏。分枝简单圆柱状或上部开展,分歧处略缢缩,直上部较粗大,与主枝的形状和大小相似。细胞表面观为角圆的亚方或长方形,直径10~27 cm,排列无序,每个细胞内有蛋白核1个(曾呈奎等,2009)。但是,石莼属种内划分的传统方法是依据形态学特征进行分类,由于石莼属具有种类多、分布广、形态特征复杂,而且许多形态特征随着环境的变化而不稳定的特点,因此,难以准确鉴定石莼属内种类(Malta et al,1999;Blomster et al,1998)。如 U.intestinalis通常没有分枝或者藻体下部稍有分枝,而U.compressa有分枝,但是低盐或盐休克可以诱导 U.intestinalis形成分枝(Blomster et al,1998)。针对这一问题,本文采用了分子鉴定的方法,保证了生活史研究所需藻体种的确定性和可靠性。
在生活史研究方面,Nordby等(1972)认为把U.mutabilis藻体切割成小的片段,可以提高孢子的形成(Nordbyetal,1972);Hiraoka等(1988)通过在孔石莼(U.pertusa)藻体上打小圆孔取样,快速诱导生殖细胞的形成(Hiraokaetal,1988);Dan 等(2002),Lin等(2008),Gao等(2009)都通过打孔取样的方法,快速诱导了浒苔(U.prolifera)的生殖细胞的快速形成(Dan et al,2002;Lin et al,2008;Gao et al,2010)。此种方法大大缩短了生活史研究的周期。
绿潮藻连续爆发6年来,国内在种类鉴定方面,采用分子生物学的方法,已经发现绿潮的组成种类包括浒苔(U.prolifera),扁浒苔(U.compressa),缘管浒苔(U.linza)和曲浒苔(U.flexuosa),其中浒苔(U.prolifera)为绿潮的优势种类,扁浒苔(U.compressa)为绿潮的组成种类之一(应成琦等,2009;陈斌斌等,2012);在生活史方面,对石莼属(Ulva)的研究对象主要是浒苔(U.prolifera)和缘管浒苔(U.linza),如王晓坤等(2007)对浒苔的生活史进行了初步的研究;马家海等(2009)对缘管浒苔的生活史进行了初步的研究;张华伟等(2011)对绿潮漂浮浒苔的繁殖特性进行了研究。但是,国内对扁浒苔(U.compressa)生活史研究的资料尚未见报道。本文通过实验观察,对扁浒苔的生活史进行了详细的报道,旨在完善绿潮藻种类的研究和对绿潮的成因、防治及漂浮的研究提供理论基础。
1 实验材料和方法
1.1 实验材料
实验样品于2011年11月-2012年6月采自江苏及山东沿岸(表1)。
1.2 实验方法
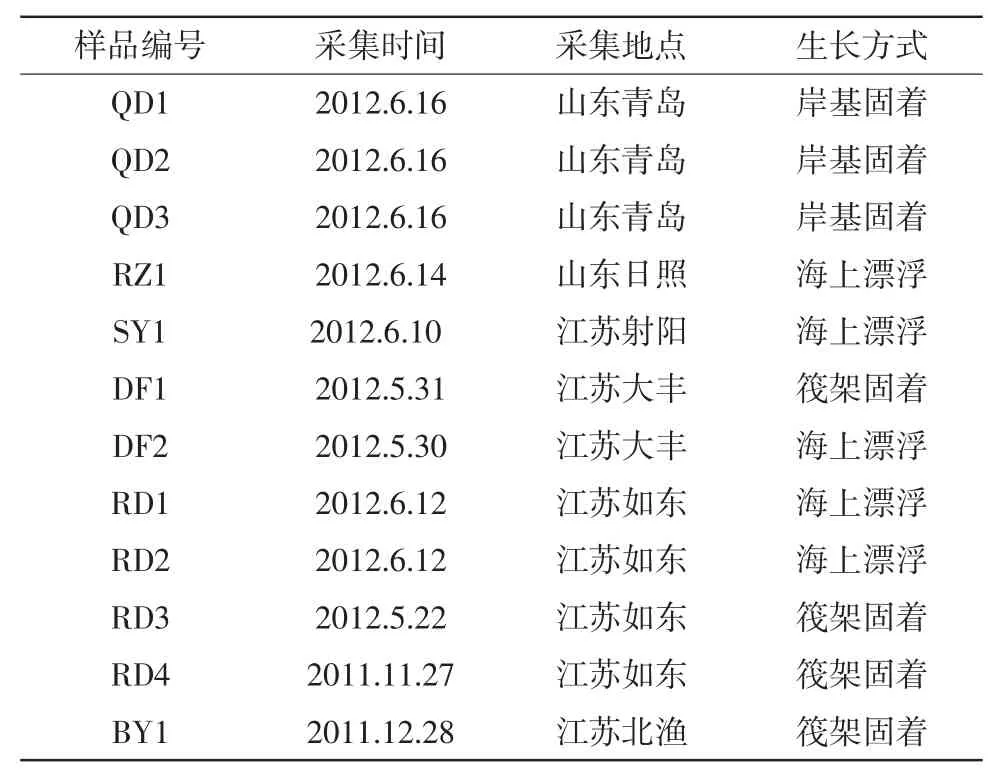
表1 样品信息样品编号采集时间采集地点生长方式
1.2.1 样品的采集与处理
所有野外采集的藻体就地清洗后放在装有冰袋的保温箱中冷藏保存并于24 h内带回实验室,挑选健康的藻体进行充气培养。培养的条件为:温度18℃,海水盐度26.9,光照强度100~160μmol/(m2·s1),光照周期12L∶12D(8∶00-20∶00为光照时间,20∶00-8∶00为黑暗时间),PES营养液浓度为2%。对研究生活史所需要的藻体,参照Hiraoka等(1988)的处理方法并有所改动。
1.2.2 DNA的提取及ITS序列的获取
DNA制备方法参照Doyle等(1990),ITS序列的扩增参考 Hayden等(2003)和 Shimada等(2008)。PCR产物经1%琼脂糖凝胶电泳和溴化乙锭染色后,通过紫外光检测。引物有上海生物工程有限公司合成,扩增产物的纯化及测序由上海迈普生物技术有限公司完成。
1.2.3 ITS序列的分析
采 用 Bioedit(Hall et al, 1999)和 ClustalX(Ver1.83)(Thompson et al,1994)对样品 ITS 序列进行编辑和对齐。同源性通过NCBI进行BLAST检测,获取序列均来自GenBank,系统发育树构建采用邻接法,采用Kimura双参数模型,使用软件为MEGA 4.1,参考Tamura等(2007),自举检验值1 000次检验节点置信度。
1.2.4 培养镜检
用1 000 uL移液器的枪头在靠近扁浒苔藻体顶端的边缘进行打孔取样,取下的小圆片藻体放置在培养皿中(高为18 mm,直径为90 mm),加含有PES营养液的灭菌海水50 ml,培养的条件为:温度18℃,海水盐度26.9,光照强度100~160 μmol/(m2·s1),光照周期12L∶12D(8∶00-20∶00为光照时间,20∶00-8∶00 为黑暗时间),每天8∶00到13∶00,用Olympus显微镜对其进行显微观察及拍照。
1.2.5 配子和孢子的趋光性
把成熟的生殖的藻体放在100 mL的小烧杯中,用手电筒将光线从锥形瓶的一侧射入,镜检聚集地方的生殖细胞以测试其趋光性。
2 实验结果
2.1 ITS 系统发育分析
以ITS序列构建的N-J系统发育树见图1。12个样品一共被分到了3个进化枝,分别是LPP(U.linza-procera-prolifera)、U.compressa、U.flexuosa.其中LPP类群占有优势,包含7个样品,U.compressa类群包含4个样品,U.flexuosa类群包含1个样品。

图1 基于ITS序列所构建的N-J系统发育树
2.2 扁浒苔(U.compressa)的生活史
2.2.1 孢子囊和配子囊的成熟过程
经过3 d到4 d的培养,样品的细胞形态发生了巨大的变化。小圆片藻体的正常细胞(图版Ⅰ-1),经过2 d左右,色素体分解为粒状,并分布在整个细胞中(图版Ⅰ-2),4 d左右,色素体融合,并集中在一个细胞的一边,空泡形成(图版Ⅰ-3),5 d左右,配子囊(图版Ⅰ-4)和孢子囊(图版Ⅰ-6)形成。
2.2.2 生殖细胞的数量及趋光性
实验结果显示,扁浒苔的每个配子囊里含有16个配子(图版Ⅰ-4-5),每个孢子囊里含有8个孢子(图版Ⅰ-6-7)。将装有一定量生殖细胞的锥形瓶置于电筒下,一段时间后烧杯内大量生殖细胞在锥形瓶接近光源的一侧聚集,形成一层绿色的膜(图版Ⅰ-8),镜检可见大量的配子;在远离光源的区域发现少量的合子。由此可见,扁浒苔的配子具有正趋光性,而合子具负趋光性。随着培养时间的延长,慢慢形成簇生小苗(图版Ⅰ-9)。
2.2.3 单性生殖、无性生殖及有性生殖的过程
通过实验观察,扁浒苔的生殖方式包含以下几种繁殖方式。单性生殖:配子体藻体放散的两鞭毛配子(图版Ⅱ-1)运动一段时间后,雌雄配子不经过结合,直接附着后发育。放散的两鞭毛配子可以直接在基质上附着发育(图版Ⅱ-2),也可以附着在原藻体上发育(图版Ⅱ-3-4)。此外,有的配子囊未完全放散配子,配子囊里含有的一个(图版Ⅱ-5)或多个(图版Ⅱ-6-7)配子直接在藻体上发育。无性生殖:孢子体藻体放散的四鞭毛游孢子(图版Ⅱ-8),游孢子运动一段时间找到合适的基质后附着发育(图版Ⅱ-9)。孢子囊同样存在放散不完全,孢子于囊内直接生长发育的现象(图版Ⅱ-10)。有性生殖:配子体藻体放散两鞭毛的配子,雌雄配子在结合的过程中,雌雄配子相互旋转,色素体向尾部移动,从头部开始融合,然后雌性配子相互靠近,再从侧面开始融合,最后完成彼此的融合,形成合子,合子慢慢变圆并附着发育(图Ⅱ-11-16)。在雌雄配子融合的全过程,雌雄配子始终保持着同步的相互旋转和移动。此外,实验还发现,有性生殖过程中,雌雄配子的结合需要一定的外部环境和时间,所以这种生殖方式较为少见。游动孢子和配子在进行附着的过程中均具有聚集性(图版Ⅱ-17),但两鞭毛的配子比四鞭毛的孢子运动的更快,更易聚集附着发育,所以单性生殖又占有主导的地位。

图版Ⅱ 扁浒苔的生活史
3 讨论
在我国,按照传统的形态学方法对石莼属进行分类,已报道的石莼属中的浒苔有7种,包括浒苔(U.prolifera)、缘管浒苔(U.linza)、扁浒苔(U.compressa)、曲浒苔(U.flexuosa)、肠浒苔(U.intestinlis)、管浒苔(U.tubulosa)、条浒苔(U.clathrata),广泛分布于东北亚沿海的近岸(Tseng et al,1983;杨君 等,2000;曾呈奎 等,1962;曾呈奎等,2009)。但是,本研究通过分子生物学的方法把所选的12个藻类样品划分为3个类群,即U.linza-procera-prolifera(LPP)复合体类群、U.compressa类群以及U.flexuosal类群。到目前为止,采用分子生物学方法只发现其中4种,为浒苔(U.prolifera),扁浒苔(U.compressa),缘管浒苔(U.linza)和曲浒苔(U.flexuosa)(应成琦等,2009;陈斌斌等,2012),其余3种到底存不存在,以及是否是由于传统形态学分类方法造成的还有待于进一步的研究。因此,在今后的石莼属藻类的种类鉴定中,考虑以现代的分子生物学方法为主,以形态学观察为辅为参考,使得分子生物学方法和传统形态学方法相辅相成,以便对绿藻种类的鉴定更为可靠。
扁浒苔是绿潮藻的组成种类,由于它的表面积一般较大,可以为绿潮藻的优势种浒苔及其他种类提供配子或者孢子的附着基质,有利于这些藻体的快速繁殖。同时,海洋生物的摄食,人类的活动,海浪潮汐等自然作用,都会导致扁浒苔破裂形成小片段藻体,这些产生的小片段的藻体可快速诱导生殖细胞的产生,这就大大的增加了包括扁浒苔在内的绿潮藻的繁殖速度,短时间内形成大量的绿潮(Nordby et al,1972;Hiraoka et al,1998;Dan et al,2002;Lin et al,2008)。
在U.compressa的生活史中,无性生殖和单性生殖占有优势。因为不论是在无性生殖过程中的孢子还是单性生殖过程中的配子,一经附着即可生长,无须经过有性生殖雌雄配子的接合过程,这就缩短了繁殖周期,为藻体在短时间内的快速生长提供了条件。此外配子囊和孢子囊放散配子和孢子的数量也是惊人的,在海况条件适宜的情况下,使U.compressa等漂浮绿藻大量繁殖,从而可能成为“绿潮”的成因之一。
本文在国内首次报道了作为黄海海域漂浮绿藻种类之一的U.compressa的生活史,掌握其生活史将有助于了解、分析黄海海域漂浮绿藻大量迅速增殖的成因,同时为“绿潮”的预测、预报与防治提供了理论依据。
Blomster J,Maggs C A,Stanhope M J,1998.Molecular and morphological analysis of Enteromorpha intestinalis and E.compressa(Chlorophyta)in the British Isles.Journal of Physiology,34(2):319-340.
Dan A,Hiraoka M,Ohno M,et al,2002.Observations on the effect of salinity and photon fluence rate on the induction of sporulation and rhizoid formation in the green alga Enteromorpha prolifera(Müller)J.Agardh(Chlorophyta,Ulvales).Fisheries Science,68(6):1182-1188.
DoyleJ J,DoyleJ L,1990.Isolation of plant DNA from fresh tissue.Focus,12(12):13-15.
Gao S,Chen X Y,Yi Q,et al,2010.A strategy for the proliferation of Ulva prolifera,main causative species of green tides,with formation of sporangia by fragmentation.PLoS ONE,5(1):e8571.
HallT A,1999.BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT.Nucleic Acids Symposium Series,41:95-98.
HaydenS H,Blomster J,Maggs A C,et al,2003.Linnaeus was right all along:Ulva and Enteromorpha are not distinct genera.European Journal of Phycology,38(3):277-294.
Hiraoka M,Enomoto S,1998.The induction of reproductive cell formation of Ulv apertusa Kjellman(Ulvales,Ulvophyceae).Phycological Research,46:199-203.
Lin A P,Shen S D,Wang J W,et al,2008.Reproduction diversity of Enteromorpha prolifera.Journal of Integrative Plant Biology,50(5):622-629
Malta E J,Draisma S G A,Kamermans P,1999.Free-floating Ulva in the southwest Netherlands:species or morphotypes?A morphological,molecular and ecological comparison.European Journal of Phycology,34(5):443-454.
Nordby Ø,Hoxmark R C,1972.Changes in cellular parameters during synchronous meiosis in Ulva mutabilis Føyn.ExptI Cell Res.75:321-328.
Shimada S,Yokoyama N,Arai S,et al,2008.Phylogeography of the genus Ulva(Ulvophyceae,Chlorophyta),with special reference to the Japanese freshwater and brackish taxa.Journal of Applied Phycology,20(5):979-989.
Tamura K,Dudley J,Nei M,et al,2007.MEGA4:Molecular Evolutionary Genetics Analysis(MEGA)software version 4.0.Molecular Biology and Evolution,24(8):1596-1599.
Thompson J D,Higgins D G,Gibson T J,1994.CLUSTAL W:improving the sensitivity of progressive multiple sequence alignment through sequence weighting,position-specific gap penalties and weight matrix choice.Nucleic Acids Research,22(22):4673-4680.
Tseng C K,1983.Common Seaweeds of China.Beijing:Science Press.
陈斌斌,马家海,张天夫,等,2012.中国沿海石莼属绿藻的ITS和5SrRNA序列分析.海洋渔业,34(3):249-255.
马家海,嵇嘉民,徐韧,等,2009.长石莼(缘管浒苔)生活史的初步研究.水产学报,33(1):45-52.
王晓坤,马家海,叶道才,等,2007.浒苔(Enteromorpha prolifera)生活史的初步研究.海洋通报,26(5):112-116.
杨君,安利佳,王茜,等,2000.石莼属(Ulva)与浒苔属(Enteromorpha)绿藻的RAPD分析.海洋与湖沼,31(4):408-413.
应成琦,张婷,李信书,等,2009.我国近海浒苔漂浮种类ITS与18SrDNA序列相似性分析.水产学报,33(2):215-219.
曾呈奎,夏邦美,周显铜,等,2009.中国黄渤海海藻.北京:科学出版社,389-390.
曾呈奎,张德瑞,张峻甫,等,1962.中国经济海藻志.北京:科学出版社,42-49.
张华伟,马家海,胡翔,等,2011.绿潮漂浮浒苔繁殖特性的研究.上海海洋大学学报,20(4):600-606.
